6
서 론
간세포암은 국내 성인남자 암사망률 2위를 차지하며 발 생원인으로는 B형-, C형 간염 바이러스의 감염, 대사성질 환에 의한 만성간질환 등이 알려져 있으나 분자생물학적 발생기전은 아직 완전히 밝혀지지 않은 상태이다.1 그 동안 의 많은 분자생물학적 연구들이 종양유전자(oncogene), 종 양억제유전자(tumor suppressor gene), 그리고 과오결합수정 유전자(mismatch repair gene) 등의 유전자 자체의 변화에 의 한 발암과정에 초점을 맞춰 왔으나,2 최근에는 유전자 서열 에는 영향을 주지 않으면서 특정 유전자의 발현을 억제하 고 세포분열 후에도 그대로 보존되는 epigenetic 기전에 관 심을 가지고 있다.3 특히 종양억제유전자와 과오결합수정 유전자의 promoter 영역의 DNA 메틸화가 해당 유전자의 발 현을 억제시키는 기전이 활발히 연구되고 있다. 간세포암 의 경우 B형-, C형 간염 바이러스의 감염이나 aflatoxin B1 과 같은 독소 등 환경적 요인이 발생원인의 대부분을 차지 하고 있어 이로 인한 DNA 메틸화와 간세포암의 발생과의 관계를 알아보는 것이 의미가 있으리라 생각한다.
p16 유전자는 9번 염색체의 단완 9p21 부위에 존재하며, 발현 단백질이 세포주기의 G1에서 cyclin D와 cdk4/6와의 결합을 억제함으로써 종양 발생을 억제하는 역할을 한다.4-6 p16 종양억제유전자는 Inhibitor of kinase [INK] 4a, Cyclin
Study of CpG Methylation of the p16 and MGMT
Promoter in Hepatocellular Carcinoma
Tae Gil Heo, M.D., Sang Hyun Shin, M.D., Yeo Goo Chang,
M.D., Seong Woo Hong, M.D., Kyung Mi Lee1, Jeong Hyun Kim, Ph.D.
1, Yun Kyung Kang, M.D.2, In Wook Paik, M.D. andHyuck Sang Lee, M.D.
Departments of Surgery, 1Institute of Molecular Biology and
2Pathology, Seoul Paik Hospital, Inje University College of Medicine, Seoul, Korea
Purpose: The aims of this study were to examine the
methylation status of the p16 and MGMT promoters in hepa- tocellular carcinoma (HCC), and to evaluate the relationship between the loss of gene expression, the promoter methyla- tion status and hepatocarcinogenesis.Methods: We included 24 HCC tissues and their adjacent
non-tumorous tissues and 5 normal liver tissues in our study, and all the specimens were obtained by hepatectomy. The methylation status of the p16 and MGMT promoter regions were evaluated by methylation-specific polymerase chain re- action (MSP) and quantitative analysis by using a Gel-pro analyzer (Media Cybernetics, CA, USA). We also analyzed the p16 and MGMT gene expressions by performing immuno- histochemical staining of the HCC tissues.Results: Methylation of the p16 promoter was detected in
HCC (100%, 24/24) and the adjacent non-tumorous tissues (79.2%, 19/24), but not in the normal liver tissues. Methy- lation of the MGMT promoter was detected in the HCC (8.3%, 2/24) and the adjacent non-tumorous tissues (4.2%, 1/24), but not in the normal liver tissues. Methylation positive HCC samples showed the loss of p16 expression in 58.3%(14/24). The loss of the p16 expression in the HCC tissues was well correlated with the increased rate of p16 promoter methylation (p=0.009). When the p16 promoter methylation status of the HCC tissues was higher than that of the adjacent non-tumorous tissues, 77.8% of the cases showed the loss of the p16 expression (p=0.002). No correlation was
책임저자:허태길, 서울시 중구 저동 2가 85번지 ꂕ 100-032, 서울백병원 외과 Tel: 02-2270-0016, Fax: 02-2270-0373 E-mail: [email protected]
본 논문은 2003년 인제대학교 연구비의 지원을 받았음.
간세포암에서 p16과 MGMT Promoter의 CpG 메틸화에 관한 연구
인제대학교 서울백병원 외과학교실, 1분자생물학연구소, 2병리학교실
허태길․신상현․장여구․홍성우․이경미1․김정현1․강윤경2․백인욱․이혁상
observed between MGMT promoter methylation and the loss of the gene expression in the HCC tissues.
Conclusion: These results suggest that methylation of the p16 promoter and the resulting loss of p16 protein expression
are significant events in hepatocarcinogenesis, and further studies are needed to evaluate the relationship between the methylation of the MGMT promoter and HCC carcinogenesis.(Korean J HBP Surg 2005;9:6-15)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Key Words:
Carcinoma, Hepatocellularp16
O
6-Methylguanine-DNA Methyltransferase
Methylation중심 단어: 간세포암, p16, O6
-methylguanine-DNA methylt-
ransferase (MGMT), 메틸화
dependent kinase inhibitor [CDK] 2A, Multiple tumor sup- pressor [MTS] 1 등으로도 알려졌는데, p16 유전자 메틸화 에 의한 불활성화는 가장 널리 알려진 epigenetic 기전 중 하나로 다양한 종류의 인체 종양 발생에 관련 있는 것으로 보고되었다.7
O6-methylguanine-DNA methyltransferase (MGMT)는 22,000 kd의 DNA 수정 효소로서 알킬화제(alkylating agents)의 발 암 및 세포독성 효과로부터 DNA를 방어하는데 중요한 역 할을 한다.8 N-nitros 복합체 같은 알킬화제는 간세포에서 주 로 대사되고 활성화되기 때문에 간조직은 항시 활성화된 알킬화제에 노출되어 있고,9 MGMT는 간에서 가장 높은 활 성도를 가지며 만성간염, 간경화, 간암에서 MGMT 단백질 활성도가 감소되어 있다고 보고되었다.10 또한 promoter 메 틸화에 의한 MGMT 유전자의 불활성화는 간세포암뿐만 아 니라 뇌종양, 대장암, 폐암 등 다른 암종들에서도 보고된 바
있다.11
이에 저자는 간세포암에서 p16 유전자와 MGMT 유전자 promoter 영역의 메틸화 유무와 정도를 알아보고 메틸화와 해당 유전자 전사억제와의 관계 및 간세포암 발생과의 관 계를 알아보고자 하였다.
