Vol. 8, No. 4, December, 2001
류마티스 활막세포에서 비스테로이드성 항염제에 의한 Vascular Endothelial Gro wth Factor 생성의 억제
가톨릭대학교 강남성모병원 류마티스내과, 가톨릭대학교 의과학연구원* ,
김용주・박종서・최진정・민소연*・조미라*・김완욱・박성환・조철수・김호연
─A b s t r a c t─
Inhibition of Vascular Endothelial Growth Factor Expression in Cultured Rheumatoid Synoviocytes by Nonsteroidal Anti-inflammatory Drugs
Yong-Ju Kim, M.D., Jong-Seo Park, M.D., Jin-Jung Choi, M.D., Mi-La Cho, M.D.*, So-Youn Min, M.D.*, Wan-Uk Kim, M.D., Sung-Hwan Park, M.D.,
Chul-Soo Cho, M.D., Ho-Youn Kim, M.D.
Division of Rheumatology, Kangnam St. Mary’s hospital,
The Catholic University of Korea, Catholic Research Institutes of Medical Sciences*,
O b j e c t i v e : Vascular endothelial growth factor (VEGF), a potent angiogenic, permeability-enhancing cytokine plays an important role in chronic inflammato- ry process of rheumatoid arthritis (RA). Nonsteroidal anti-inflammatory drugs (NSAIDs) are the most widely used drugs for the treatment of RA. However, the effect of NSAIDs on angiogenesis in rheumatoid synovium is unclear. In this study, we investigated the effects of NSAIDs such as indomethacin (IDC) on TGF-β-induced VEGF production in rheumatoid synoviocytes.
M e t h o d s : Fibroblast-like synoviocytes (FLS) from RA were stimulated with T G F -β(10 ng/ml) for 24hr in the presence of the various concentrations of IDC.
The levels of VEGF were measured in culture supernatant by ELISA. In addition, COX-2 and VEGF mRNA expression of cultured FLS were evaluated by RT-PCR.
<접수일 : 2001년 9월 27일, 심사통과일 : 2001년 10월 8일>
※통신저자 : 김 호 연
서울특별시 서초구 반포동 505번지 가톨릭대학교 내과학교실 류마티스내과
Tel : 02) 590-2702, Fax : 02) 537-4673, E-mail : [email protected]
*본 논문은 한국과학재단(2000-2-21200-001-3) 연구비 지원으로 이루어짐.
서 론
류마티스 관절염은 혈관형성 촉진인자와 억제인자 사이의 전체적 균형에 의해 좌우되는 신생혈관형성 의존성 질환으로1 , 2 ), 조직학적으로 증식된 활막세포, 염증세포, 혈관으로 구성된 활막의 종양성 증식을 특징으로 한다3 ). 섬유혈관육아조직의 과성장 및 활 막내 단핵세포 이주를 위해서는 풍부한 혈관이 요구 되므로, 신생혈관형성은 류마티스 활막염의 진행 및 영속성에 중요한 역할을 한다.
신생혈관형성은 이미 존재하는 모세혈관에서 새로 운 혈관이 성장하고 증식하는 것으로, 정상적인 생 리적 성장, 창상치유, 신생물 형성, 당뇨병성 망막병 증, 염증성 장애 등과 같이 다양한 생리적, 병적인 과정에 필수적 역할을 한다4 ). 정상 관절연골은 혈관 이 없고 신생혈관형성에 대해 저항성이 있지만, 류 마티스 관절염에서는 이러한 장애물이 파괴되어 신 생혈관이 관절연골내로 성장하게 된다. 정상 및 염 증성 활막에서 신생혈관형성에 중요한 혈관형성 촉 진인자로는 acidic fibroblast growth factor (FGF), basic FGF, platelet derived growth factor (PDGF), transforming growth factor- β( T G F -β), tumor necrosis factor-α( T N F -α) , vascular endothelial growth factor (VEGF), angiogenin 등이 알려져 있다1 , 3 , 4 ).
V E G F는 류마티스 관절염 환자의 활성화된 대식 세포와 활막섬유아세포에서 분비되는 이가(二價)성 의 당단백질로써, 주로 혈관내피세포에 작용하여 혈
관내피세포의 증식, 혈관형성, 혈관 투과성을 증가 시킨다5 ). VEGF는 골관절염 또는 다른 형태의 관절 염보다 류마티스 관절염 환자의 활액에서 의미있게 증가되어 있으며5 , 6 ), VEGF 분비를 증가시키는 인자 로서 저산소증, 프로스타글란딘 E2( P r o s t a g l a n d i n E2, PGE2), IL-1β, IL-6, TNF-α, CD40 ligand 등7 )이 알려져 있다.
