232 책임저자:최영현, ꂕ 614-052, 부산시 부산진구 진리 1로 100
동의대학교 한의과대학 생화학교실 Tel: 051-850-7413, Fax: 051-853-4036 E-mail: [email protected]
접수일:2005년 9월 28일, 게재승인일:2005년 10월 24일
Correspondence to:Yung Hyun Choi
Department of Biochemistry, Dongeui University College of Oriental Medicine, 100 Jilli-1 lo, Busanjin-gu, Busan 614-052, Korea
Tel: +82-51-850-7413, Fax: +82-51-853-4036 E-mail: [email protected]
신령버섯 추출물에 의한 A549 인체 폐암세포의 Apoptosis 유발
동의대학교 한의과대학 1생화학교실, 5해부학교실, 바이오물질제어학과 및 한의학연구소, 부산대학교
자연과학대학 2생물학과 및 3미생물학과, 4제주대학교 해양과학대학 해양과학부,
6동의대학교 자연과학대학 응용생명과학과
최우영1,2․박 철1․이재윤3․김기영4․최병태5․이원호2․정영기6․최영현1
Apoptotic Cell Death by Water Extract of Agaricus blazei Murill in Human Lung Carcinoma Cell Line A549
Woo Young Choi1,2, Cheol Park1, Jae-Yoon Lee3, Gi-Young Kim4, Byung Tae Choi5, Won Ho Lee2, Yong-Kee Jeong6 and Yung Hyun Choi1
Departments of 1Biochemistry, 5Anatomy, Biomaterial Control and Research Institute of Oriental Medicine, Dongeui University College of Oriental Medicine, Busan 614-052, Departments of 2Biology, 3Microbiology,
College of Natural Sciences, Pusan National University, Busan 614-714, 4School of Applied Marine Science, College of Ocean Science, Cheju National University, Jeju-do 690-756, 6Department of Microbiology,
College of Nutural Science, Dongeui University, Busan 609-735, Korea
Agaricus blazei Murill is a traditional herb medicine which has been used for patient suffering from cancer in Oriental medicine. It used to be a source of antitumor and immunoactive compounds in Oriental medicine, and considered a health food in many countries. In the present study, it was examined the biochemical mechanisms of apoptosis by water extract of Agaricus blazei Murill (WEAB) in human lung carcinoma cell line A549. It was found that WEAB could inhibit the cell growth of U937 cells in a dose-dependent manner, which was associated with apoptotic cell death such as formation of apoptotic bodies and DNA fragmentation. Flow cytometry analysis confirmed that WEAB treatment increased populations of apoptotic-sub G1 phase. Apoptosis of A549 cells by WEAB was associated with a down-regulation of anti-apoptotic Bcl-2 expression and an up-regulation of pro-apoptotic Bax expression.
WEAB treatment induced the proteolitic activation of caspase-3 and caspase-9, and a concomitant degradation of poly (ADP-ribose) polymerase (PARP) and β-catenin protein. Taken together, these results indicated that the anti-proliferative effects of WEAB were associated with the induction of apoptotic cell death through regulation of several major growth regulatory gene products such as Bcl-2 family expression and caspase protease ativity, and WEAB may have therapeutic potential in human lung cancer treatment. (Cancer Prev Res 10, 232-241, 2005)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Key Words: Agaricus blazei Murill, Apoptosis, Bax, Caspase
서 론
1940년대 초반에 미국에서 처음 발견되었으며, 미국 의 플로리다와 중남미의 중원지대에 자생되는 버섯으로 최근 국내에서 재배되기 시작한 신령버섯(Agaricus blazei Murill)은 분류학상으로 균계, 진균문, 담자균아문, 주름 버섯목, 주름버섯과의 주름버섯속에 속한다. 이는 양송 이와 같은 속으로서, 송이버섯, 팽이버섯, 느타리버섯과 같은 목에 속하는 버섯으로서 강력한 면역증강 활성물 질이 함유되어 있다고 알려져 있다.1) 최근까지 많은 연 구가 일본이나 중국의 학자들에 의해서 이루어져 왔는 데, 특히 일본의 학자들은 중남미의 잉카지역 주민들이 암 관련 질환이나 각종 성인병환자가 적으며 장수하는 사람이 유난히 많다는 점에 주목하고, 이 지역주민들의 장수원인에 대한 비밀을 연구한 결과, 그 원인이 그 지역 주민들이 즐겨 식용하는 버섯에 있음을 밝혀졌다.2,3) 신 령버섯의 주요 효과에는 항종양, 면역증강, 항염작용, 혈 당 강하 작용, 강심작용, 혈압 강화 작용, 콜레스테롤 저 하 작용, 치매개선 효과, 비만억제 효과, 고혈압, 고지혈 증 등의 성인병과 미용효과가 있는 것으로 연구발표 되 고 있으며,4∼6) 건강보조식품 등 여러 다양한 분야에서 각광을 받고 있어서 앞으로 더욱 많은 관심을 야기시킬 것으로 생각된다.7,8)
세포의 여러 가지 활동 중 세포의 죽음은 necrosis와 apoptosis로 구분되며, 이것은 세포의 형태학적 및 생화학 적인 특성에 의하여 구분될 수 있다. Apoptosis는 개체의 발생단계나 DNA 손상, 바이러스 감염 등에 의한 유전적 조절 하에서 일어나는 정교한 방어기전이라는 점에서 생리적이거나 화학적인 외상에 의한 세포의 죽음인 ne- crosis와는 구별된다.9∼11) 또한 apoptosis는 개체보존의 수 준에서 손상된 세포들의 제거를 위한 중요한 수단이기 도 하다. Apoptosis의 유발에 종양억제유전자 p53, Bcl-2 family 및 caspase member 등과 같은 유전자가 관여한다는 사실이 알려지면서 apoptosis와 연관된 분자적 기전이 최 근 많이 밝혀지고 있는데, Bcl-2 family에 속하는 Bcl-2는 apoptosis를 억제하는 반면, Bax의 과발현이 이루어졌을 때는 apoptosis를 유발시킨다고 알려져 있다.12) 그리고 caspase라고 이름 붙여진 ICE/CED-like protease family 역시 apoptosis 유발에 중요한 역할을 한다고 알려져 있으며, 이들은 proenzyme 형태로 존재하다가 apoptosis 유도를 활 성화시키는 신호에 의해 활성화된 cysteine-related prote- ases로 되어 직접 또는 간접으로 세포 내에 존재하는 많 은 표적 단백질의 분해에 관여하게 된다.13∼15)
본 실험에서는 인체 폐암세포의 증식에서 신령버섯 추출물에 의한 암세포 성장억제 효과와 apoptosis의 유발 에 관한 기전 해석의 시도를 위하여 암세포의 성장 및 생존율 억제, 이로 인한 세포와 핵의 형태적 변화와 DNA fragmentation 등을 조사하였으며, apoptosis의 유발 과 연관성을 가지는 주요 유전 인자들의 발현 변화를 조 사하였다.
