금앵자 에탄올 추출물에 의한 3T3-L1 지방세포의 분화억제 효과와 그 메커니즘 규명
정현영1, 정인교2, 남소연2, 윤희정2, 김병우1,2, 권현주1,2*
1동의대학교블루바이오소재개발및실용화지원센터
2동의대학교생명응용학과
Received: November 23, 2015 / Revised: January 14, 2016 / Accepted: January 14, 2016
서 론
비만이란섭취한에너지와소비하는에너지사이의심각 한불균형으로인하여신체내지방이축적되는현상으로, 고지혈증, 간기능이상, 당뇨병, 고혈압, 심혈관계질환등의 성인병이나각종암발병의직·간접적원인이되고있어비 만의치료및예방이시급한실정이며[2, 16, 24, 31, 32], 이
를위한연구가활발히진행되고있다[35]. 체내지방은지
방전구세포가지방세포로분화한후지방세포의증식과정에
서지방이축적되는현상이다. 지방전구세포가과다분화하 거나분화된지방세포가과다증식하면지방세포의수가많 아지고, 지방세포내지방축적량이증가할수록지방세포의 크기가커져비만으로이어질수있다[7, 25]. 따라서, 지방 세포의증식및분화억제, 지방세포내지방축적억제및축 적된지방의분해를통해비만을억제할수있다.
지방전구세포가지방세포로분화하는과정은세포의형태 나호르몬감수성, 다양한유전자들상호작용등이동반되는 복합한과정이다. 이러한반응은여러가지전사인자들의단 계적 작용에 의해 일어나는데, 그 중 CCAAT/enhancer- binding protein(C/EBP) family와 peroxisome proliferator activated receptor γ가가장대표적인전사인자로알려져있 다[6,36]. 지방전구세포인 3T3-L1은 confluence 상태즉, 세 Inhibitory Effects and Molecular Mechanism of Adipocyte Differentiation by Rosae laevigata Fructus Ethanol Extracs
Hyun Young Jeong1, In Kyo Jeong2, So Yeon Nam2, Hee Jung Yun1,2, Byung Woo Kim1,2, and Hyun Ju Kwon1,2*
1Blue-Bio Industry Regional Innovation Center, 2Department of Life Science and Biotechnology, College of Natural Science, Dong-Eui Uni- versity, Busan 47340, Republic of Korea
Obesity is caused by excess accumulation of body fat and contributes to various pathological disorders such as diabetes, hypertension, cardiovascular disease, and cancer. In this study, we investigated the effect of a 30% ethanol extract of Fructus Rosae laevigata (RLE) on adipogenesis in 3T3-L1 adipocytes, measured by triglyceride accumulation and expression of adipogenesis-related transcription factors during differen- tiation of pre-adipocytes into adipocytes. RLE decreased the intracellular triglyceride contents (assessed by Oil Red-O staining) in a dose-dependent manner. It also downregulated the expression of adipogenic transcription factors and inhibited cell proliferation during the mitotic clonal expansion phase of adipo- cyte differentiation by inducing G1 phase arrest. We investigated the alterations in the levels of G1 phase arrest–related proteins. The expression of p21 protein significantly increased, while the levels of Cyclin E, Cdk2, and phospho-Rb decreased in a dose-dependent manner in 3T3-L1 cells treated with RLE. These results suggest that RLE inhibits the differentiation of 3T3-L1 adipocytes by suppressing the expression of adipogenic transcription factors and inducing G1 phase arrest in the early stages of adipocyte differentia- tion.
