Introduction
Neuron restrictive silencer factor (NRSF), also called the repressor element 1 (RE-1) silencing transcription factor (REST), is a major transcriptional repressor for neuron-specific genes in non-neuronal cells and neuronal progenitor cells [1-
3]. NRSF/REST mediates transcription repression through the association of its amino terminal repression domain with the mSin3/histone deacethylase 1/2 (HDAC 1/2) complex [4-6].
and through the association of its carboxy terminal repression domain with the CoREST/HDAC complex [7,8]. NRSF/
REST bind to a 21 bp consensus sequence of neural restrictive silencer element (NRSE), also known as the RE-1, in the regulatory regions of many neuron-specific genes: ion channels, neurotranssmitter receptor, neurotrophic receptor, neuronal cell adhesion molecules, neuronal growth factors, neuronal cytoskeleton, and others [9,10].
The SLIT and NTRK-like family is a family of structurally related transmembrane proteins with an extracellular leucine-
Regulation of SLITRK1 gene by neuron restrictive silencer factor in NMB cells
Myung Mi Kim 1,† , Jin-Soo Kim 1,† , Sung-Min Moon 1 , Mi Suk Choi 1 , Bo Ram Park 1 , Dong-Seol Lee 2 , Shin-Yeob Mo 2 , Seon-Ho Cho 2 , Chun Sung Kim 1, *
1
Department of Oral Biochemistry,
2Oral Biology Research Institute, School of Dentistry, Chosun University, Gwangju, Korea
ABSTRACT
Purpose: The Slitrk family are neuronal transmembrane proteins that participate in the regulation of neurite outgrowth and are selectively expressed in the brains of adult mice. Neuron restrictive silencer factor (NRSF), also called RE1-silencing transcription factor, was originally discovered as a transcriptional repressor in non-neuronal cells. However, in this study, we show that the neuron restrictive silencer element (NRSE) of SLITRK1 functions as a critical regulator to repress SLITRK1 gene expression in NMB cells.
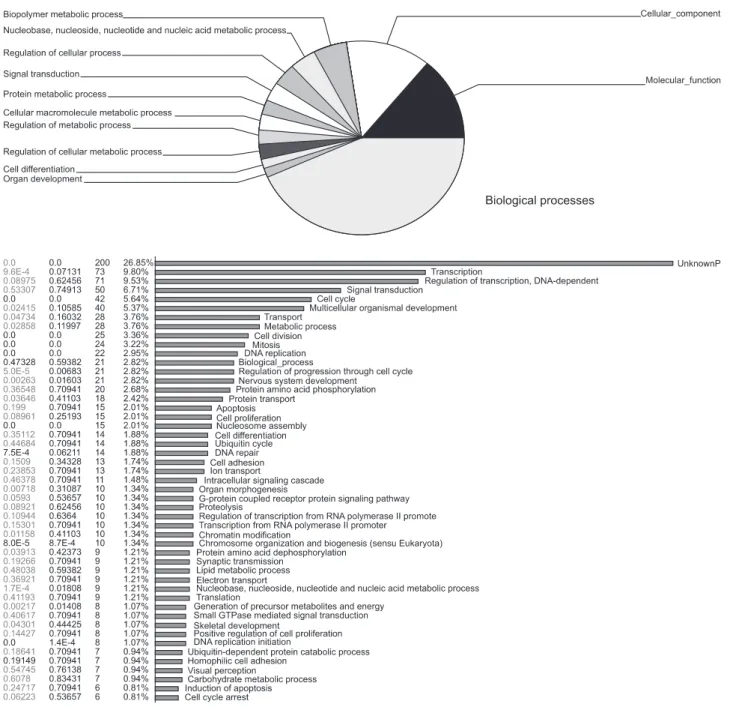
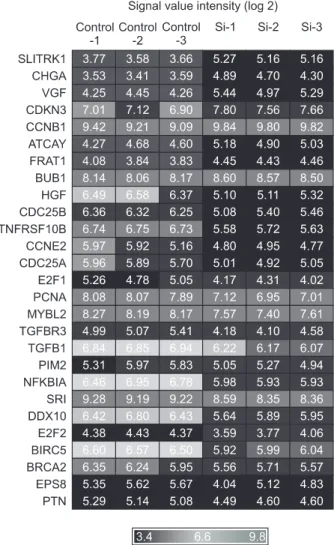
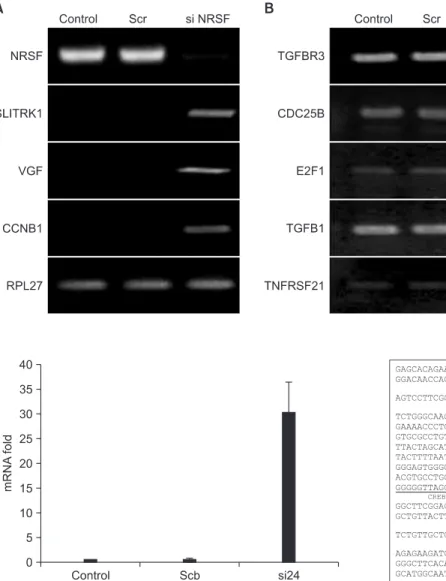
Materials and Methods: To screen NRSF/NRSE-regulated specific genes, we performed microarray analysis of NRSF siRNA- transfected NMB cells. We selected eight genes and confirmed their expression by RT-PCR analysis.
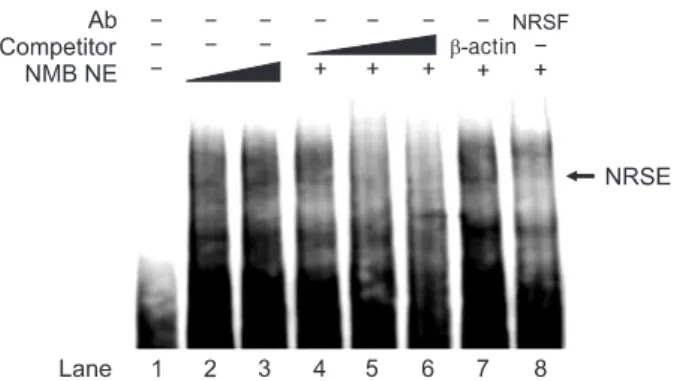
Results: Expression levels of SLITRK1, VGF, and CCNB1 were up-regulated in NRSF siRNA-transfected NMB cells compared with control NMB cells, whereas TGFBR3, CDC25B, E2F1, TGFB1, and TNFRSF21 were slightly down-regulated or showed no change. Especially, we identified three putative NRSF-binding sequences in the SLITRK1 gene, and its transcription was significantly up-regulated in NRSF siRNA-transfected NMB cells as measured by real-time PCR. Using co-transfection studies, we determined that the second NRSE of the SLITRK1 gene was functional in NMB cells. In addition, NRSF bound to the second NRSE of the SLITRK1 gene in a sequence-specific manner as confirmed by supershift assay. Our data suggest that NRSF interacts with and represses expression of the SLITRK1 gene.
Conclusion: NRSF plays an important role in regulating the growth and development of neuronal genes via SLITRK1 gene transcription.
Key Words: RE1-silencing transcription factor, Neuron restrictive silencer element, Transcriptional repressor, SLITRK1
Received Jun 19, 2013; Revised version received Jul 31, 2013 Accepted Jul 31, 2013
Corresponding author: Chun Sung Kim
Department of Oral Biochemistry, School of Dentistry, Chosun University, 309 Pilmun-daero, Dong-gu, Gwangju 501-759, Korea Tel: 82-62-230-7088, Fax: 82-62-224-3706
E-mail: [email protected]
†