재료 및 방법 1. 암세포배양 및 시료의 처리
본 연구에 사용된 A549 인체 폐암세포는 한국생명공 학연구소(KRIBB, Taejeon, Korea)에서 분주 받아 사용하였 으며, 암세포의 배양을 위해 90%의 RPMI-1640 배지 (Gibco BRL, Grand Island, NY, USA), 10% fetal bovine serum (FBS, Gibco BRL, Grand Island, NY, USA)에 1%의 penicillin 및 streptomycin (Gibco BRL, Grand Island, NY, USA)이 포함 된 성장배지를 사용하여 37oC, 5% CO2 조건 하에서 배양 하였다. 본 실험에 사용된 신령버섯(Agaricus blazei Murill) 은 김해시 소재 청원농산에서 제공받았으며, 100 g을 1,000 ml의 증류수에 3시간 이상 끓인 후, 3,000 rpm으로 20분간 원심 분리시켜 침전물을 제거하였다. 이를 다시 0.45 um의 여과지를 이용하여 부유 성분을 걸러낸 후 수 용성분을 동결 건조하여 사용하였다. 준비된 신령버섯 수용액 추출물(water extracts of A. blazei Murill, WEAB)은 3차 증류수에 용해하여 멸균과 여과 과정을 거쳐 배지에 적정 농도로 희석하여 처리하였다.
2. Hemocytometer를 이용한 세포 생존율의 측정
세포 배양용 6 well plate에 인체 폐암세포를 3×104개/
ml의 개수로 well 당 2 ml씩 분주하고 24시간 동안 안정 화시킨 다음 WEAB를 배지에 희석하여 각 well당 1∼5 mg/ml의 농도로 처리한 후 배양하였다. 48시간 후 배지 를 제거하고 0.05% trypsin-EDTA를 처리하여 세포를 부 유시킨 다음 phosphate-buffered saline (PBS)를 각 well 당 적정량을 첨가하여 세포를 모은 다음, 2,000 rpm으로 5분 간 원심 분리하였다. 상층액을 제거하고 세포만 남긴 다 음 다시 PBS를 1 ml 첨가하여 잘 섞은 후 세포 부유액과 0.5% trypan blue (Gibco BRL)를 동량으로 섞은 후 2분간 처리한 후 위상차 현미경(Carl Zeiss, Germany)을 이용하여 살아있는 세포를 계수하였다. 이에 따른 결과를 Microsoft EXCEL program을 사용하여 분석하였다.
3. DAPI staining에 의한 세포핵의 형태 관찰
WEAB 처리에 의한 암세포의 apoptosis 유발 여부 확인 을 위한 핵의 형태적 변화를 관찰하기 위하여 준비된 세 포를 모은 다음 37% formaldehyde 용액과 PBS를 1 : 9의 비율로 섞은 fixing solution을 모아진 세포에 500μl 첨가 하여 잘 섞어준 후, 실온에서 10분 동안 고정하였다.
1,000 rpm으로 5분간 원심 분리한 후 상층액을 제거하고 PBS 200μl를 넣어서 충분히 섞은 후, slide glass 위에 80μ l 정도 떨어뜨려 900 rpm에서 5분간 cytospin하였다. PBS 로 2∼3회 washing하고 PBS가 다 마르기 전에 0.2%의 Triton X-100을 첨가하여 실온에서 10분간 고정하였다.
그 후 다시 PBS로 washing하고 4’,6-diamidino-2-phenylin- dole (DAPI, Sigma, St. Louis, MO, USA) 용액을 세포가 고 정된 slide glass 위에 적당량을 떨어뜨린 후 빛을 차단하 고 실온에서 염색시켰다. 15분 가량 염색시킨 후, PBS로 DAPI 용액을 충분하게 washing하고 형광 현미경(Carl Zeiss)을 이용하여 400배의 배율로 각 WEAB 처리 농도에 따른 인체 폐암세포의 핵의 형태 변화를 관찰하였다.