Keywords: Rosae laevigatae fructus 30% EtOH extract, inhibition of adipocyte differentiation, inhibition of clonal expansion, inhibition of adipogenic transcription factor
*Corresponding author
Tel: +82-51-890-1519, Fax: +82-505-182-6871 E-mail: [email protected]
© 2016, The Korean Society for Microbiology and Biotechnology
포배양용 dish에세포들이가득차면 contact inhibition으 로인하여세포주기가정지되어세포의성장이멈추게된
다. 이 상태에서 지방세포 유도 복합체인 MDI(insulin,
dexamethasone, 3-isobutyl-1-methylxanthine)를처리하면 정지된세포주기가다시진행되어세포의수가 2−3배늘어 나게 되는데, 이 과정을 clonal expansion 단계라 부르고,
3T3-L1 지방전구세포가지방세포로분화될때매우중요한
과정이며, 이과정이진행되지않으면분화가일어나지않는 다고보고되어있다[3, 8, 34]. 지방세포의 clonal expansion
단계에서핵심적인역할을하는인자는바로 C/EBPβ이다.
지방세포유도복합체처리에의해 C/EBPβ, C/EBPδ의발 현이유도되고세포주기의진행이시작된다. 또한 C/EBPβ 는 C/EBPα와 PPARγ의 promoter에결합하여발현을유도 한다[4, 33, 38]. 지방세포의분화를조절하는핵심인자인 C/
EBPα와 PPARγ가발현이되면지방세포유도복합체에의
해진행되던세포주기가다시정지되며, C/EBPα와 PPARγ 가서로상호작용을하면서다양한지방세포의분화및지 방합성과저장에관여하는유전자들의발현을유도하여세 포내지방이축적되고분화가완료된다[1, 10, 20, 21, 26].
금앵자(Rosae laevigata fructus)는열대지방에서자라는 상록성관목의장미과식물이며그열매와뿌리를약용으로 이용한다. 금앵자는가시가있고마치적갈색의작은석류와 흡사한모양을하고있으며열매속에 30−40개의황갈색종 자가들어있다. 금앵자의성미는평(平)하고, 시고떫은맛 이난다[23]. 성분으로는꽃받침과열매에유기산, 탄닌, 정 유, 비타민 C, 수지, 사포닌이있다. 한방에서는금앵자의꽃 받침과열매를강장, 수렴, 지사, 유정, 유뇨, 빈뇨, 만성장염,
설사등에사용되어왔다[13]. 금앵자의항산화효능, 혈액성
상에관한연구[14, 17], 항염증효능[9]에관한연구가보고 되어있으나, 항비만효능에관한연구는아직보고되어있 지않다.
본연구에서는금앵자가항비만효능을가지고있는지확
인하기위해 3T3-L1 지방전구세포를인위적으로분화시킬
때금앵자추출물을처리하여나타나는변화를확인하였고, 지방세포분화에관련된인자들인 C/EBPα, C/EBPβ, PPARγ 등의발현에미치는영향을조사하였다. 또한분화초기지 방세포유도복합체에의해세포가증식하는과정에서금앵 자가미치는영향을확인하였다.
재료 및 방법
금앵자 추출
금앵자(Rosae laevigatae fructus) 100 g을분말화한다음, 30% EtOH 1 L를넣고 75oC에서 3시간동안 3회반복하여추 출하였다. 추출액은 ADVANTEC filter paper 2(ADVANTEC, Bunkyo-ku, Tokyo, Japan)로여과하여불순물을제거하고 감압농축하여추출물 29.75 g을얻었다. 이후금앵자 30%
EtOH 추출물을 RLE(Rosae laevigatae fructus 30% EtOH extracts)로표기하고자한다.