싸이클로옥시겐효소(cyclooxygenase, COX)는 세포막에서 유리된 아라키돈산(arachidonic acid)을 프로스타글란딘으로 전환시키는 효소로 두 종류가 있 다. COX-1은 여러 종류의 세포에서 내재적으로 (constitutively) 발현되나 C O X - 2는 다양한 염증유 발물질, 성장인자 등에 의해서 유도된다. 최근 보고 에 의하면8 - 1 0 ), 다양한 염증조직, 종양, 류마티스 관 절염의 활막조직에 고농도로 존재하는 C O X - 2와 프 로스타글란딘은 류마티스 관절염 및 종양 진행과 연 관되어 있음이 제시되고 있다. 다양한 비스테로이드 성 항염제(nonsteroidal anti-inflammatory drugs, NSAIDs)가 신생혈관형성 억제를 통해 종양 세포의 증식억제 및 세포사멸( a p o p t o s i s )에 관여한 다는 보고는 많으나1 1 - 1 5 )
, 류마티스 관절염 환자의 배 양된 활막세포에 대한 신생혈관형성 억제효과에 대해 서는 아직 보고 된 바가 없다. 본 연구에서는 류마티 스 관절염의 치료에 사용되는 비스테로이드성 항염제 가 류마티스 활막세포에서 VEGF 생성과 관련된 신 생혈관형성에 미치는 영향을 알아보고자 하였다.
R e s u l t s : VEGF production from FLS was significantly increased in the presence of TGF-β. IDC exerted a dose-dependent inhibitory effect on the production of VEGF induced by TGF-β. RT-PCR analysis showed that IDC also inhibited TGF-β- i n d u c e d COX-2 and VEGF mRNA expression in cultured FLS by a dose-dependent manner.
C o n c l u s i o n : Our results demonstrate that NSAIDs inhibit VEGF production and the expression of its mRNA and COX-2 mRNA in synovial cells of RA patients. These findings suggest that NSAIDs may suppress progression and per- petuation of rheumatoid synovitis by anti-angiogenic activity.
Key Words: Vascular endothelial growth factor, Nonsteroidal anti-inflammatory drugs, Angiogenesis, Rheumatoid arthritis.
대상 및 방법
1. 재료 1) 시약
재 조 합 T N F -α와 I L - 1β는 E n d o g e n Inc.(Woburn, MA)으로부터 구입하였으며, 재조 합 T G F -β및 I F N -γ는 R & D Systems (Min- neapolis, MN)로부터 구입하였다. Indomethacin ( I D C )는 Sigma Bio Sciences (St. Louis, MO) 에서 구입하였다.
2. 방법
1) 활막세포 분리 및 배양
활막조직은 관절 전치환수술을 시행한 류마티스 관절염 환자에서 얻었다. 얻어진 조직을 2 ~ 3 m m 크기로 잘게 부순 후 0.01% collagenase (Sigma Bio Sciences)로 2시간 동안 처리한 후 3 7℃ 항온 수조에서 흔들어 준 후, 분리된 세포를 1 0분 동안 원심분리( 1 2 0 0 r p m )하여 상층액은 제거하고 남아 있는 p e l l e t을 serum-free DMEM (Life Tech- nologies, Inc., Geithersburg, MD)으로 2회 씻 어주었다. 잘 현탁된 세포를 취해 총 세포수를 계산 한 후 p e l l e t을 1 ~ 2×1 06c e l l / m l가 되도록 1 0 % DMEM[10% FBS(Life Technologies), 2mM glutamine, penicillin(100U/ml), strepto- m y c i n ( 1 0 0㎍/㎖)가 포함된 D M E M ]에서 세포를 재현탁한 후 2 5 c m2 플라스크에 분주하였다. 이후 3 7℃, 5% CO2에서 배양하였고, 배양액은 매 3일마 다 교환하였으며 플라스크 바닥의 9 0 ~ 9 5 %가 활막 세포로 충만 되었을 경우 계대 배양하였다.
2) 각종 싸이토카인과 i n d o m e t h a c i n의 VEGF 생성에 대한 영향
활막세포(fibroblast-like synoviocytes, FLS) 는 4~8 계대 배양된 세포를 사용하였다. FLS는 1 mL DMEM/5% FBS에 5×1 04세포가 포함되도록 하여 각 w e l l에 분주한 후 3 7℃, 5% CO2 조건의 배양기에서 2 4시간 동안 배양하였다. 그후 배지를 insulin-transferrin-selenium A (ITSA; Sigma Bio Sciences)가 포함된 serum-free DMEM으로 교환한 후 4 8시간 배양하였다. 다시 신선한 배양액
으로 교환 후, 배지에 각각 T G F -β ( 1 0 n g / m l ) , T N F -α(1ng/ml), IL-1β(10ng/ml), IFN-γ ( 1 0 0 0 U / m l )를 첨가한 후 2 4시간 배양하였다. 또한 각 w e l l에 다양한 농도의 I D C ( 1 ~ 1 0 0μM )을 처리한 후 6시간째에 T G F -β로 자극하였다. 배양 상층액의 V E G F는 sandwich ELISA법으로 측정하였다.
3) ELISA법에 의한 배양 상층액의 VEGF 측정 9 6개의 well microtiter plate에 0 . 4μl / m l의 g o a t anti-human VEGF165(R&D systems, Inc.)를 5 0 μl를 붙인 후 50 mM sodium carbonate (pH 9.6) 를 완충액으로 사용하였다. 4℃에서 밤새 배양한 후 실온에서 1% BSA/PBS로 2시간 동안 차단시켰다.