4. DNA flow cytometry에 의한 분석
정상 및 WEAB를 처리한 배지에서 48시간 동안 배양 시킨 암세포를 PBS로 씻어 내고 0.05% trypsin-EDTA를 처리하여 부유시킨 다음 1,000 rpm으로 10분간 원심 분 리하여 상층액을 버리고 세포들만 모았다. 여기에 다시 PBS를 첨가하여 충분히 씻은 다음 1,000 rpm으로 10분간 원심 분리한 후 상층액만 버리고 남은 세포에 0.5 ml의 PBS로 잘 부유시키고, 차가운 ethanol 0.5 ml을 첨가하여 4oC에서 한 시간 동안 고정시켰다. 5×106개의 고정된 세 포들을 원추형 vial에 넣어서 1,000 rpm으로 수분간 원심 분리하여 상층액을 제거하고, 1% bovine serum albumin (BSA, Sigma)이 함유된 PBS로 2∼3회 washing 과정을 거친 후 다시 수 분간 원심 분리하였다. 세포 침전물을 1%
BSA를 함유한 PBS 0.8 ml로 부유시키고 DNA intercalating dye propidium iodide (PI, concentration, 50μg/ml; Sigma)와 0.1 mg/ml의 RNase (Sigma)를 처리하여 암실(4oC)에서 1시 간 동안 염색과정을 거쳤다. PBS로 두 번 washing 과정을 거친 후, 부유액을 만들고, 35μm pore size의 nylon mesh에 부유액을 pipette으로 통과시켜 단일 세포로 분리시킨 후 DNA flow cytometry (Becton Dickinson, San Jose, CA, USA) 에 적용시켜 형광반응에 따른 histogram을 ModiFit LT (Becton Dickinson) program을 사용하여 분석하였다.
5. DNA fragmentation의 분석
Apoptosis가 유발된 세포에서 관찰될 수 있는 DNA fragmentation의 분석을 위하여 정상 및 WEAB가 처리된 배지에서 48시간 동안 배양된 세포를 모아 lysis buffer [5 mM Tris-HCl (pH 7.5), 5 mM EDTA, 0.5% Triton X-100]를 4oC에서 30분간 처리하였다. 그 후 14,000 rpm에서 20분 간 원심분리하고, 그 상층액에 proteinase K (Sigma)를 0.5 mg/ml의 농도로 처리하였다. 그 후 phenol:chloroform:
isoamyl alcohol 혼합 용액(25:24:1, Sigma)을 첨가하였 다. 30분간 rotate시킨 다음 14,000 rpm에서 10분간 원심 분리하였다. 여기서 얻어진 상층액에 적정량의 isopro- panol (Sigma)과 5 M NaCl를 첨가하였다. 24시간 정도 냉 장 보관한 후, 14,000 rpm (4oC)에서 30분간 원심분리 시 킨 후 상층액을 버리고, RNase A를 이용하여 pellet을 녹 이고 여기에 6X gel loading dye (Bioneer, Daejeon, Korea)를 섞어 주었다. 1.5% agarose gel을 만들어서 1시간 가량 50 V로 전기영동 시킨 후 ethidium bromide (EtBr, Sigma)로 염색하여 ultra vilolet (UV)하에서 사진 촬영하였다.
6. RT-PCR에 의한 mRNA 발현의 분석
상기와 동일 조건에서 준비된 세포들을 모아 PBS로 washing한 후 TRIzol reagent (Invitrogen Co., Carlsbad, CA, USA)를 4oC에서 1시간 동안 처리하여 세포 내 총 RNA를 분리하였다. 분리된 RNA를 spectrophotometer를 이용하여 정량한 후 각각의 primer, DEPC water 그리고 ONE-STEP RT-PCR PreMix Kit (Intron, Korea)를 넣고 Mastercycler gradient (Eppendorf, Hamburg, Germany)를 이용하여 증폭 하였다. 각 PCR product의 양적인 차이를 확인하기 위하 여 1x TAE buffer로 1% agarose gel을 만든 후 각 well에 PCR product와 DNA gel loading solution (5x, Bioneer)을 섞 어서 loading한 후 100 V에서 전기영동으로 분리하였다.
분리가 끝난 gel을 EtBr을 이용하여 염색한 후 UV하에서 확인하고 Picture works photo enhancer program을 이용하여 촬영하였다. 본 연구에서 사용한 primer의 종류와 염기서 열은 Table 1에 나타낸 것과 같으며 glyceraldehyde-3- phosphate dehydrogenase (GAPDH) 유전자를 internal control 로 사용하였다.
7. Western blot analysis에 의한 단백질 발현의 분석
정상 및 WEAB가 처리된 배지에서 48시간 배양된 세 포들을 PBS로 washing하고 0.05% trypsin-EDTA를 처리하 여 부유시킨 다음 수분간 원심 분리하여 세포를 모았다.
이렇게 모은 세포에 적정량의 lysis buffer [25 mM Tris-Cl
(pH 7.5), 250 mM NaCl, 5 mM EDTA, 1% NP-40, 1 mM phenymethylsulfonyl fluoride (PMSF), 5 mM dithiothreitol (DTT)]를 500μl 첨가하여 4oC에서 30분간 반응시킨 후, 14,000 rpm으로 30분간 원심 분리하여 그 상층액을 취하 였다. 상층액의 단백질 농도는 Bio-Rad 단백질 정량 시약 (Bio-Rad, Hercules, CA, USA)과 그 사용방법에 따라 정량 한 다음 동량의 Laemmli sample buffer (Bio-Rad)를 섞어서 sample을 완성한 후, 분석하고자 하는 단백질의 분자량 에 따라 적정 농도의 sodium dodecyl sulphate (SDS)-polya- crylamide gel을 이용하여 전기영동으로 분리하였다. 분리 된 gel을 nitrocellulose membrane (Schleicher and Schuell, Keene, NH, USA)으로 electroblotting에 의해 2∼3시간 가량 전이 시켰다. 전체 단백질이 전이된 membrane에 비특이적 단 백질에 대한 blocking을 위해 10% skin milk를 함유한 PBS-T (0.1% Tween 20 in PBS) 용액으로 상온에서 1시간 incubation하고 PBS-T로 15분(5분씩 나누어 3번)정도 washing하였다. 준비된 membrane에 각각의 적정 항체를 처리하여 4oC에서 1시간 이상 반응시킨 다음 PBS-T로 15 분간 washing한 후 다시 5분씩 5번을 washing하였다. 그 후, 처리된 1차 항체에 대응하는 2차 항체(PBS-T로 1:
1000으로 희석하여 사용)를 처리하고 실온에서 1시간 반 응시켰다. 다시 PBS-T로 10분간 3번, 5분간 3번의 wash- ing을 하고 enhanced chemiluminoesence (ECL) 용액(Amers- ham Life Science Corp., Arlington Heights, IL, USA)을 처리 한 다음 암실에서 Kodak X-ray film에 감광시켜 특정단백 질의 양을 분석하였다.