3T3-L1 지방전구세포의 배양
3T3-L1(mouse embryonic fibroblast cell line) 지방전구 세포는 ATCC(American type culture collection)에서구입 하여 Dulbecco's modified Eagle's medium(DMEM, WelGene Biopharmaceuticals, Daegu, Korea)에 10% bovine calf serum(BCS, WelGene Biopharmaceuticals, Daegu, Korea), antibiotics(penicillin/streptomycin 100 units/ml, Bioshop, Burlington, Ontario, Canada)를 첨가하고 37oC, 5% CO2
세포배양기에서배양하였다. 지방세포로의 분화
지방전구세포인 3T3-L1을지방세포로분화시키기위하여
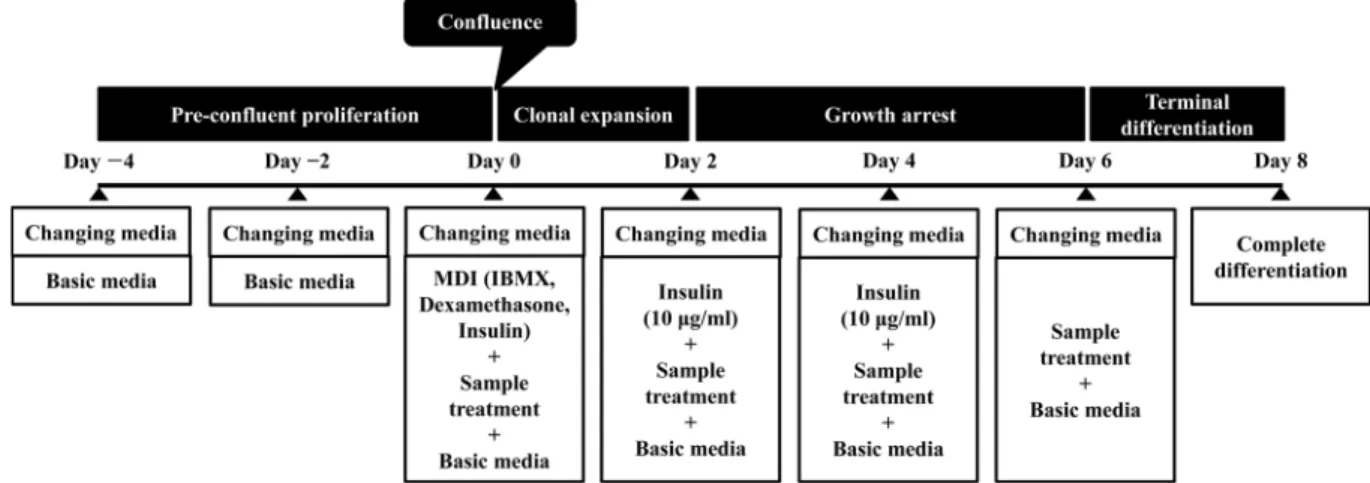
Fig. 1. Diagram of adipocyte differentiation process.
35-mm dish(SPL Life Sciences, Seoul, Korea)에 5×104 cells/dish의농도로세포를 seeding한후, 10% fetal bovine serum(FBS), penicillin/streptomycin solution(100 units/
ml)이첨가된 DMEM 배지로 confluence 상태가될때까지 배양하였다(Fig. 1, Day 0). 그후, 지방세포분화유도제인 MDI[10 μg/ml insulin(Sigma-Aldrich, St Louis, MO, USA), 0.25 μM dexamethasone(Sigma-Aldrich, St Louis, MO, USA), 0.5 mM 3-isobutyl-1-methylxanthine(IBMX, Sigma- Aldrich, St Louis, MO, USA)]를첨가한 DMEM 배지를세 포에처리하여 2일간배양하였다(Day 2). 이후, 지방세포로 의분화를촉진하기위해 10μg/ml insulin이포함된 DMEM 으로배지를교환하여 4일간배양하였고(Day 6), 완전분화 를위해 DMEM에 10% FBS, antibiotics를첨가하여 2일간 더배양하였다(Day 8). RLE가지방세포분화에미치는영 향은 MDI나 insulin 처리시, RLE를함께처리하여확인하 였다.
Oil Red O 염색
RLE가지방세포 분화에 미치는 영향을 확인하기위해
MDI와 insulin 처리및완전분화를위한배지에 RLE를함 께처리하였다. 0.5% DMSO를처리한대조군이 100% 분화 가되었을때, 배지를제거하고차가운 phosphate buffered saline(PBS, Gibco, Grand Island, NY, USA)으로 cell을 washing 한 후, 10% formalin(Junsei Chemical, Tokyo, Japan)을처리하여실온에서 1시간고정시켰다. Formalin 을 완전히 제거한 다음 60% isopropanol(Burdick and Jackson, Muskegon, MI, USA)로 헹군 뒤 Oil Red O working solution 을넣고실온에서 10분간염색을하였다. 이 때 Oil Red O working solution은 0.35 g Oil Red O powder(Sigma-Aldrich, St Louis, MO, USA)를 100 ml isopropanol에녹인뒤증류수에 6 : 4의비율로희석한다음 여과하여사용하였다. 염색후 Oil Red O working solution
을완전히제거하고증류수로 4번 washing한뒤현미경으
로세포의염색상태를확인하였으며, 흡광도측정을위하여 100% isopropanol에용출시켜 spectrophotometer를이용하 여 500 nm에서흡광도를측정하였다.