인형재조합 V E G F1 6 5(R&D systems, Inc.) 혹은 검 체를 각각의 w e l l에 첨가 후 실온에서 2시간 동안 반 응시켰다. Plate는 100 ng/㎖ biotinylated goat anti-human VEGF1 6 5 (R&D systems, Inc.)를 처 리한 후 실온에서 2시간 동안 반응시킨 후, 1:2000으 로 희석된 p e r o x i d a s e가 표지된 extravidin (Sigma Bio Sciences)을 처리한 후 실온에서 2시간 동안 반응 시켰다. 발색반응은 기질용액으로 T M B / H2O2를 이용 하여 유도하였고, 30분 후에 1M phosphoric acid를 이용하여 반응을 중지시켰다. 흡광도는 파장이 450nm 일 때 자동화된 마이크로평판 판독기( V m a x , Molecular Devices, Palo Alto, CA)를 이용하여 측 정하였다. 각 단계사이의 세척은 0.1% Tween 20을 포함하는 P B S를 사용하였다. 10에서 2000pg/ml 사 이의 표준측정(calibration standard)은 배양배지에 서 인형재조합 V E G F1 6 5농도의 로그값에 대한 흡광도 를 이용한 표준곡선을 그려 교정하였다.
4) RNA 분리
활막세포( 5×1 04)가 포함된 세포 배양액 1 m l를 1 0 0 c m2d i s h에 부유한 후 다양한 농도의 I D C ( 1 ~ 1 0 0μ M )을 넣고 2 4시간 3 7℃, 5% CO2조건의 배양기에서 배양하였다. 이렇게 처리한 활막세포로부터 R N A z o l B isolation kit (TEL-TEST, Inc., Friendswood, T X )를 이용하여 총 R N A를 추출하였다.
5) cDNA 제조
추출한 총 R N A를 주형으로 역전사 효소에 의하 여 c D N A를 만들었으며 이를 위해 B o e h r i n g e r
Manheim (Manheim, Germany)사의 cDNA 합 성 키트를 사용하였다. Diethylpyrocarbonate (DEPC) 증류수에 용해되어 있는 총 R N A에서 5
㎍를 취하여 총 용적이 1 3μl가 되게 증류수를 추가한 후 2μl의 random hexamer (Boehringer Man- h e i m )를 가하고 7 0℃의 수조에서 5분간 방치한 후 얼음에 담가 급냉시켰다. 5μl의 first strand cDNA 합성 반응 완충액(Life Technologies, Inc.; 250 mM Tris-HCl, 15 mM MgCl2, 375 mM KCl, pH 8.5), 2.5μl의 0.1M DTT (Life Technolo- gies, Inc.), 1.25μl의 10 mM dNTP 혼합물 (Life Technologies, Inc.), 1.25μl의 RNase 억제 제(Takara Shuzo Co., Ltd., Shiga, Japan)를 포함하는 혼합액 1 0μl에 1μl의 역전사 효소( L i f e Technologies, Inc., 200U/μl )를 섞은 뒤 G e n e Amp PCR system 2400 (Roche Molecular Sys- tem, Inc., Branchburg, NJ)내에서 4 2℃에서 2 시간 동안 반응시켜 c D N A를 얻었다.
6) Polymerase-Chain Reaction (PCR)
본 실험에 사용한 COX-2, VEGF, 그리고 g l y c- eraldehyde-3-phosphate dehydrogenase ( G A P D H )의 시발체( p r i m e r )는 모두 G e n o t e c Co(Seoul, Korea)에서 구입하였고 염기서열은 표- 1과 같다. COX-2, VEGF, GAPDH의 PCR 산물 은 각각 539 bp, 350 bp, 450 bp로써 P C R이 끝 난 후 전기영동상 분절의 크기를 확인하였다. PCR 과정 중 각각 서로 다른 양의 역전사된 c D N A와 PCR cycle수를 변경하면서 P C R을 시행하여 대수 적으로 증가하는 c D N A양과 PCR cycle 수를 결정 하였다. PCR을 위한 반응 화합물은 총 2 5μl가 되도 록 하였고 용액의 조성은 2 . 5μl의 10X 반응 완충액 (Takara Shuzo Co., Ltd.; 100 mM Tris-HCl, pH 8.3, 15mM MgCl2, 500mM KCl), 2μl의 세 가지 종류의 시발체, 0.5 unit Taq DNA poly- merase (Takara Shuzo Co., Ltd.), 4μl의 2.5mM dNTP (Takara Shuzo Co., Ltd.)를 함 유한 용액내에 2μl의 역전사된 c D N A를 첨가한 후 시행하였다. 증폭을 위해 Gene Amp PCR sys- tem 2400을 사용하였다. 변성을 위하여 9 4℃에서 5분간, annealing을 위하여 C O X - 2에서는 5 5℃에 서 4 5초, VEGF에서는 6 2℃에서 1분, GAPDH에
서는 5 5℃에서 1분, extension을 위해서는 7 2℃에 서 3 0초간의 과정을 C O X - 2에서는 2 3회, 72℃에서 1분간의 과정을 V E G F에서는 2 7회, 72℃에서 4 5초 간의 과정을 G A P D H에서는 3 5회 반복하였다. 음성 대조군으로 역전사 효소 대신 증류수를 사용하여 시 행한 PCR 반응에서 GAPDH PCR 산물이 관찰되 지 않도록 하여 PCR 오염이 없도록 확인하였다.