방 법
1. 연구대상
서울백병원 외과학교실에서 간절제술로 얻은 24예의 간 세포암 조직과 그 인접부위 비종양 간조직을 대상으로 하 였다. 조직은 수술 직후 획득하여 냉동고(-80oC)에 보관해 오던 것들이었다. 대상 환자들의 평균 연령은 55.6세(범위:
43∼73세)였으며 남자는 22명, 여자는 2명이었다. B형 간염 양성 18예, C형 간염 양성 2예, B형 C형 간염 모두 양성 1
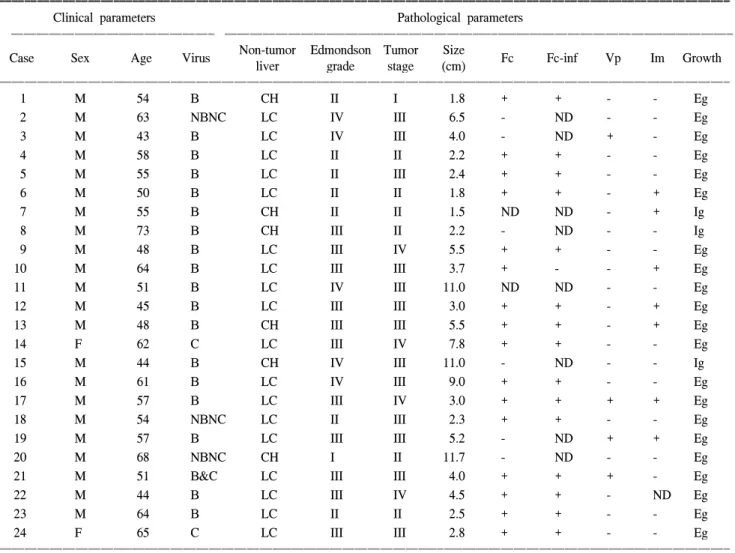
Table 1. Summary of the clinicopathologic parameters in hepatocellular carcinoma patients
ꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚ
Clinical parameters Pathological parameters
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Non-tumor Edmondson Tumor Size
Case Sex Age Virus Fc Fc-inf Vp Im Growth
liver grade stage (cm)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
1 M 54 B CH II I 1.8 + + - - Eg
2 M 63 NBNC LC IV III 6.5 - ND - - Eg
3 M 43 B LC IV III 4.0 - ND + - Eg
4 M 58 B LC II II 2.2 + + - - Eg
5 M 55 B LC II III 2.4 + + - - Eg
6 M 50 B LC II II 1.8 + + - + Eg
7 M 55 B CH II II 1.5 ND ND - + Ig
8 M 73 B CH III II 2.2 - ND - - Ig
9 M 48 B LC III IV 5.5 + + - - Eg
10 M 64 B LC III III 3.7 + - - + Eg
11 M 51 B LC IV III 11.0 ND ND - - Eg
12 M 45 B LC III III 3.0 + + - + Eg
13 M 48 B CH III III 5.5 + + - + Eg
14 F 62 C LC III IV 7.8 + + - - Eg
15 M 44 B CH IV III 11.0 - ND - - Ig
16 M 61 B LC IV III 9.0 + + - - Eg
17 M 57 B LC III IV 3.0 + + + + Eg
18 M 54 NBNC LC II III 2.3 + + - - Eg
19 M 57 B LC III III 5.2 - ND + + Eg
20 M 68 NBNC CH I II 11.7 - ND - - Eg
21 M 51 B&C LC III III 4.0 + + + - Eg
22 M 44 B LC III IV 4.5 + + - ND Eg
23 M 64 B LC II II 2.5 + + - - Eg
24 F 65 C LC III III 2.8 + + - - Eg
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ + = positive; - = negative; ND = not determined; size = tumor longest diameter; Virus B = HBsAg positive; C = HCV Ab positive;
NBNC = HBsAg and HCV Ab negative; LC = liver cirrhosis; CH = chronic hepatitis; Fc = capsule formation; Fc-inf = capsule infiltration;
Vp = portal vein thrombus; Im = intrahepatic metastasis; Ig = infiltrative growth; Eg = expanding growth.
예, B형 C형 간염 모두 음성 3예였다(Table 1). 정상 대조군 으로는 대장암 간전이로 간절제술을 시행한 후 획득한 5예 의 종양 주위 정상 간조직을 사용하였으며, 모두 B형 C형 간염 음성이었고, 지방간 및 간경화 소견은 없었다.
2. 방법
1) Genomic DNA의 추출: 30∼50 mg의 간조직을 500μl 의 PBS buffer (137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 1.76 mM KH2PO4)로 세척한 후, homogenizer를 이용하여 600μl의 Nuclei lysis solution (10 mM Tris-HCl (pH8.0), 400 mM NaCl, 2 mM EDTA (pH8.0))에 현탁하였다. 파쇄된 조직 에 Protease K (0.3 mg/ml), RNase A (0.05 mg/ml), SDS (0.5%
w/v)를 첨가하여, 65oC에서 3시간 동안 반응시키고, 600μl phenol/chloroform 처리하여 단백질을 제거하였다. Ethanol 침전을 수행하여 침전된 genomic DNA를 100∼200μl의 멸 균증류수에 용해시켰다.
2) Genomic DNA의 Bisulfite Modification: Genomic DNA
의 bisulfite modification은 CpGenomeTM DNA Modification Kit (Serologicals Corporation, CA, USA)를 이용하였다. Ge- nomic DNA 1μg에 reagent IV 2μl와 NaOH 7μl를 첨가하고, 50oC에서 10분간 반응시켰다. 이 용액에 reagent I 550μl를 넣고 50oC에서 16시간 반응시킨 후, reagent III 5μl와 re- agent II 750μl를 넣고 상온에서 10분간 반응시켰다. 6,500 rpm에서 원심분리하여 얻은 DNA를 ethanol (70% v/v)을 이 용하여 3회 세척하였다. 이 DNA에 20 mM NaOH/90% etha- nol 50μl를 넣고 상온에서 5분간 방치한 후, 다시 6,500 rpm 에서 원심분리하여 DNA를 침전시키고, Tris-EDTA (pH8.0) 20μl에 용해시켰다.3) 메틸화 특이성 중합효소연쇄반응(Methylation-specific polymerase chain reaction, MSP): 메틸화 특이성 중합효소연
쇄반응에 이용한 primer의 염기서열은 다음과 같다.12,13 p16-UF : p16 unmethylated specific forward primer 5'-TTATTAGAGGGTGGGGTGGATTGT-3' p16-UR : p16 unmethylated specific reverse primer 5'-CAACCCCAAACCACAACCATAA-3'p16-MF : p16 methylated specific forward primer 5'-TTATTAGAGGGTGGGGCGGATCGC-3'
p16-MR : p16 methylated specific reverse primer 5'-GACCCCGAACCGCGACCGTAA-3'
MGMT-UF : MGMT unmethylated specific forward primer 5'-TTTGTGTTTTGATGTTTGTAGGTTTTTGT-3' MGMT-UR : MGMT unmethylated specific reverse primer 5'-AACTCCACACTCTTCCAAAAACAAAACA-3' MGMT-MF : MGMT methylated specific reverse primer 5'-TTTCGACGTTCGTAGGTTTTCGC-3'
MGMT-MR : MGMT methylated specific reverse primer 5'-GCACTCTTCCGAAAACGAAACG-3'
PCR 조건은 다음과 같다. genomic DNA 100 ng, 각 primer 300 ng씩, Taq polymerase 1 unit를 사용하였으며, reaction volume은 25μl로 실시하였다. PCR reaction은 predena- turation 95oC에서 5분 1 cycle, denaturation 95oC에서 45초, 각 annealing 온도에서 45초, extension 72oC에서 45초로 35 cycle 그리고 final extension 72oC에서 3분 1 cycle 수행하였 다. Annealing 온도는 p16-W, M primer의 경우 65oC, p16-U primer는 60oC, MGMT-W, U, M primer는 61oC에서 각각 수 행하였다. 모든 PCR 반응에는 67 mM MgCl2, 12.5 mM dNTP를 사용하였고, 10 mM 2-Mercaptoethanol을 첨가하였다.