8. In vitro caspase-3 및 caspase-9의 activity 측정
Caspase-3와 caspase-9의 in vitro 활성 측정을 위한 colori- metric assay kits는 CLONTECH Lab. (Palo Alto, CA, USA) 및 R&D Systems (Minneapolis, MN, USA)에서 각각 구입하 였으며, 제시된 방법에 준하여 활성의 증감 여부를 조사 하였다. 이를 위하여 정상 및 WEAB가 처리된 배지에서 48 시간 배양된 세포를 모은 뒤 단백질을 추출하고 정량 하여 각각 20μg의 단백질을 fluorogenic peptide 기질 100 uM이 함유된 extraction buffer [40 mM HEPES (pH 7.4), 20% glycerol (v/v), 1 mM EDTA, 0.2% NP-40 and 10 mM DL-DTT] 50μl에 혼합하였으며, microtiter plate에 다시 extraction buffer에 희석하여 각 sample 당 총 volume이 100 μl가 되게 하였다. 실험에 사용된 기질은 caspase-3의 경 우에는 Asp-Glu-Val-Asp (DEVD)-p-nitroaniline (pNA)이었 으며, caspase-9은 Leu-Glu-His-Asp (LEHD)-pNA였다. 준비 된 plate를 37oC에서 2시간 동안 incubation 시킨 후 VERSAmax tunable microplate reader를 이용하여 405 nm의 흡광도를 이용하여 반응의 정도를 측정하였다.
결과 및 고찰
1. WEAB가 인체 폐암세포의 증식에 미치는 영향
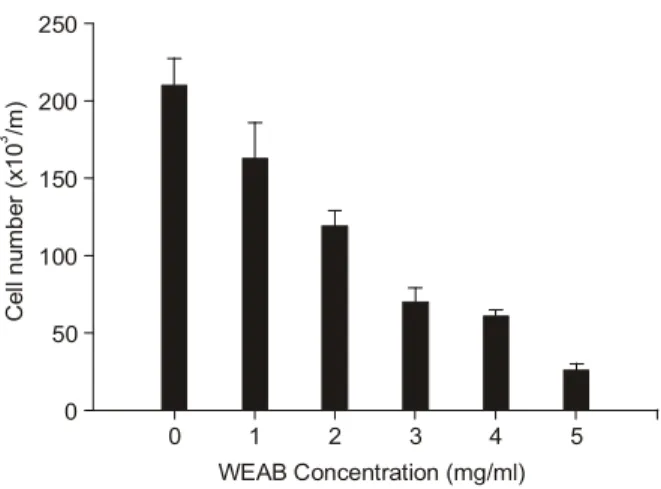
인체 폐암세포 A549의 세포 생존율에 WEAB가 각 처 리 농도별로 어떤 영향을 미치는지에 대해 hemocyto- meter를 이용한 조사를 통하여 분석한 결과는 Fig. 1에서 나타낸 바와 같다. 결과에서 알 수 있듯이 WEAB를 48시 Table 1. Gene-specific primers for RT-PCR
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Gene name Sequence
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Bax Sence 5’-ATG-GAC-GGG-TCC-GGG-GAG-3’
Antisence 5’-TGG-AAG-AAG-ATG-GGC-TGA-3’
Bcl-2 Sence 5’-CAG-CTG-CAC-CTG-ACG-3’
Antisence 5’-GCT-GGG-TAG-GTG-CAT-3’
Bcl-XL Sence 5’-CAG-CTG-CAC-CTG-ACG-3’
Antisence 5’-GCT-GGG-TAG-GTG-CAT-3’
XIAP Sence 5’-GAA-GAC-CCT-TGG-GAA-CAA-CA3’
Antisence 5’-CGC-CTT-AGC-TGC-TCT-CTT-CAG-T3’
cIAP-1 Sence 5’-TGA-GCA-TGC-AGA-CAC-ATG-C-3’
Antisence 5’-TGA-CGG-ATG-AAC-TCC-TGT-CC-3’
cIAP-2 Sence 5’-CAG-AAT-TGG-CAA-GAG-CTG-G-3’
Antisence 5’-CAC-TTG-CAA-GCT-GCT-CAG-G-3’
GAPDH Sence 5’-CGG-AGT-CAA-CGG-ATT-TGG-TCG-TAT-3’
Antisence 5’-AGC-CTT-CTC-CAT-GGT-GGT-GAA-GAC-3’
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
간 동안 처리한 결과 인체 폐암세포에서 처리농도 의존 적으로 세포의 생존율이 감소하였음을 알 수 있었다. 특 히 WEAB 처리 농도가 3 mg/ml 이상부터는 생존율이 50% 이상 억제되었으며, 최종 농도인 5 mg/ml에서는 세 포의 수가 정상적인 배지에서 배양된 세포보다 약 10%
정도로 줄어들었다. 따라서 WEAB를 처리하였을 경우 인체 폐암세포의 성장과 생존율에 처리 농도 의존적인 억제 효과가 있음을 알 수 있었다. 이는 선행 연구에서의 MTT assay 분석을 통한 세포증식 억제 조사에서 관찰된 것과 유사한 결과였다.16)
2. WEAB 처리에 의한 apoptosis의 유발
다음은 WEAB의 처리에 의한 인체 폐암세포의 생존율 감소 및 성장 억제가 apoptosis 유발에 연관성이 있을 것 으로 추측되어 WEAB를 48시간 동안 처리한 후 인체 폐 암세포의 각 세포주기를 비교하였다. 이를 위하여 각 농 도별로 WEAB를 처리한 후 PI염색을 이용하여 세포의 핵을 염색한 다음 DNA flow cytometry analysis를 이용하여 조사하였다. 이미 선행연구에 의해 WEAB는 인체 폐암 세포의 G2/M기의 arrest를 유발한다는 사실이 알려져 있 으며 형태학적 측면에서도 처리 농도 의존적으로 변화 가 있음을 검증하였다.16) Fig. 2C에 나타낸 바와 같이 apoptosis 유발 집단에 해당하는 sub-G1기가 WEAB처리 농도 의존적으로 매우 증가하였음을 알 수 있었다. 아울 러 apoptosis의 직접적인 증거를 제시하기 위하여 정상 및