7) PCR 산물의 정량
PCR 산물 2 4μl에 6X loading buffer 5μl와 혼 합하고 1.5% agarose gel (Sigma Bio Sciences) 에서 100 volt로 전기영동을 시행하였다. Size m a r k e r로는 100 bp DNA ladder (Life Tech- nologies, Inc.)를 사용하였다. 전기영동한 겔은 imaging plate (Fuji Photofilm, Japan)에 노출 시켰으며 BAS 판독기(Fuji Film Co, Japan)를 이용하여 영상을 얻어낸 후 TINA program를 사용 하여 COX-2, VEGF, GAPDH mRNA의 농도계 측적( d e n s i t o m e t r i c )분석을 시행하였다. COX-2, VEGF mRNA의 발현은 GAPDH mRNA의 결과 로 나누어 분석 비교하였다.
3. 통계분석
결과는 산술평균±표준편차로 표시하였고 통계처 리는 W i n d o w s용 SPSS 8.0 프로그램( S P S S , Chicago, IL)을 사용하여 분석하였다. VEGF E L I S A에서 얻어진 결과는 비 모수적 검정( M a n n - Whitney test)을 통해 p<0 . 0 5일 때 통계적으로 유의하다고 분석하였다.
결 과
1. 활막세포의 V E G F생성에 대한 각종 싸이토카인의 영향
여러 가지 염증유발 싸인토카인에 의한 활막세포 의 V E G F생성을 보기 위해, FLS는 각종 싸이토카 인( T G F -β, IL-1β, TNF-α, IFN-γ)으로 자극한 후 2 4시간 배양하였다. 활막세포 단독 배양으로 생성된 V E G F양에 대한 각종 싸이토카인 자극으로 생성된 V E G F양은 T G F -β의 경우 5배, IL-1β는 2 . 6 4배, T N F -α는 1 . 5 1배, IFN-γ는 1 . 6 9배로 증가하여, T G F -β(10 ng/㎖)가 V E G F생성을 위한 가장 강력
한 자극제임을 알 수 있었다(그림 1 ) .
2. 배양된 활막세포에서 i n d e m e t h a c i n의 V E G F생 성에 대한 영향
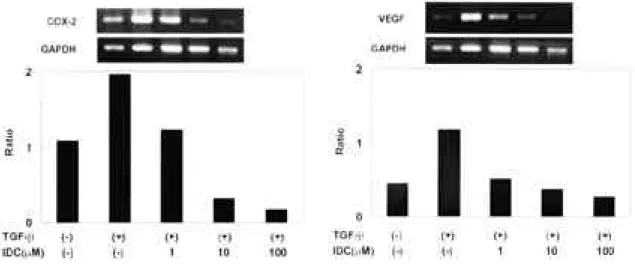
T G F -β(10 ng/㎖) 자극이 없었던 경우, 활막세 포에서의 V E G F생성은 I D C의 1 μM 농도에서는 통 계적으로 유의한 감소가 없었지만(p=0.054), 점차 농도를 증가시킴에 따라(10, 100 μM) 활막세포에서 의 V E G F생 성 은 의 미 있 게 감 소 하 였 다 ( p
<0 . 0 0 5 ) (그림. 2). 또한 활막세포를 I D C ( 1 ~ 1 0 0μ M )로 처리한 후 6시간 뒤에 T G F -β( 1 0 n g /㎖)로 자극한 경우, IDC농도가 증가함에 따라 V E G F생성 은 통계적으로 유의하게 감소하였고( p<0 . 0 0 5 ) , I D C를 100 μM 처리한 경우 처리하지 않은 경우보 다 1 . 8배 감소하였다(그림 2). 선택적 COX-2 억제 제인 NS398 역시 I D C와 유사한 결과를 보였다(결 과 생략). IDC는 사용된 농도에서 비특이적 독성에 의한 세포사가 관찰되지 않았다(결과 생략) .
3. COX-2 mRNA발현에 대한 i n d o m e t h a c i n의 영향
F L S를 배지단독, TGF-β(10 ng/㎖), TGFβ
( 1 0 n g /㎖)+IDC(1~100 μM )의 조건하에서 6시간 배양하였고, 세포를 수집한 후 얻어진 총 R N A를 방법에서 제시한 대로 RT-PCR 하였다. TGF-β (10 ng/㎖)를 처리한 경우(lane 2)의 C O X - 2 mRNA 표현은 처리하지 않을 경우(lane 1)보다 3 배 증가하였고, 여기에 I D C를 처리한 경우 약물 농 도가 증가함에 따라 COX-2 mRNA 활성도가 농도 의존적으로 감소하였다(그림. 3). 선택적 C O X - 2 억제제인 NS398 역시 I D C와 유사한 결과를 보였다 (결과 생략).