4) PCR product의 분석: Genomic DNA로부터 증폭된 각
PCR product들은 2% agarose 전기영동을 이용하여 전개하 였고, PCR 성공의 여부는 DNA의 크기와 증폭된 양을 기준 으로 결정하였다. PCR product의 정량은 Gel-pro analyzer (Media Cybernetics, CA, USA)를 이용하였으며, 100 ng 이상 은 ++, 100 ng 이하는 +로 표시하였다.5) 면역조직화학염색방법: 병리조직학적 검색을 통해 종
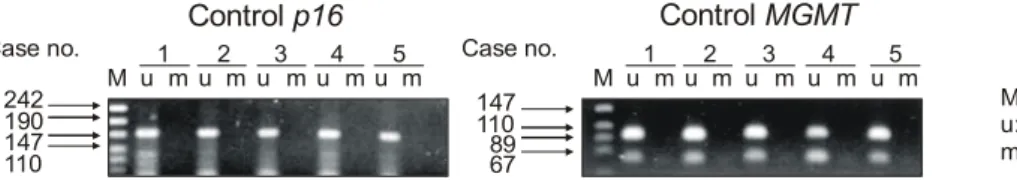
양이 충분히 포함된 블록을 선택하여 5μm의 연속 절편 2 장씩을 박절한 후 xylene으로 파라핀을 제거하고, 에틸알콜 에서 단계적으로 함수한 후 citrate buffer가 포함된 용기 내 에서 microwave로 5분간 가열하여 항원회복을 시도한 뒤, labeled streptavidin-biotin 방법으로 LSAB kit (DAKO, CA, USA)를 이용하여 면역조직화학염색을 시행하였다. 일차항 체로는 p16 단클론항체(희석비율 1:1,000, clone F-12, Santa Cruz Biotechnology, CA, USA)와 MGMT 단클론항체 (희석비율 1:100, clone MT3.1, Chemicon, CA, USA)를 사 용하였으며, 실온에서 1시간 반응시켰다. p16은 발현을 증Fig. 1. Methylation-specific PCR of p16 and MGMT promoter in normal liver tissues. Chromosomal DNA was modified as described in materials and methods. In all normal liver tissues assayed in this research, the discrete PCR products were amplified with unmethylated specific primers and no product with methylated specific primers, indicating no methylation of p16 and MGMT promoters in normal liver tissues. M = pUC18/Hpall DNA Marker; u = PCR with unmethylated specific primers; m = PCR with methylated specific primers.
1 1
242 147
190 110
147 89
110 67
M u mu2m u3mu4mu5m M u mu2m u3m u4m u5m
Case no. Case no.
M: pUC18/Hpall DNA Marker u: unmethylated primer m: methylated primer
Control p16 Control MGMT
폭시키기 위하여 catalyzed signal amplification system (K- 1500, DAKO, CA, USA)를 이용하여 증강효과를 얻었고, AEC (3-amino-9-ethylcarbazole) chromogen을 이용하여 발색
한 후 Mayer's hematoxylin으로 대조염색하였고, MGMT는 DAB (3,3'-diaminobenzidine) chromogen을 이용하여 발색한 후 Mayer's hematoxylin으로 대조염색하였다. 면역조직화학 염색의 판독은 p16, MGMT 모두 핵 내 발현이 전체종양세 포의 10% 미만에서 발현되는 경우 단백소실로 판정하였다.
6) 임상병리학적 인자와의 비교: 간세포암 조직에서 환자
의 나이, 성별, B형-, C형 간염여부, 종양의 크기, 잔존간의 상태, AFP 수치, TNM 병기(대한간암연구회, 원발성 간암 규약집, 2001년, 제1판), Edmondson grade, 피막형성 유무, 피막침범 유무, 문맥 색전형성 유무, 간내전이 유무, 성장양 식 등의 여러 임상병리학적 인자들과 p16 promoter의 메틸 화 정도를 비교하였다.3. 통계분석
통계학적 분석은 메틸화와 유전자 발현과의 관계, 메틸 화와 임상병리학적 인자와의 관계 모두 Pearson chi-square test와 Fisher's exact test를 이용하였으며 p 값이 0.05 미만을 유의수준으로 정의하였다.