WEAB가 처리된 배지에서 48시간 동안 배양한 인체 폐 암세포를 대상으로 DAPI staining을 통해 핵의 형태적인 변화를 조사하였다.9) Fig. 2A에서 나타난 바와 같이 WEAB가 처리된 배지에서 자란 A549 인체 폐암세포는 정상 배지에서 배양된 세포에서는 관찰할 수 없는 apo- ptosis 유발 특이적인 핵 내의 DNA 단편화에 의한 apo- ptotic body를 관찰할 수 있었다.11) 또한, WEAB의 처리 농도 의존적으로 A549인체 폐암세포의 밀도 역시 크게 감소하였으며, apoptotic body의 출현 빈도도 크게 증가하 였다. 이런 apoptotic body의 출현 빈도의 증가는 DNA flow cytometry analysis (Fig. 2C)의 결과에서의 sub-G1 집단 의 증가와도 유사하였다. 아울러 WEAB를 48시간 처리 한 후 세포내 총 DNA를 추출하여, DNA fragmentation 여 부를 분석하여 본 결과, 정상 배지에서 자란 인체 폐암세 포에 비해서 WEAB가 처리한 세포에서 DNA fragmenta- tion 유발현상을 어느 정도 관찰할 수 있었다(Fig. 2B). 이 는 선행 연구들의 결과에 의해 WEAB에 의해 인체 폐암 세포에서 apoptosis가 유발되었다는 또 다른 증거가 될 수 있으며,17) Fig. 2A의 DAPI staining에서의 결과에서 나타 난 apoptotic body 형성에 따른 결과로 생각된다. 따라서 WEAB 처리에 의한 인체 폐암세포의 증식 억제는 apo- ptosis와 직접적인 관련이 있음을 알 수 있었다.
3. Bcl-2 및 Bax의 발현에 미치는 WEAB의 영향
이상의 결과들을 근거로 하여 WEAB의 처리로 인한 apoptosis 유발 현상과 연관된 몇 가지 기본적인 기전 해 석을 위하여 RT-PCR과 western blotting을 통해 apoptosis 유발과 연관된 여러 인자들의 발현 변화를 조사하였다.
그 중 Bcl-2/Bax family에 속하는 유전자인 Bcl-2는 apo- ptosis 유발을 억제하는 anti-apoptotic 분자이며, Bax는 apoptosis 유발을 촉진하는 pro-apoptotic 분자이다.18,19) 이 두 유전자는 세포의 mitochodria로부터 cytochrome c를 유 리시켜 종양 억제 유전자인 p53, caspase 및 DNA 단편화 와 연관된 endonuclease 등의 활성을 조절한다.18,20,21) 이들 두 유전자는 dimer를 이루고 있는데 Bax의 발현이 증가 하고 Bcl-2의 발현이 감소하면 apoptosis가 유발될 수 있 는 것으로 알려져 있다.18,22) Fig. 3의 결과와 같이 Bax의 전사수준과 단백질 발현의 정도는 WEAB의 처리 농도 의존적으로 그 양이 증가하였으나 dimer를 이루는 Bcl-2 는 전사수준에서는 변화없이 단백질의 발현이 고농도 처리군에서는 다소 감소되었다. 따라서 WEAB 처리에 의한 인체 폐암세포의 apoptosis 유발에는 Bcl-2의 발현 억제 및 Bax의 발현 증가와 직접적인 연관성이 있는 것 으로 추정된다.
Fig. 1. Effects of water extracts of Agaricus blazei Murill (WEAB) on the viability of A549 human carcinoma cells. Cells were seeded at 3×104 per ml in a 6-well plate and incubated for 24 h. Cells were in the absence (0, control) or presence of increasing concentrations of WEAB for 48 h. The viable cell number was determined by hemocytometer counts of trypan blue-excluding cells. The data shown are means±SD of three independent expriments.
0
Cell number (x10/m)3
WEAB Concentration (mg/ml) 0
50 100 150 200 250
2 3 4 5
1
4. Caspase-3 및 caspase-9의 활성에 미치는 WEAB의 영향
Caspase는 세포의 apoptosis 유발에 핵심적인 역할을 하 는 인자로서 세포내의 핵과 mitochondria의 외막에 불활 성 상태로 존재하다가 apoptosis를 유도하는 자극에 의하 여 활성화될 수 있다.20) 따라서 caspase의 활성화는 apo- ptosis의 유발에 대한 또 다른 증거가 될 수 있으며 많은 선행연구 등에서 검증되어 왔다.23) 그 활성화의 정도를 in vitro caspase activity assay를 통하여 실험해본 결과는 Fig.
4와 같았다. 그 결과에서 알 수 있듯이 인체 폐암세포 A549에서 WEAB의 처리에 따른 농도 의존적으로 cas- pase-3 및 caspase-9의 활성 증가 현상을 관찰할 수 있었으 며, 특히 caspase-9보다 caspase-3의 활성이 더 크게 증가하 였음을 알 수 있었다.