4. VEGF mRNA발현에 대한 i n d o m e t h a c i n의 영향 I D C에 의한 V E G F의 생성감소가 전사과정 (transcriptional level)에서 조절되는가 알아보기 위해 F L S를 배지단독, TGF-β(10ng/ml), TGFβ ( 1 0 n g / m l )+I D C ( 1 ~ 1 0 0μM )의 조건하에서 6시간 배양하였고, 그 후 얻어진 총 R N A를 방법에서 제 시한 대로 RT-PCR 하였다. TGF-β( 1 0 n g / m l )만 처리한 경우(lane 2)의 VEGF mRNA 활성도는 아무것도 처리하지 않은 경우(lane 1)보다 2 . 6배 증 가하였고, 여기에 I D C을 처리한 경우 약물 농도가 Fig. 1. Effect of cytokines on VEGF production
from FLS. FLS were cultured in the pres- ence or the absence of cytokines, TGF-β ( 1 0 n g /㎖), IL-1β( 1 0 n g /㎖), TNF-α( 1 n g /㎖) , and IFN-γ( 1 0 0 0 U /㎖) for 24h. The amount of VEGF in supernatants were determined by ELISA. Data are expressed as means±
SD in three experiments
Fig. 2. Effect of indomethacin(IDC) on VEGF production from FLS. FLS were culture for 24h with different dose of IDC(1 to 100 μM) in the absence or presence of T G F -β( 1 0 n g /㎖). The amount of VEGF in supernatants were determined by ELISA.
Data are expressed as means±SD in three experiments. *Mann-Whitney test, p
<0.005 compared to culture media only
증가함에 따라 VEGF mRNA 활성도가 농도 의존 적으로 감소하였다(그림 4). 선택적 COX-2 억제제 인 NS398 역시 I D C와 유사한 결과를 보였다(결과 생략).
고 찰
신생혈관형성(angiogenesis, neovasculariza- t i o n )은 이미 존재하는 혈관에서 새로운 혈관이 생성 되는 복잡한 과정이다. 생리적 반응으로 일어나는 경
우, 이는 배아형성, 성장, 조직 재생에 필수적인 요소 이다. 그러나 병적으로 일어날 경우의 신생혈관형성 은 기관 손상, 당뇨병성 망막병증에서의 실명, 종양의 성장 및 전이에 중심적 역할을 한다4 ). 신생혈관이 형 성되기 위해서는 혈관내피세포의 이동과 증식, 이에 따른 모세혈관형성, 기저막의 침착, 평활근세포 및 주 위세포의 이동과 증식이 필수적이다. 이러한 복잡한 과정에는 각 단계마다 신생혈관형성을 위한 촉진 또 는 억제인자가 필요하다. 지금까지 알려진 신생혈관 형성 억제인자로는 leukemia inhibitory factor, Fig. 3. Effect of indomethacin(IDC) on COX-2
mRNA expression. FLS were cultured for 6 hours in media alone (lane1), 10ng/ml of TGF-β(lane 2), 10ng/ml of TGF-βa n d 1 μM of IDC (lane 3), 10ng/ml of TGF-β and 10 μM of IDC (lane 4), or 10ng/ml of T G F -βand 100 μM of IDC (lane 5). Total RNA was isolated and 5 ㎍ of RNA was reverse transcribed. Then RT-PCR was performed as described in materials and methods. Similar results are obtained from three experiments
Fig. 4. Effect of indomethacin (IDC) on VEGF mRNA expression. FLS were cultured for 6 hours in media alone(lane1), 10 ng/㎖ o f T G F -β(lane 2), 10ng/ml of TGF-βand 1μ M of IDC(lane 3), 10ng/ml of TGF-βa n d 1 0μM of IDC (lane 4), or 10ng/ml of TGF- βand 100μM of IDC (lane 5). Total RNA was isolated and 5 ㎍ of RNA was reverse transcribed. Then RT-PCR was performed as described in materials and methods.
Similar results are obtained from three e x p e r i m e n t s
Table 1. Primer sequence for PCR amplication of COX-2, VEGF, and GAPDH cDNA
N a m e Sequence (5
’
- 3’
)sense primer antisense primer
C O X - 2 5
’
- G C A G T T G T T C C A G A C A A G C A - 3’ 5’
- C A G G A T A C A G C T C C A C A G C A - 3’V E G F 5
’
- T C T T G G G T G C A T T G G A G C C T C - 3’ 5’
- A A G C T C A T C T C T C C T A T G T G C - 3’G A P D H 5
’
- C G A T G C T G G G C G T G A G T A C - 3’ 5’
- C G T T C A G C T C A G G G A T G A C C - 3’thrombospondin, IFN-γ, tissue inhibitors of metalloproteinase (TIMP)-1 또는 - 2가 있다4 ). 급 성기 때의 신생혈관형성 촉진인자로는 b r a d y k i n i n , substance P, calcitonin gene-related peptide, angiotensin Ⅱ, PGE2, nitric oxide, histamine 등이 있고, 만성기 때의 촉진인자로는 VEGF, basic FGF, IL-1, IL-8, angiogenin, TGF-β, TNF-α, PDGF 등이 있다4 ). 이중 가장 강력한 혈관생성 촉진 인자로 알려진 VEGF/vascular permeability fac- tor (VPF)는 종양발생 및 염증반응에 필요한 혈관내 피세포에 존재하는 두 개의 VEGF/VPF 수용체 (Flt-1, Flk-1/KDR)를 통해서 신생혈관형성에 관여 한다고 알려져 있다1 6 ).