세포 독성
RLE의 독성을 알아보기 위하여 PreMix WST-1 Cell
Proliferation Assay System(Takara, Tokyo, Japan)을 이 용하였다. 3T3-L1 지방전구세포를 96-well plate(5×103 cells/well)에 seeding 하고 24시간배양하였다. 이후, 대조 군으로 0.5% DMSO(Sigma-Aldrich, St Louis, MO, USA) 를처리하였으며, RLE를 600−1000 μg/ml의농도로처리하 고 48시간 배양하였다. WST-1 reagent를각 well에 20 μl
씩 첨가하여 30분 반응시킨 다음 ELISA reader(Molecular Devices, Sunnyvale, CA, USA)로 450 nm에서흡광도를측 정하였다.
분화초기와 분화후기에 발현되는 전사인자들의 발현양상 확인
RLE에의한지방전구세포에서지방세포로의분화과정 동안발현되는전사인자들의발현변화를확인하기위해, 지 방전구세포에 MDI와함께 RLE를처리하고 2일후세포를 회수하여 C/EBPβ 발현양상을확인하였다(Day 2). 또한지
방세포분화후기에발현되는 C/EBPα와 PPARγ의발현양
상을확인하기위해 MDI와 RLE를함께처리하여 2일간반 응시킨다음, FBS가첨가된 DMEM 배지에 insulin만처리 할때 RLE를함께처리하여 4일간반응시킨다음세포를회 수하였다(Day 6).
Western blot analysis
RLE를처리한 3T3-L1 세포를회수하여 PBS로 washing 한 후, lysis buffer(Cell signaling technology, Danvers, MA, USA)를세포에처리하고 4oC에서 1시간반응시켜용 해시킨후 14,000 rpm, 4oC에서 20분간원심분리하여상등 액을얻었다. 상등액의단백질농도는 BCA법을이용하여정 량한후동량의단백질을 (SDS)-polyacrylamide gel에서전 기영동하였다. 전기영동후 gel 내의단백질을 polyvinylidene fluoride(PVDF) membrane(Bio-Rad, Hercules, CA, USA) 에전사시키고 Blocking solution[0.15 M NaCl, 1 M Tris- HCl(pH 7.5), 0.1% Tween-20, 5% BSA]을사용하여 4oC에 서 16시간 blocking 시켰다. 1차항체는 4oC에서 16시간반 응시켰으며, TBS[50 mM Tris-HCl(pH 7.5), 0.15 M NaCl]
에 0.1% Tween-20이첨가된 buffer를사용하여 membrane 을 washing 한다음 2차항체를넣고 4oC에서 16시간반응 시켰다. 면역반응단백질은화학발광시스템(Chemi-luminescence system; Super Signal West Femto Maximum sensitivity Substrate, Pierce, USA)으로검출하였다. 본실험에사용한 1차항체는 Cell signaling technology(Danvers, MA, USA) 에서 C/EBPα, C/EBPβ, Rb, phospho-Rb를구입하였으며, Santa Cruz Biotechnology Inc.(Santa Cruz, CA, USA)에 서 p21, Cyclin E, Cdk2, Actin를구입하여사용하였다. 2차 항체인 anti-rabbit IgG-HRP, anti-mouse IgG-HRP는 Santa Cruz Biotechnology Inc.(Santa Cruz, CA, USA)에서구입 하여사용하였다.