5. PARP, β-catenin 의 발현에 미치는 WEAB의 영향
세포의 내부나 외부의 자극에 의해 apoptosis가 일어나 면 poly (ADP-ribose) polymerase (PARP) 및 β-catenin 등의 단백질이 부분적으로 잘리는 분해과정이 나타난다고 알 Fig. 2. WEAB induced apoptosis in A549 cells. (A) Cells were incubated variable concentrations of WEAB for 48 h, then sampled, fixed and stained with DAPI. After 10 min incubation at room temprature, stained nuclei were observed under a fluorescent microscope using a blue filter. Original magnification, ×400. (B) After WEAB treatment for 48 h, cells were collected and DNA was extracted.
The DNA fragmentations were separated on 1.5% agarose gel electrophoresis and visualized under UV light after staining with EtBr.
(C) Cells were exposed for 48 h to an increasing concentration of WEAB. Then the cells were collected and stained with PI for flow-cyrometric analysis. The percentages of cells with hypodiploid DNA contents represent the fractions undergoing apoptotic DNA degradation. Data are the mean of two different experiments.
C
C 1 3 5
WEAB (mg/ml)
C 1 2 3 4 5
Sub-G1 (%)
WEAB (mg/ml) 0
5 10 15 20 25 30
A
3 4 5
0 1 2
WEAB (mg/ml)
B
려져 있다.24,25) PARP 단백질은 정상적인 세포의 DNA repair나 genomic stability의 유지에 중요한 역할을 하며,26) apoptosis 과정 중 caspase의 활성에 의해 단백질 분해가 일어나게 된다면 PARP의 정상적인 repair 기능의 상실이 일어난다.14) 또한, β-catenin 단백질은 세포 내 골격유지 와 부착성 세포의 전사조절 및 세포유착에 관계된 apo- ptosis 조절과 관련이 있으며,27,28) β-catenin 단백질 역시 세포 유착성 apoptosis가 유발되면 caspase 활성과 연관되
어 단편화가 일어나는 것으로 알려져 있다.29,30) 따라서 WEAB 처리에 의한 apoptosis 유발에도 이러한 현상들이 관련되어 있는지의 여부를 조사하였다. Fig. 5의 결과에 서 알 수 있듯이 PARP와 β-catenin 단백질에서 특히 PARP가 빠르게 단편화된다는 것을 알 수 있었으며, 이 것은 WEAB의 처리 농도 의존적인 인체 폐암세포의 성 장억제가 caspase의 활성화에 의한 apoptosis의 유발과 직 접적인 관련이 있다는 사실을 보여주고 있는 것이다.
Fig. 4. Activation of caspase-3 and caspase-9 by WEAB treatment in A549 cells. Cells were treated with indicated concentrations of WEAB for 48 h, collected and then lysed. Aliquots (100 μg proteins) were incubated with DEVD-pNA (A) and LEHD-pA (B) for caspase-3 and caspase-9 activity, respectively, at 37oC for 1 h. The released fluorescent products were measured. Data represent the mean of two independent experiments.
A B
Relative caspase-9 activity
0 1 2 3 4 5
WEAB (mg/ml) 0
0.02 0.04 0.06 0.08 0.1 0.12
0 1 2 3 4 5
Relative caspase-3 activity
WEAB (mg/ml) 0
0.04 0.08 0.12 0.16 0.2
Fig. 3. Induction of Bax and inhibition of Bcl-2 and Bcl-XS/Lby WEAB in A549 cells. (A) After 48 h incubation with WEAB, total RNA was isolated and reverse-transcribed. The resulting cDNA was subjected to PCR with Bax, Bcl-2 and Bcl-XL primers and the reaction products were subjected to electrophoresis in 1% agarose gel and visualized by EtBr staining. GAPDH was used as an internal control. (B) The cells were lysed, and cellular proteins were separated by SDS-polyacrylamide gel and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control.
Bax
Bcl-2
Bcl-XL
GAPDH
Bax
Bcl-2
Bcl-XL
Actin
0 1 2 3 4 5
WEAB (mg/ml)
0 1 2 3 4 5
WEAB (mg/ml)
A B
6. IAP family의 발현에 미치는 WEAB의 영향
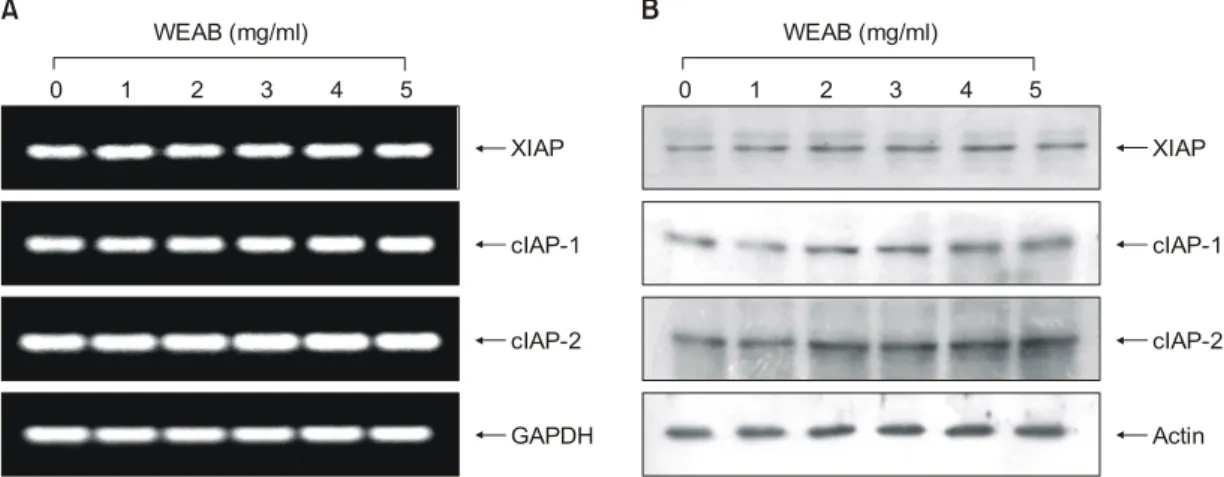
한편 apoptosis에 관여하는 여러 인자들 중 최근 밝혀진 또 다른 class인 inhibitor of apoptosis proteins (IAPs) family에 속하는 여러 유전자 산물은 곤충세포에서 바이러스 감 염에 의한 apoptosis를 억제하기 위한 baculoviral 단백질 군들로서 외부 신호에 의한 세포 내 anti-apoptotic 활성을 지닌다. 그리고 이들 중 어떤 IAPs는 caspases와의 직접적 인 결합을 통하여 그들의 apoptotic 활성을 억제할 수 있 을 것으로 밝혀져, IAP family들의 잠재적인 역할에 대한 관심이 높아지고 있다. 더구나 이들 IAP family의 발현은
전사 조절인자인 nuclear factor kappa B (NF-κB)에 의해 조절 받는 것으로 밝혀져 apoptosis의 신호전달 연구에서 의 새로운 영역을 제시하여 주고 있다.31∼33) WEAB에 의 한 apoptosis 유발에 이러한 cIAP family의 관여 여부를 조 사한 결과(Fig. 6), 조사된 IAP family 인자들(XIAP, cIAP-1 및 cIAP-2)의 mRNA 수준 및 단백질 발현 모두에서 WEAB 처리에 따른 뚜렷한 변화가 없었다.