류마티스 관절염은 증식된 활막세포와 혈관으로 구 성된 활막의 현저한 확장을 특징으로 하는 신생혈관 형성 의존성 질환 중의 하나이다. 따라서 혈관형성인 자의 조절은 섬유혈관육아조직내 내피혈관의 수를 감 소시켜 섬유혈관육아조직의 성장을 제한시킴으로써 연골 및 골미란으로 인한 관절파괴를 억제한다. 그러 므로 신생혈관형성을 억제하는 약물은 질환의 진행을 조절하는 새로운 접근이 될 수 있을 것이다.
기저상태에서의 C O X - 1은 대부분의 조직(위장관 점막세포, 혈소판, 혈관내피세포, 신수질 집합관과 간질세포)에서 내재적으로 발현되는 “h o u s e k e e p- ing enzyme”으로 위점막의 결집, 혈소판의 정상기 능, 신장내 혈류조절과 같은 항상성 유지에 필요한 프로스타글란딘 생성에 필수적이다. 또한 C O X - 1은 종양조직에도 발현된다1 3 ). 반면에 C O X - 2는 발현이 신속하고 엄격히 조절되는 “i m m e d i a t e - e a r l y e n z y m e”의 역할을 한다. COX-2는 생리적 조건의 정상상태에서는 뇌와 신피질을 제외한 대부분의 조 직에서 발견되지 않으나, 염증반응이 있는 경우 여 러 세포-활막세포, 혈관내피세포, 연골세포, 골수아 세포, 단핵구, 대식세포-에서 분비되는 싸이토카인 (IL-1, TNF-α)이나 유사분열물질( m i t o g e n )에 의 해서 신속히 발현된다1 3 , 1 7 ). 정상적으로 C O X - 2를 발 현하는 세포는 뇌의 해마, pyriform cortex의 pyramidal cell, amygdala complex, 신장의 juxtaglomerular apparatus의 macular densa와 H e n l e씨 고리의 상행각에 위치한다1 7 ). 또한 C O X - 2 는 일부 종양세포 및 종양조직에서 그 발현이 증가 되어 있다. 종양발생과 COX-2 발현과의 관계에는
세포사멸에 대한 저항성이 관여할 것으로 생각된다.
종양내 신생혈관 및 종양세포에 발현되는 C O X - 2에 대한 억제제는 종양발생 예방약제로서1 4 , 1 5 ) 뿐만 아니 라, 종양(대장암, 유방암, 전립선암, 폐암)치료에도 유용할 것으로 보인다1 3 , 1 8 - 2 0 )
. Masferrer 등에1 3 )의하 면, COX-2는 다양한 종양의 혈관내피세포, 종양상 피세포, 실질세포, 파골세포, 일부 염증세포에서 발 현되고, 선택적 COX-2 억제제는 신생혈관형성과 종양생성를 억제한다고 보고하였다. 이런 실험의 결 과로 혈관내피세포에 존재하는 C O X - 2가 신생혈관 형성을 억제시키는 치료의 목표물임을 알 수 있다.
Nagashima 등은2 1 ) 항류마티스약제( d i s e a s e modifying anti-rheumatic drugs) 중의 하나인 b u c i l l a m i n e이 류마티스 관절염 환자의 배양된 활막 세포에서 V E G F생성 감소를 통해 신생혈관형성 및 활막세포 증식을 억제한다고 보고하였다. 류마티스 관절염에서 N S A I D s는 소염 및 진통을 목적으로 사 용되고 있지만 류마티스 관절염의 경과에 중요한 골 미란 및 관절변형에는 도움이 되지 않는 것으로 알려 져 왔다. 그러나 일부 보고에 의하면2 2 - 2 5 ), NSAIDs 는 adjuvant arthritic rat model에서 프로스타글 란딘 생성억제를 통해 증식성 활막염, 관절종창, 연 골 및 골미란을 억제하는 등 부분적으로 관절손상을 막아주는 질환조절(disease-modifying) 효과가 있 음이 보고되고 있다. Katori 등2 6 )은 급성 삼출성 또 는 증식성 염증반응 모델을 이용하여 육아종 형성과 미세혈관 형성시 C O X - 2의 발현과 PGE2 생성이 증 가됨을 설명하였고, 이러한 현상들은 N S A I D s에 의 해 억제됨을 보고하였다. 이를 이용하여 활막액의 증 가, 섬유혈관육아조직 증가, 골흡수 증가, 신생혈관 형성을 특징으로 하는 류마티스 관절염의 치료에 N S A I D s가 사용될 수 있음을 설명하였다.