결 과
1. p16 promoter methylation
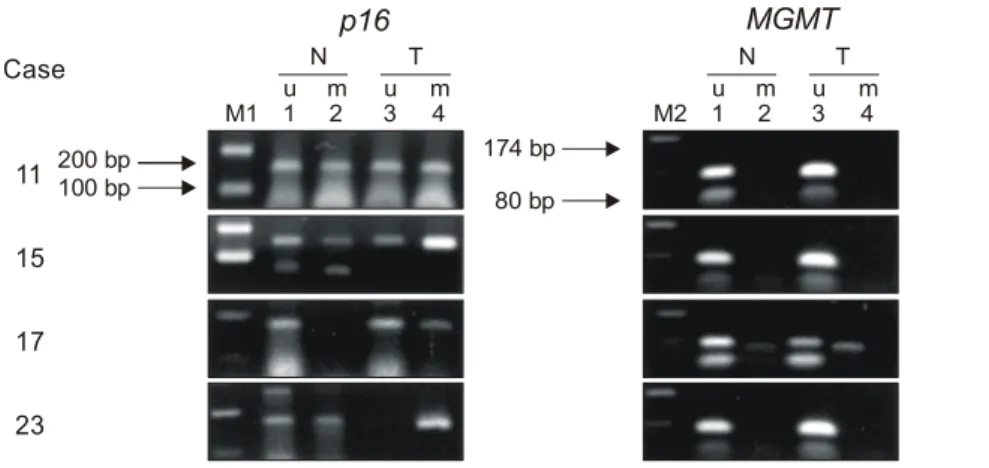
1) p16 promoter methylation의 빈도: 간세포암 조직에서
는 100% (24/24)에서 메틸화되었고, 인접 비종양 간조직에 서는 79.2% (19/24)에서 메틸화되었으며, 대조군으로 사용 한 정상 간조직에서는 메틸화된 예가 없었다(Fig. 1). 메틸 화 특이성 중합효소연쇄반응 후 정량분석 결과 메틸화 정 도는 간세포암 조직에서 2+가 70.8% (17/24), 1+가 29.2%(7/24), 음성 예는 없었으며, 인접 비종양 간조직에서는 2+
가 4.2% (1/24), 1+가 75.0% (18/24), 음성이 20.8% (5/24)로 나타나 인접 비종양 간조직보다 간세포암 조직에서 메틸화 정도가 높았다(p<0.001)(Table 2, 3)(Fig. 2).
2) p16 유전자 발현: p16 유전자 발현을 면역조직화학염색 으로 조사한 결과, 간세포암 조직에서는 58.3% (14/24)에서
Table 3. Methylation status of p16 and MGMT promoter in hepatocellular carcinoma tissues and adjacent non-tumorous tissues ꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚ
p16 MGMT
Status ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Tumor (%) Non-tumor (%) Tumor (%) Non-tumor (%)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
- 0 (0) 5 (20.8) p<0.001* 22 (91.6) 23 (95.8) p=0.599*
+ 7 (29.2) 18 (75.0) 1 (4.2) 1 (4.2)
++ 17 (70.8) 1 (4.2) 1 (4.2) 0 (0)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Total (%) 24 (100%) 24 (100%) 24 (100%) 24 (100%)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
*By Pearson chi-square test, Fisher's exact test. Methylation: + = below 100 ng; ++ = above 100 ng by quantitative analysis of PCR product using Gel-pro analyzer.
Table 2. Methylation status of p16 and MGMT promoter in hepatocellular carcinoma tissues and adjacent non-tumorous tissues ꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚ
p16 MGMT
Case ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Tumor Non-tumor Tumor Non-tumor
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
1 + + - -
2 + + - -
3 + - - -
4 ++ + - -
5 ++ + - -
6 ++ + - -
7 ++ ++ + -
8 ++ + - -
9 ++ + - -
10 ++ + - -
11 + + - -
12 ++ - - -
13 ++ + - -
14 ++ + - -
15 ++ + - -
16 + + - -
17 ++ - ++ +
18 ++ - - -
19 + + - -
20 ++ - - -
21 ++ + - -
22 + + - -
23 ++ + - -
24 ++ + - -
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ + = below 100 ng; ++ = above 100 ng by quantitative analysis of PCR product using Gel-pro analyzer.
소실되었고, 나머지 10예에서는 발현되었다(Table 4)(Fig. 3).
인접 비종양 간조직에서는 발현이 미약하여 단백소실여부 를 판정하지 못하였다.
3) p16 promoter methylation과 p16 유전자 발현과의 관
계: 간세포암 조직에서 p16 유전자 promoter의 메틸화 정도 가 2+인 경우 76.5% (13/17), 1+인 경우 14.3% (1/7)에서 p16 유전자 발현이 소실되어, 메틸화 정도가 높을수록 p16 유전자 발현 소실률이 높았다(p=0.009). 간세포암 조직 중 18예에서 인접 비종양 간조직보다 메틸화 정도가 증가하였 는데, 증가군의 p16 유전자 발현 소실은 18예 중 14예 (77.8%)에서 관찰되어, 증가하지 않은 6예에 비해 증가한 간세포암 조직에서 p16 유전자 발현 소실률이 높았다(p=0.002)(Table 5). Case 7의 경우 간세포암 조직과 인접 비종 양 간조직 모두 2+로 평가되었으나 간세포암 조직에서의 메틸화 정도가 현저하게 높아 메틸화가 증가한 군으로 간 주하였다(Table 2).
2. MGMT promoter methylation
1) MGMT promoter methylation의 빈도: 간세포암 조직
에서는 8.3% (2/24), 인접 비종양 간조직에서는 4.2% (1/24) 에서 관찰되었고, 대조군으로 사용한 정상 간조직에서는 메틸화된 예가 없었다(Fig. 1). 간세포암 조직에서 메틸화 정도는 2+가 1예, 1+가 1예, 인접 비종양 간조직에서는 1 +가 1예로 간세포암 조직과 메틸화 정도와는 유의한 상관 관계가 없었다(p=0.599)(Table 2, 3)(Fig. 2).2) MGMT 유전자 발현: MGMT 유전자 발현을 면역조직
화학염색으로 조사한 결과, 간세포암 조직에서는 33% (8/24)에서 소실되었고, 나머지 16예에서는 발현되었다(Table 4)(Fig. 4). 인접 비종양 간조직에서는 발현이 미약하여 단백 소실여부를 판정하지 못하였다.
Table 4. Methylation status and expression of p16 and MGMT in hepatocellular carcinoma tissues
ꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚ
p16 MGMT
Case ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Methylation Expression Methylation Expression ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
1 + + - +
2 + + - +
3 + - - -
4 ++ - - +
5 ++ - - +
6 ++ - - -
7 ++ - + -
8 ++ - - -
9 ++ - - -
10 ++ - - -
11 + + - +
12 ++ + - +
13 ++ - - +
14 ++ + - -
15 ++ + - +
16 + + - +
17 ++ + ++ +
18 ++ - - -
19 + + - +
20 ++ - - +
21 ++ - - +
22 + + - +
23 ++ - - +
24 ++ - - +
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Methylation: + = below 100 ng; ++ = above 100 ng by quan- titative analysis of PCR product using Gel-pro analyzer. Expres- sion: - = absent; + = present.
Fig. 2. Methylation-specific PCR of p16 and MGMT promoter in HCC tissues and adjacent non-tumorous tissues. The PCR products were quantified by Gel-Pro analyzer as described in materials and methods. M1 = 100 bp DNA ladder; M2 = pUC19/HaeIII; N = PCR with non-tumor; T = PCR with HCC; u = PCR with unmethylated specific primers; m = PCR with methylated specific primers.