결 론
강력한 항암 및 면역강화 작용을 가진 것으로 알려져 있는 물질 중의 하나인 신령버섯(Agaricus blazei Murill)의 수용성 추출물(WEAB)을 이용하여 암세포 성정 억제 기 전을 해석하기 위하여 인체 폐암세포 A549 세포주를 대 상으로 조사하였다. WEAB가 처리된 A549 인체 폐암세 포는 처리 농도 의존적으로 성장 및 생존율이 현저히 감 소되었으며, apoptosis가 유발된 세포에서 특징적으로 관 찰되는 chromatin condensation 현상을 유발하였고 DNA Flow cytometry 분석결과 apoptotic sub-G1기에 해당하는 세포들의 빈도가 처리농도 의존적으로 증가하였다. Bcl-2/
Bax family의 전사 및 번역수준에서 관찰한 결과 pro- apoptotic 인자인 Bax의 경우 mRNA 및 protein 수준에서 모두 발현이 증가되었지만 anti-apoptotic 인자인 Bcl-2의 경우 mRNA수준에서는 아무런 변화가 없었지만 protein 수준에서는 고농도 처리군에서 약간의 감소가 관찰되었 다. 또한, WEAB는 caspase-3 및 caspase-9의 활성 증가에
Fig. 6. Effects of WEAB on the levels of IAP family members in A549 cells. (A) After 48 h incubation with WEAB, total RNA was isolated and reverse-transcribed. The resulting cDNA was subjected to PCR with XIAP, cIAP-1 and cIAP-2 primers and the reaction products were subjected to electrophoresis in 1% agarose gel and visualized by EtBr staining. GAPDH was used as an internal control. (B) The cells were lysed, and cellular proteins were separated by SDS-polyacrylamide gel and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control.
XIAP
cIAP-1
cIAP-2
GAPDH Actin
0 1 2 3 4 5
WEAB (mg/ml)
0 1 2 3 4 5
WEAB (mg/ml)
XIAP
cIAP-1
cIAP-2
A B
Fig. 5. Effects of WEAB on the levels of PARP and β-catenin in A549 cells. The cells were lysed, and cellular proteins were separated by SDS-polyacrylamide gel and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control.
PARP
β-catenin
Actin
0 1 2 3 4 5
WEAB (mg/ml)
영향을 주었으며 특히 caspase-3의 활성에 큰 영향을 주 었다. 이런 caspase의 활성화로 인해 PARP와 β-catenin 단 백질의 분절현상 등이 관찰되었으며 이는 apoptosis가 유 발되었다는 또 다른 증거가 될 수 있다. 이상의 결과에서 WEAB에 의한 암세포의 생존율 저하는 apoptosis 조절에 중요한 유전자들의 선택적 발현 조절을 통한 apoptosis 유 발과 밀접한 관련이 있었으며, 이는 신령버섯 추출물이 강력한 항암 및 암예방 효능의 잠재력을 가지고 있음을 의미하며 지속적인 연구가 필요할 것으로 생각된다.
감사의 글
본 연구는 2002년도 농림부 농림기술 개발사업(첨단 기술개발분야)의 연구비 지원에 의해 이루어진 결과의 일부분이며 이에 감사드립니다.
참 고 문 헌
1) Ito H, Shimura H, Itoh M, Kaurode M. Antitumour effects of a new polysaccharideprotein complex (ATOM) prepared from Agaricus blazei (Iwade Strain 101) “Himematsuke” and its mechanisms in tumor-beairng mice. Anticancer Res 17, 277-284, 1997.
2) Ebina T, Fujimiya Y. Agaricus blazei. Biotherapy 11, 259-265, 1998.
3) Mizuno TK. Bioactive biomolecules of mushrooms. food, func- tion and medicinal effect of mushroom fungi. Food Rev Int 11, 7-21, 1995.
4) Guterrez ZR, Mantovani MS, Eira AF, Ribeiro LR, Jordao BQ.
Variation of the antimutagenicity effects of water extracts of Agricus blazei Murill in vitro. Toxicol in Vitro 18, 301-309, 1994.
5) Kuo YC, Huang YL, Chen CC, Lin YS, Chuang KA, Tsai WJ. Cell cycle progression and cytokine gene expresssion of human peripheral blood mononuclear cells modulated by Agaricus blazei. Natl Res Insti Chin Med 155, 176-187, 2002.