본 연구에서는 류마티스 활막염에 관여하는 염증 유발 물질인 T G F -β, IL-1β, TNF-α, IFN-γ등의 자극에 의해 V E G F생성이 증가됨을 알 수 있었다.
또한 N S A I D s인 i n d o m e t h a c i n의 농도가 증가함에 따라 COX-2 및 VEGF mRNA 발현이 억제되어 V E G F생성이 감소됨을 알 수 있었다. 이상의 결과 로 N S A I D s는 신생혈관형성, 활막액의 증가, 골흡 수에 중심적 역할을 하는 V E G F생성 감소를 통해 류마티스 관절염의 진행 및 영속성에 중요한 신생혈 관형성의 억제에 관여함을 알 수 있었다. 비특이적
비스테로이드성 항염제의 부작용을 최소화하고 활막 염 발생 및 진행에 관여하는 C O X - 2에 대한 선택적 억제제는 류마티스 관절염의 경과를 조절할 수 약제 로 사용될 수 있을 것으로 생각된다.
류마티스 활막조직에서 C O X에 의해 주로 생성되 는 프로스타글란딘은 P G E2, PGF2 a, thromboxa- men A2, prostacyclin이 있으며, 이중 연골 및 골 미란과 관련된 신생혈관형성에 관여하는 것은 P G E2
로 알려져 있다. Feldmann 등2 7 )은 활막세포에서 P G E2에 의해 V E G F가 발현됨으로써, PGE2가 신 생혈관형성을 유도할 수 있다고 하였다. 또한 P a z i t
등2 8 )은, PGE2가 활막세포 표면에 표현되어 있는
EP2 수용체와 결합하면 protein kinase A path- w a y가 활성화되어 VEGF mRNA발현이 증가됨을 보고하였다.
앞으로 N S A I D s에 의한 V E G F생성 감소가 단순 한 COX-2 활성도 억제로 인한 프로스타글란딘 생 성 감소로 인한 것인지, 아니면 또 다른 기전- VEGF mRNA 발현에 필요한 전사인자( N F -κB , A P - 1 )조절이 함께 관여하는지 연구가 필요할 것으 로 사료된다.
결 론
비스테로이드성 항염제는 류마티스 관절염의 활막 염과 통증에 대한 효과에 대해서는 널리 알려져 있 지만, 활막염의 진행억제에 관여하는 인자에 대한 연구는 많지 않았다. 또한 비스테로이드성 항염제가 가지고 있는 사용상의 부작용-위궤양, 위출혈, 부종, 혈액응고장애로 인해 장기간 사용하는데 제한적인 요소가 있었다. 본 연구에 의하면, 최근 많이 사용되 고 있는 C O X - 2를 선택적으로 억제하는 비스테로이 드성 항염제는 단순한 염증억제 뿐 만 아니라 신생 혈관형성을 차단하여 류마티스 관절염의 질병의 경 과를 변화시킬 수 있을 것으로 생각된다.
R E F E R E N C E S
11 ) Colville-Nash PR, Scott DL. Angiogenesis and
rheumatoid arthritis:pathogenic and therapeutic implications. Ann Rheum Dis 1992;51:919-25.12 ) Stephens RW, Ghosh P, Taylor TK. The patho-
genesis of osteoarthrosis. Med Hypotheses 1 9 7 9 ; 5 : 8 0 9 - 1 6 .
13 ) Paleolog EM. Angiogenesis: A critical process
in the pathogenesis of RA-a role for VEGF ? Br J Rheumatol 1996;35:917-9.14 ) Walsh DA. Angiogenesis and arthritis.
Rheumatology 1999;38:103-12.
15 ) Alisa EK, Lisa AH, George KH, Edward PA,
Flaine NUi, Wei LW, et.al. Vascular endothe- lial growth factor:A cytokine modulating endothelial function in rheumatoid arthritis. J Immunol 1994;152:4149-56.16 ) Lee SS, Joo YS, Kim WU, Park SH, Cho CS,
Kim HY, et.al. Vascular endothelial growth fac- tor level in the serum and synovial fluid of patients with rheumatoid arthritis. Clin Exp Rheumatol 2001;19:321-24.17 ) Cho CS, Kim WU, Min DJ, Park SH, Choe J,
Kim HY, et. al. CD40 engagement on synovial fibroblast up-regulates production of vascular endothelial growth factor. J Immunol 2000;164:5 0 5 5 - 6 1 .