11 15 17 23
N T N T
200 bp 174 bp
100 bp 80 bp
M1 1u m2 3u m4 M2 1u m2 3u m4 Case
M1: 100bp ladder marker M2: pUC19/Haelll marker N: non-tumorous tissue T: HCC tissue
u: unmethylated m: methlated
p16 MGMT
(3) MGMT promoter methylation과 MGMT 유전자 발현과
의 관계: MGMT 유전자 promoter 메틸화와 MGMT 유전자 발현과는 유의한 상관관계를 보이지 않았다(p=1.000)(Table 6).3. p16 promoter methylation과 임상병리학적 인자와의
비교환자의 나이, 성별, B형-, C형 간염여부, 종양의 크기, 잔존간의 상태, AFP 수치, TNM 병기(대한간암연구회, 원발 성 간암 규약집, 2001년, 제1판), Edmondson grade, 피막형성 유무, 피막침범 유무, 문맥 색전형성 유무, 간내전이 유무, 성장양식 등의 인자들과 p16 promoter의 메틸화 정도를 중 합효소연쇄반응 후 정량분석을 이용하여 비교하였다. Ed- mondson grade가 높아질수록 간세포암 조직에서 메틸화 정 도가 높아짐을 보였다(p=0.045). 그러나 다른 인자와는 유 의한 상관관계가 없었다. 간세포암 조직이 인접 비종양 간 조직보다 메틸화 정도가 증가한 군의 경우에도 이들 임상
양상 및 병리소견과 유의한 상관관계를 보이지 않았다 (Table 7).
Table 5. Relationship of p16 methylation status and p16 expression in hepatocellular carcinoma tissues
ꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚ
Loss of p16 Methylation (%) Methylation (%) Enhanced Enhanced
p-value* p-value*
expression + ++ methylation (%)(-) methylation (%)(+)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
(-) 6 (25.0) 4 (16.7) 0.009 6 (25.0) 4 (16.7) 0.002
(+) 1 (4.2) 13 (54.2) 0 (0.0) 14 (58.3)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
*By Pearson chi-square test, Fisher's exact test. Methylation + = below 100 ng; Methylation ++ = above 100 ng by quantitative analysis of PCR product using Gel-pro analyzer. Enhanced methylation = HCC methylation status is higher than its adjacent non-tumorous tissue;
- = absent; + = present.
Fig. 3. Immunohistochemical stains of p16 expression in hepato- cellular carcinoma. (a) p16 expression. The majority of tumor cells showed strongly stained nuclei (×200). (b) p16 expression loss.
The tumor cells showed loss of p16 expression compared to positive reaction of inflammatory cells (internal control)(×200).
a
b
Fig. 4. Immunohistochemical stains of MGMT expression in hepatocellular carcinoma. (a) MGMT expression. The majority of tumor cells showed strongly stained nuclei (×200). (b) MGMT expression loss. The tumor cells showed loss of MGMT expression compared to positive reaction of inflammatory cells (internal con- trol)(×200).
a
b
Table 6. Relationship of MGMT methylation status and MGMT expression in hepatocellular carcinoma tissues
ꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚ
MGMT MGMT
Loss of MGMT
methylation methylation p-value*
expression
(%) (-) (%) (+)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
(-) 15 (62.5) 1 (4.2) 1.000
(+) 7 (29.2) 1 (4.2)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
* By Pearson chi-square test, Fisher's exact test. - = absent; +
= present.
Table 7. Methylation status of p16 promoter and relationships to the clinicopathologic parameters in hepatocellular carcinoma tissues ꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚꠚ
Methylation (%) Methylation (%) Enhanced (%) Enhanced (%)
p-value* p-value*
+ ++ methylation (-) methylation (+)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Age (yr) 53.3±7.8 56.5±8.4 0.575 55.0±7.0 55.8±8.8 0.648
Sex 1.000 1.000
Male 7 (29.2) 15 (62.5) 6 (25.0) 16 (66.7)
Female 0 (0.0) 2 (8.3) 0 (0.0) 2 (8.3)
Size (cm) 6.0±3.1 4.3±3.1 0.472 6.3±3.3 4.3±3.0 0.348
HBsAg 1.000 1.000
(-) 1 (4.2) 4 (16.7) 1 (4.2) 4 (16.7)
(+) 6 (25.0) 13 (54.2) 5 (20.8) 14 (85.3)
HCV Ab 0.530 0.546
(-) 7 (29.2) 14 (58.3) 6 (25.0) 15 (62.5)
(+) 0 (0.0) 3 (12.5) 0 (0.0) 3 (12.5)
Ntm liver 0.629 1.000
Hepatitis 1 (4.2) 5 (20.8) 1 (4.2) 5 (20.8)
Cirrhosis 6 (25.0) 12 (50.0) 5 (20.8) 13 (54.2)
AFP 0.659 0.357
≤20 ng 3 (12.5) 10 (41.7) 2 (8.3) 11 (45.8)
>20 ng 4 (16.7) 7 (29.2) 4 (16.7) 7 (29.2)
Tumor stage 0.140 0.156
I 1 (4.2) 0 (0.0) 1 (4.2) 0 (0.0)
II 0 (0.0) 6 (25.0) 0 (0.0) 6 (25.0)
III 5 (20.8) 8 (33.3) 4 (16.7) 9 (37.5)
IV 1 (4.2) 3 (12.5) 1 (4.2) 3 (12.5)
Edmondson grade 0.045 0.231
I 0 (0.0) 1 (4.2) 0 (0.0) 1 (4.2)
II 1 (4.2) 6 (25.0) 1 (4.2) 6 (25.0)
III 2 (8.3) 9 (37.5) 2 (8.3) 9 (37.5)
IV 4 (16.7) 1 (4.2) 3 (12.5) 2 (8.3)
Fc 0.283 0.585
(-) 3 (13.6) 3 (13.6) 2 (9.1) 4 (18.2)
(+) 3 (13.6) 13 (59.1) 3 (13.6) 13 (59.1)
Fc-inf 1.000 1.000
(-) 0 (0.0) 1 (6.3) 0 (0.0) 1 (6.3)
(+) 3 (18.8) 12 (75.0) 3 (18.8) 13 (81.3)
Vp 0.584 0.964
Negative 2 (8.3) 5 (20.8) 2 (8.3) 5 (20.8)
Micro 3 (12.5) 10 (41.7) 3 (12.5) 10 (41.7)
Gross 2 (8.3) 2 (8.3) 1 (4.2) 3 (12.5)
Intrahepatic metastasis 0.621 1.000
(-) 5 (21.7) 11 (47.8) 4 (17.4) 12 (52.2)
(+) 1 (4.2) 6 (26.1) 1 (4.3) 6 (26.1)
Growth 0.530 0.546
Ig 0 (0.0) 3 (12.5) 0 (0.0) 3 (12.5)
Eg 7 (29.2) 14 (58.3) 6 (25.0) 15 (62.5)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
*By Pearson chi-square test, Fisher's exact test; Methylation + = below 100 ng; Methylation ++= above 100 ng by quantitative analysis of PCR product using Gel-pro analyzer; Enhanced methylation = HCC methylation status is higher than its adjacent non-tumorous tissue;
Ntm liver = non tumor liver.