6) Takeshi T, Yoshiyuki K, Hiromichi O. Isolation of an anti- tumor compound from Agaricus blazei Murill and its mecha- nism of action. Am Soc Nutr Sci 131, 1409-1413, 2001.
7) Bellini MF, Giacomini NL, Eira AF, Ribeiro LR, Mantovani MS. Anticlastogenic effect of aqueous extracts of Agaricus blazei on CHO-k1 cells, studying different developmental phases of the mushroom. Toxicol in Vitro 17, 465-469, 2003.
8) Martins de Oliveira J, Jordao BQ, Ribeiro LR, Ferreira da Eira A, Mantovani MS. Anti-genotoxic effect of aqueous extracts of sun mushroom (Agaricus blazei Murill lineage 99/26) in mammalian cells in vitro. Food Chem Toxicol 40, 1775-1780, 2002.
9) Lieberthal W, Koh JS, Levine JS. Necrosis and apoptosis in
acute renal failure. Semin Nephrol 18, 505-518, 1998.
10) Searle J, Kerr JF, Bishop CJ. Necrosis and apoptosis: distinct modes of cell death with fundamentally different significance.
Pathol Annu 17, 229-259, 1982.
11) Zimmermann KC, Bonzon C, Green DR. The machinery of programmed cell death. Pharmacol Ther 92, 57-70, 2001.
12) Chiarugi V, Magneli L, Cinelli M, Basi G. Apoptosis and the cell cycle. Cell Mol Biol Res 40, 603-612, 1994.
13) Evans VG. Mutiple pathways to apoptosis. Cell Biol Int 17, 461-476, 1993.
14) Nagata S. Apopotosis by death factor. Cell 88, 355-365, 1997.
15) Schultz DR, Harrington WJ Jr. Apoptosis: programmed cell death at a molecular level. Semin Arthritis Rheum 32, 345-369, 2003.
16) Chio WY, Park C, Lee JY, Kim GY, Park YM, Jeong YK, Lee YH, Choi YH. Anti-proliferative effects of water extract of Araricus blazei Murill in human cancer cell line A549. J Korean Soc Food Sci Nutr 33, 1237-1247, 2004.
17) Fabian D, Koppel J, Maddox-Hyttel P. Apoptotic processes during mammalian preimplantation development. Theriogeno- logy 64, 221-231, 2005.
18) Antonsson B, Martinou JC. The Bcl-2 protein family. Exp Cell Res 256, 50-57, 2000.
19) Jurgensmeier JM, Xie Z, Deveraux Q, Ellerby L, Bredesen D, Reed JC. Bax directly induces release of cytocrome c from isolated mitochondria. Proc Natl Acad Sci USA 95, 4997-5002, 1998.
20) Reed JC. Bcl-2 family proteins. Oncogene 17, 3225-3236, 1998.
21) Rosse T, Olivier R, Monney L, Rager M, Conus S, Fellay I, Jansen B, Borner C. Bcl-2 prolongs cell survival after Bax- induced release of cytochrome c. Nature 391, 496-499, 1998.
22) Lenaz G, Bovina C, Formiggini G, Castelli GP. Mitochondria, oxidative stress, and antioxidant defences. Acta Biochim Pol 46, 1-21, 1999.
23) Vegran F, Boidot R, Oudin C, Riedinger JM, Lizard-Nacol S.
Implication of alternative splice transcripts of caspase-3 and survivin in chemoresistance. Bull Cancer 92, 219-226, 2005.
24) Kaufmann SH, Desnoyers S, Ottaviano Y, Davidson NE, Poi- rier GG. Specific proteolytic cleavage of poly (ADP-ribose) polymerase: an early marker of chemotherapy-induced apop- tosis. Cancer Res 53, 3976-3985, 1993.
25) Lazebnik YA, Kaufmann SH, Desnoyers S, Poirier GG, Earn- shaw WC. Cleavage of poly (ADP-ribose) polymerase by a proteinase with properties like ICE. Nature 371, 346-347, 1994.
26) Tewari M, Quan LT, O’Rourke K, Desnoyers S, Zeng Z, Bei- dler DR, Poirier GG, Salvesen GS, Dixit VM. Yama/CPP32, a mammalian homolog of CED-3, is a CrmA-inhibitable pro- tease that cleaves the death substrate poly (ADP-ribose) poly- merase. Cell 81, 801-809, 1995.
27) Rowlands TM, Symonds JM, Farookhi R, Blaschuk OW.
Cadherins: crucial regulators of structure and function in reproductive tissues. Rev Reprod 5, 53-61, 2000.
28) Wijnhoven BP, Dinjens WN, Pignatelli M. E-cadherin-catenin cell-cell adhesion complex and human cancer. Br J Surg 87, 992-1005, 2000.
29) Fukuda K. Apoptosis-associated cleavage of β-catenin in human colon cancer and rat hepatoma cells. Int J Biochem Cell Biol 31, 519-529, 1999.
30) Steinhusen U, Badock V, Bauer A, Behrens J, Wittman- Liebold B, Dorken B, Bommert K. Apoptosis-induced cleav- age of β-catenin by caspase-3 results in proteolytic fragments
with reduced transactivation potential. J Biol Chem 275, 16345-16353, 2000.
31) Holcik M, Gibson H, Korneluk RG. XIAP: apoptotic brake and promising therapeutic target. Apoptosis 6, 253-261, 2001.
32) Hussein MR, Haemel AK, Wood GS. Apoptosis and melanoma: molecular mechanisms. J Pathol 199, 275-288, 2003.
33) Salvesen GS, Duckett CS. IAP proteins: blocking the road to death’s door. Nat Rev Mol Cell Biol 3, 401-410, 2002.