18 ) Crofford LJ, Wilder RL, Ristimaki AP, Sano
H, Remmers EF, Hla T, et.al. Cyclooxygenase- 1 and -2 expression in rheumatoid synovial tis- sues. J Clin Invest 1994;93:1095-101.19 ) Angel J, Berenbaum F, Denmat C, Nevalainen
T, Masliah J, Fournier C. Interleukin-1- induced prostaglandin E2 biosynthesis in human synovial cells involves the activation of cytosolic phospholipase A2 and cyclooxygenase- 2. Eur J Biochem 1994;226:125-31.1 0 ) Chinery R, Beauchamp RD, Shyr Y, Kirkland SC, Coffey RJ, Morrow JD. Antioxidants reduce cyclooxygenase-2 expression, prostaglandin pro- duction, and proliferation in colorectal cancer cells. Cancer Res 1998;58:2323-7.
1 1 ) Sheng H, Shao J, Kirkland SC, Isakson P, Coffey RJ, Morrow J, et.al. Inhibition of human colon cancer cell growth by selective inhibition of cyclooxygenase-2. J Clin Invest 1 9 9 7 ; 9 9 : 2 2 5 4 - 9 .
1 2 ) Sawaoka H, Tsuji S, Tsujii M, Gunawan ES, Saski Y, Kawano S, et.al. Cyclooxygenase inhibitors suppress angiogenesis and reduce tumor growth in vivo. Lab Invest 1999;79:1469- 7 7 .
1 3 ) Masferrer JL, Leahy KM, Koki AT, Zweifel BS, Settle SL, Woerner BM, et.al. Antiangio-
genic and antitumor activities of cyclooxyge- nase-2 inhibitors. Cancer Res 2000;60:1306-11.
1 4 ) Watson AJ. Chemopreventive effects of NSAIDs against colorectal cancer: regulation of apoptosis and mitosis by COX-1 and COX-2.
Histol Histopathol 1998;13:591-7.
1 5 ) Tsujii M, Kawano S, Tsujii S, Sawaoka H, Hori M, DuBois RN. Cyclooxygenase regulates angiogenesis induced by colon cancer cells. Cell 1 9 9 8 ; 9 3 : 7 0 5 - 1 6 .
1 6 ) Dvorak HF, Detmar M, Claffey KP, Nagy JA, van de Water L, Senger DR. Vascular perme- ability factor/vascular endothelial growth fac- tor:an important mediator of angiogenesis in malignancy and inflammation. Int Arch Allergy Immunol 1995;107:233-5.
1 7 ) Crofford LJ. COX-1 and COX-2 tissue expres- sion implications and predictions. J Rheumatol 1997;(suppl 49)24:15-9.
1 8 ) Decensi A, Costa A. Recent advances in cancer chemoprevention with emphasis on breast and colorectal cancer. Eur J Cancer 2000;36:694- 7 0 9 .
1 9 ) Langman MJ, Cheng KK, Gilman EA, Lan- cashire RJ. Effect of anti-inflammatory drugs on overall risk of common cancer:case-control in general practice research database. BMJ 2000;
3 2 0 : 1 6 4 2 - 6 .
2 0 ) Wechter WJ, Leipold DD, Murray ED Jr, Quiggle D, MxCracker JD, Barrios RS, et. al.
E-7869 (R-flurbiprofen) inhibits progression of prostate cancer in the TRAMP mouse. Cancer Res 2000;60:2203-8.
2 1 ) Nagashima M, Yoshino S, Aono H, Takai M,
Sasanc M. Inhibitory effects of anti-rheumat- ic drugs on vascular endothelial growth factor in cultured rheumatoid synovial cells. Clin Exp Immunol 1999;116:360-5.
2 2 ) Ackerman NR, Rooks WH, Shott L, Genant H, Maloney P, West E. Effects of naproxen on connective tissue changes in the adjuvant arthritic rat. Arthitis Rheum 1979;22:1365-74.
2 3 ) Peakock DJ, Banquerigo ML, Brahn E. A novel angiogenesis inhibitor suppresses rat adju- vant arthritis. Cell Immunol 1995;160:178-84.
2 4 ) Weichman BM, Chau TT, Rona G. Histologic evaluation of the effects of etodolac in estab- lished adjuvant arthritis in rats:evidence of reversal of joint damage. Arthritis Rheum 1987;
3 0 : 4 6 6 - 7 0 .
2 5 ) Ackerman NR, Kappas K, Maloney P. Possible disease-modifying effects of naproxen in the adjuvant-arthritic rat. Agents Actions 1984;15:
4 2 8 - 3 5 .
2 6 ) Katori M, Majima M, Harada Y. Possible back- ground mechanism of the effectiveness of cyclooxygenase-2 inhibitors in the treatment of rheumatoid arthritis. Inflamm Res 1998;(suppl 2 ) 4 7 : S 1 0 7 - 1 1 .
2 7 ) Feldmann M, Brennan FM, Maini RN. Role of cytokines in rheumatoid arthritis. Annu Rev Immunol 1996;14:397-440.
2 8 ) Pazit BA, Crofford LJ, Wilder RL, Hla T.
Induction of vascular endothelial growth factor expression in synovial fibroblasts by prostaglandin E and interleukin-1 : a potential mechanism for inflammatory angiogenesis.
FEBS Letters 1995;372:83-7.