고 찰
간세포암의 발생은 유전자 발현의 변화와 연관되어 있는 여러 단계의 과정에 의해 일어나며, 유전자 발현에 있어 전 암 단계의 변화는 DNA 메틸화, B형-, C형 간염 바이러스 의 작용, 특정세포 유전자에서의 점돌연변이, 또는 대립 유 전자 소실 등의 결과로 일어나고, 특정 종양억제유전자의 대립 유전자 소실과 종양유전자의 증폭이 암의 진행과정에 서 특징적으로 일어난다.14 또한 DNA 메틸화는 유전자를 불활성화 시키는 epigenetic 기전의 하나로 작용하여 직접 유전자 발현을 억제하기도 하고 이와 연관된 유전자에 돌 연변이가 일어나도록 하기도 한다.7,15
척추동물에서 생물학적으로 의미 있는 메틸화는 CpG di- nucleotide쌍의 cytosine 염기에서만 일어나는 것으로 알려져 있다. 메틸화된 DNA 서열에는 메틸결합단백(methylbinding protein, MBP)과 corepressor 분자 및 histone deacetylase (HDAC)로 구성된 단백질 복합체가 결합하게 되는데 메틸 결합단백은 메틸화된 CpG 결합 도메인과 다양한 corepres- sor 분자 및 histone deacetylase 단백과 복합체를 형성하는 전사억제도메인(transcriptional repressor domain)을 가진다.
메틸화된 DNA에 이 복합체가 결합하면 DNA를 둘러싸고 있는 histone이 deacetylation되고 핵질은 더욱 압축되어 전사 가 일어나지 못하게 된다. 또한 메틸화되지 않은 cytosine은 탈아민화되면 uracil로 변하고 이것은 uracil DNA glycosy- lase에 의해 제거되고 DNA 과오결합수정과정에 의해 cytosine-guanine 염기쌍으로 복구되지만, 메틸화된 cytosine (5-methylcytosine)은 저절로 탈아민화되어 thymine으로 변 하는데 thymine은 정상 DNA 염기이고 탈아민화되지 않고 thymine-guanine의 잘못된 염기쌍을 인지하여 제거하는 thymine DNA glycosylase는 효과가 없기 때문에 cytosine- guanine 염기쌍이 결과적으로 thymine-adenine 염기쌍으로 변하게 되는 결과를 초래하여 점돌연변이를 야기할 수 있 으며 이 과정이 종양억제유전자에 일어난다면 세포증식을 조절하는 능력을 상실하게 된다.15 인체의 많은 유전자의 promoter 영역에는 cytosine이 guanine과 인산염에 의해 결합 하고 있는 CpG dinucleotide가 연속해서 많이 나타나는데 이 부위를 CpG island라고 한다.16 Promoter 영역의 CpG island 의 메틸화는 종양세포에서 해당 유전자 전사의 불활성화에 관여하여 DNA 메틸화 연구의 중요한 대상이 되고 있다.
DNA 메틸화를 분석하는 방법으로는 Southern blot법, methylation-sensitive restriction enzyme PCR법, Combined bisulfite restriction analysis (COBRA)법, Bisulfite-PCR-SSCP (BiPS)법, PCR and fluorescence melting curve analysis법, Bisulfite sequencing법, Methylation-sensitive single-nucleotide primer extension (Ms-SNuPE)법, Methylation-specific PCR (MSP)법, Fluorescence-based real-time quantitative PCR anal-
ysis법 등이 있다.17
본 연구에서는 Methylation-specific PCR법을 사용하였는 데 메틸화된 cytosine을 관찰하기 위해 DNA를 먼저 bisulfite 로 화학반응시켜 메틸화되지 않은 모든 cytosine을 uracil로 변환되게 하고, 메틸화된 cytosine (5-methylcytosine)은 mo- dification되지 않고 그대로 남게 하여, 이를 PCR로 증폭하 여 cytosine의 메틸화를 확인하는 방법이다. PCR 증폭에는 unmodified DNA를 증폭하는 w-primer, CpG island의 cyto- sine에 메틸화된 DNA를 특이적으로 증폭하는 m-primer, CpG island의 cytosine에 메틸화가 되지 않은 DNA를 증폭하 는 u-primer 등 세 primer를 각각 사용하였다. 이 방법은 제 한효소를 사용하지 않아 불완전절단과 관련된 문제들을 피 할 수 있고, 민감도와 특이도가 높아 극소량의 sample로도 분석이 가능하며, 위양성의 결과가 나올 확률이 낮다는 장 점이 있다.17
세포주기의 진행은 촉진인자인 활성화된 cyclin 및 cyclin dependent kinase (CDK) 4/6과 억제인자인 cyclin dependent kinase inhibitors (CDKIs)에 의해 조절된다. Cyclin D1-CDK 4/6 복합체에 의해 retinoblastoma (Rb) 단백질이 인산화(pho- sphorylation)되면 세포주기 중 G1기가 S기로 전환하는데 CDKIs는 cyclin-CDK 복합체에 결합하여 그 기능을 억제함 으로써 세포주기를 조절하는 G1 조절점(checkpoint) 역할을 한다.4-6 CDK 억제제로 작용하는 단백질에는 p16INK4a, p15INK4b, p18INK4c, p19INK4d, P21CIP1, p27KIP1, p57- KIP2 등이 알려져 있다.4,18 이중 p16 종양억제유전자는 Inhi- bitor of kinase [INK] 4a, Cyclin dependent kinase inhibitor [CDKN] 2A, Multiple tumor suppressor [MTS] 1 등으로도 알 려졌는데, p16 유전자의 메틸화에 의한 불활성화가 가장 널 리 알려진 epigenetic 기전 중 하나로 식도암, 대장 및 직장 암, 간암, 유방암, 방광암, 췌장암, 폐암, 난소암, 흑색종, 가 스트린종, 두경부암, 림프종 등 여러 종류의 종양 발생에서 관련이 있는 것으로 보고되었다.7
O6-methylguanine-DNA methyltransferase (MGMT)은 22,000 kd의 DNA 복구단백질로, 메틸화된 guanine을 빠르게 복원 시키는 데 중요한 역할을 하는데,11,19 O6-guanine 위치는 DNA 알킬화에 의한 강력한 mutagenic region으로 그 원인은 주로 DNA 복제 시 O6-methylguanine이 thymine과 짝을 이루 기 때문에 결과적으로 guanine-cytosine 염기쌍이 adenine- thymine 염기쌍으로 변환되기 때문이다.20 MGMT의 활성도 는 종마다, 그리고 장기마다 크게 차이가 나는데, 인간에서 는 간에서 가장 높은 활성도를 가지며,9 만성간염, 간경화, 간세포암에서 MGMT 단백질의 활성도가 감소한다.10 Promoter 메틸화에 의한 MGMT 유전자의 불활성화는 뇌 종양, 대장암, 폐암 등 다른 암종들에서 알려졌는데,11 이러 한 발현 소실은 MGMT 유전자의 결손이나 변이, 재배열에 의한 것은 드물고 MGMT 유전자에서 불연속적으로 존재하 는 CpG island의 메틸화와 연관되어 있으며,21-23 비정상적인
MGMT의 메틸화는 mRNA 발현의 소실,24 MGMT 단백질의 부족,11 효소 활성도의 소실로 이어진다.25
본 연구에서는 간세포암으로 간절제술을 시행한 후 얻은 24예의 간세포암 조직과 인접 비종양 간조직에서 대장암 간전이로 간절제술 시행 후 획득한 종양 주위 정상 간조직 5예를 대조군으로 사용하여, p16과 MGMT 유전자 promoter 의 메틸화 빈도와 정도를 메틸화 특이성 중합효소연쇄반응 과 정량분석을 이용하여 알아보고 면역화학조직염색을 통 하여 간세포암 조직에서의 발현을 조사하였다. 비종양 간 조직에서는 p16과 MGMT의 발현이 전반적으로 미약하여 단백소실여부를 판정하지 못하였다.
p16 유전자의 경우 간세포암 조직에서는 100% (24/24)에 서 메틸화되었고, 인접 비종양 간조직에서는 79.2% (19/24) 에서 메틸화되었으며, 정상 대조군에서는 한 예도 메틸화 되지 않아(0/5), 정상 간조직은 물론 인접 비종양 간조직보 다 간세포암 조직에서 p16 유전자 promoter 영역의 메틸화 빈도가 높았다. 메틸화 정도 또한 간세포암 조직에서는 2+
가 70.8% (17/24), 1+가 29.2% (7/24), 음성이 0% (0/24), 인 접 비종양 간조직에서는 2+가 4.2% (1/24), 1+가 75.0%
(18/24), 음성이 20.8% (5/24)로 나타나 인접 비종양 간조직 보다 간세포암 조직에서 메틸화 정도가 높았다. 이는 p16 유전자 promoter 메틸화가 간세포암 발생에 관여함을 시사 한다. 정상 간조직은 물론 인접 비종양 간조직보다 간세포 암 조직에서 p16 promoter 메틸화가 빈도뿐만 아니라 정도 에서도 높게 나타났는데 이는 간세포암 발생과정에서 새 로운 메틸화가 계속해서 일어남을 시사하는 소견이라 생 각된다. 간세포암 조직에서 100%의 메틸화를 보였고, 인접 비종양 간조직에서는 만성간염의 경우 6예 중 5예(83.3%), 간경화의 경우 18예 중 14예(77.8%)의 메틸화를 보여 각각 72.7%, 23.5%, 29.4%의 메틸화를 보인 Kaneto 등26의 연구보 다 높게 나타났다.
p16 유전자 발현은 메틸화 양성을 보인 총 24예의 간세포 암 조직 중 58.3% (14/24)에서 소실되었고, 나머지 10예에서 발현하였는데, 메틸화 양성 간세포암 조직에서 100% 발현 소실을 보인 Kaneto 등26의 연구와 차이가 있었다. 메틸화 정도가 2+인 17예 중 13예(76.5%)에서, 1+인 7예 중 1예 (14.3%)에서 발현 소실을 보여 메틸화 정도가 높을수록 발 현 소실률이 높았다. 또한 인접 비종양 간조직보다 메틸화 정도가 증가한 군에서 증가하지 않은 군보다 p16 유전자 발 현 소실률이 높았다. 메틸화 양성이었으나 발현한 10예의 경우 메틸화 정도가 2+가 4예, 1+가 6예였는데, p16 유전 자 발현 소실은 p16 유전자 promoter CpG island의 메틸화된 cytosine 염기의 축적으로 나타나는 축적효과(dose depen- dent manner)에 의한 것이거나, 특정부위의 메틸화가 p16 유 전자 전사억제에 중요하게 작용하는 것으로 발생하리라 생 각되어, 향후 promoter 영역의 메틸화와 유전자 전사억제 기 전에 관한 연구가 필요하리라 생각된다.
간세포암 조직에서 여러 임상병리학적 인자들과 p16 promoter의 메틸화 정도를 비교하였는데 Edmondson grade 가 높을수록 메틸화 정도가 높아짐을 보였으나, B형-, C형 간염의 종류 및 간염의 유무 등 다른 인자들과는 유의한 상관관계가 없었다. 특히 인접 비종양 간조직은 만성간염 6예 중 5예, 간경화 18예 중 14예에서 메틸화 양성을 보여 잔존 간 상태인 만성간염, 간경화 여부와 메틸화 정도와는 유의한 상관관계를 보이지 않았다.
MGMT 유전자 promoter 메틸화는 간세포암 조직에서는 8.3% (2/24)에서, 인접 비종양 조직에서는 4.2% (1/24)에서 관찰되었으나 대조군으로 사용한 정상 간조직에서는 한 예 도 메틸화되지 않았다(0/5). 간세포암 조직에서 MGMT 유전 자 발현은 33% (8/24)에서 소실되었는데 promoter의 메틸화 정도와 유전자 발현 소실과는 유의한 상관관계를 보이지 않았다. Matsukura 등27은 46예의 간세포암 조직 중 28예 (60.9%)에서 메틸화를 보인다고 발표하였는데 promoter 영 역 중 주로 앞쪽(up stream)의 CpG island (No. 1∼9)에서 메 틸화를 보여 유전자 전사부위와 가까운 뒤쪽(down stream) 을 조사한 본 연구와 차이가 있었다.
p16 유전자 promoter 영역의 메틸화는 정상 간조직에서는 관찰되지 않았고, 인접 비종양 간조직보다 간세포암 조직 에서 더 많은 빈도로 관찰되었고, 간세포암에서 메틸화 정 도가 높을수록 높은 p16 유전자 소실률을 보였으며, 인접 비종양 간조직보다 간세포암 조직에서 메틸화가 증가될수 록 간세포암 조직의 p16 유전자 소실률이 높았다. 이와 같 은 소견을 감안하였을 때 p16 유전자의 promoter 영역의 메 틸화에 의한 p16 유전자 발현 소실이 간세포암 발생과정에 있어 중요한 기전중의 하나임을 시사한다. MGMT 유전자의 경우 promoter 영역의 메틸화와 MGMT 유전자 소실, 간세포 암 발생과의 관계는 추후 발전된 분자생물학적 실현기법을 이용한 추가연구가 필요하리라 생각한다.
참 고 문 헌
1) Hwang IS, Hong SW, Nah YW, Jang YG, Kim KH, Lee HS.
Analysis of 38 long-term survivors after liver resections for hepatocellular carcinomas. Korean J HBP Surg 2000;4:67-76.
2) Tahara E. Molecular mechanism of stomach carcinogenesis. J Cancer Res Clin Oncol 1993;119:265-272.
3) Baylin SB, Herman JG, Graff JR, Vertino PM, Issa JP. Al- terations in DNA methylation: a fundamental aspect of neo- plasia. Adv Cancer Res 1998;72:141-196.
4) Wijnhoven BP, Tilanus HW, Dinjens WN. Molecular biology of Barrett's adenocarcinoma. Ann Surg 2001;233:322-337.
5) Serrano M, Hannon GJ, Beach D. A new regulatory motif in cell-cycle control causing specific inhibition of cyclin D/
CDK4. Nature 1993;366:704-707.
6) Sherr CJ, Roberts JM. Inhibitors of mammalian G1 cyclin-
dependent kinase. Genes Dev 1995;9:1149-1163.
7) Herman JG, Merlo A, Mao L, et al. Inactivation of the CDKN2/p16/MTS1 gene is frequently associated with aberrant DNA methylation in all common human cancers. Cancer Res 1995;55:4525-4530.
8) Pegg AE. Mammalian O6-alkylguanine-DNA alkyltransferase:
regulation and importance in response to alkylating carcino- genic and therapeutic agents. Cancer Res 1990;50:6119-6129.
9) Gerson SL, Trey JE, Miller K, Berger NA. Comparison of O6-alkylguanine-DNA alkyltransferase activity based on cellu- lar DNA content in human, rat and mouse tissue. Carcinoge- nesis 1986;7:745-749.
10) Major GN, Collier JD. Repair of DNA lesion O6-methylgua- nine in hepatocellular carcinogenesis. J Hepatobiliary Pancreat Surg 1998;5:355-366.
11) Esteller M, Hamilton SR, Burger PC, Baylin SB, Herman JG.
Inactivation of the DNA repair gene O6-methylguanine-DNA methyltransferase by promoter hypermethylation is a common event in primary human neoplasia. Cancer Res 1999;59:793- 797.
12) Herman JG, Graff JR, Myohanen S, Nelkin BD, Baylin SB.
Methylation-specific PCR: a novel PCR assay for methylation status of CpG islands. Proc Natl Acad Sci USA 1996;93:
9821-9826.
13) Zhang YJ, Chen Y, Ahsan H, et al. Inactivation of the DNA repair gene O6-methylguanine-DNA methyltransferase by pro- moter hypermethylation and its relationship to aflatoxin B1- DNA adducts and p53 mutation in hepatocellular carcinoma.
Int J Cancer 2003;104:440-444.
14) Feitelson MA, Sun B, Satiroglu Tufan NL, Liu J, Pan J, Lian Z. Genetic mechanisms of hepatocarcinogenesis. Oncogene 2002;21:2593-2604.
15) Wajed SA, Laird PW, DeMeester TR. DNA methylation: an alternative pathway to cancer. Ann Surg 2001;234:10-20.
16) Bird AP. CpG-rich islands and the function of DNA methy- lation. Nature 1986;321:209-213.
17) Liu ZJ, Maekawa M. Polymerase chain reaction-based methods of DNA methylation analysis. Anal Biochem 2003;317:
259-265.
18) Myung N, Kim MR, Chung IP, Kim HS, Jang JJ. Loss of p16 and p27 is associated with progression of human gastric cancer. Cancer Lett 2000;153:129-136.
19) Pegg AE, Byers TL. Repair of DNA containing O6-alkylgua- nine. FASEB J 1992;6:2302-2310.
20) Coulondre C, Miller JH. Genetic studies of the lac repressor.
III. Additional correlation of mutational sites with specific amino acid residues. J Mol Biol 1977;117:525-567.
21) Costello JF, Futscher BW, Tano K, Graunke DM, Pieper RO.
Graded methylation in the promoter and body of the O6- methylguanine DNA methyltransferase (MGMT) gene corre- lates with MGMT expression in human glioma cells. J Biol Chem 1994;269:17228-17237.
22) Qian XC, Brent TP. Methylation hot spots in the 5' flanking region denote silencing of the O6-methylguanine-DNA methyl- transferase gene. Cancer Res 1997;57:3672-3677.
23) Watts GS, Pieper RO, Costello JF, Peng YM, Dalton WS, Futscher BW. Methylation of discrete regions of the O6- methylguanine DNA methyltransferase (MGMT) CpG island is associated with heterochromatinization of the MGMT transcription start site and silencing of the gene. Mol Cell Biol 1997;17:5612-5619.
24) Esteller M, Toyota M, Sanchez-Cespedes M, et al. Inactivation of the DNA repair gene O6-methylguanine-DNA methyltrans- ferase by promoter hypermethylation is associated with G to A mutations in K-ras in colorectal tumorigenesis. Cancer Res 2000;60:2368-2371.
25) Herfarth KK, Brent TP, Danam RP, et al. A specific CpG methylation pattern of the MGMT promoter region associated with reduced MGMT expression in primary colorectal cancers.
Mol Carcinog 1999;24:90-98.
26) Kaneto H, Sasaki S, Yamamoto H, et al. Detection of hyper- methylation of the p16 (INK4A) gene promoter in chronic hepatitis and cirrhosis associated with hepatitis B or C virus.
Gut 2001;48:372-377.
27) Matsukura S, Soejima H, Nakagawachi T, et al. CpG methy- lation of MGMT and hMLH1 promoter in hepatocellular carcinoma associated with hepatitis viral infection. Br J Cancer 2003;88:521-529.