Clonal expansion 저해
3T3-L1 지방전구세포를 100-mm dish(SPL Life Sciences, Seoul, Korea)에 0.5×106 cells/dish의농도로 seeding 하여
confluent 상태가 될 때까지 2일간 더 배양하였다. 그 후
MDI가포함된 DMEM으로배지를교환할때 RLE를처리
하였다. RLE 처리후각각 0, 24, 48시간에세포를회수한 후 0.4% Trypan blue solution(Sigma-Aldrich, St Louis,
MO, USA)과 세포 현탁액을 1:1 비율로 염색한 다음
hemacytometer를이용하여생세포수를측정하였다.
세포 주기 확인
3T3-L1 지방전구세포를 60-mm dish에 1×105 cells/dish (SPL Life Sciences, Seoul, Korea)의농도로 seeding 하여 배양하였다. 세포를 confluence 상태까지배양한후 MDI를 포함하는배지로교환할때 RLE를처리하였다. 24시간후 에세포를회수하여 PBS로세척후 70% EtOH로고정시키 고, MuseTM Cell Cycle Reagent(Millipore Co. Milford, MA, USA)를 처리하여 30분간 염색한 후 MuseTM Cell
Analyzer(Millipore Co. Milford, MA, USA)로분석하였다.
통계분석
실험결과는평균(mean)± 표준편차로(SD)로나타내었으며, Statistical Package for the Social Sciences(SPSS 18.0;
SPSS Inc., Chicago, IL, USA) 통계프로그램을 사용하여 분석하였다. 각군간의유의성검정은 t-test를수행하여검 증하였다.
결과 및 고찰
RLE의 지방세포 내 triglyceride 축적억제 효과 확인
RLE가지방세포분화에영향을미치는지알아보기위하
여 3T3-L1 지방전구세포가 confluent 상태가된이후전체 분화과정동안 100−1000 μg/ml 농도의 RLE를함께처리하
Fig. 2. Inhibitory effects of Rosae laevigatae fructus 30% EtOH extracts (RLE) on the triglyceride accumulation in fully dif- ferentiated 3T3-L1 adipocytes. 3T3-L1 pre-adipocytes were differentiated by adding adipogenic inducers (insulin, dexamethasone and 3-isobutyl-1-methylxanthine) when the cells reached confluence, followed by 2 days of incubation. Complete differentiation into adipocytes was achieved by further culturing the cells in 10% FBS/DMEM with 10 µg/ml insulin for 4 days and then in 10% FBS/
antibiotics/DMEM for 2 days. RLE was treated during all differentiation process. (A) Intracellular triglycride was stained with Oil Red O.
(B) Oil Red O was dissolved with isopropanol and optical density detected at 500 nm. Values are represented as the mean ± SD (n = 3) *p < 0.05, **p < 0.01 compared by control.
여지방세포내 triglyceride 축적정도를확인하였다. 그결 과 RLE를처리하지않고 0.5% DMSO를처리한대조군의분
화정도를 100%로하였을때, RLE를농도별로처리한실험
군들의 triglyceride의축적율이 RLE의농도가증가할수록 감소하였다(Fig. 2). 그러나, RLE의농도 100−500 μg/ml에 서는효과가미미하였고, 600 μg/ml부터 triglyceride 축적억 제효과가뚜렷하게나타나, 최고농도인 1000 μg/ml에서는 대조군에비해약 84%의 triglyceride 축적억제효능을나타 내었다. 따라서 RLE는지방전구세포에서지방세로로의분 화를억제하는효능이높은것으로사료되며, 이후실험은 triglyceride 축적억제효능이뚜렷하게나타나는 600−1000 μg/ml의농도의 RLE를사용하였다.
RLE의 3T3-L1 세포에서의 독성확인
앞서확인한 RLE에의한 triglyceride 축적억제효능이세
포독성에의한효능인지확인하기위하여 PreMix WST-1
Cell Proliferation Assay System을사용하여세포생존율을 확인하였다. Triglyceride 축적억제효과가있는 600−1000 μg/ml의 RLE 농도로세포생존율을확인한결과, RLE 농 도 600, 700, 800, 900 그리고 1000 μg/ml에서각각 99.5± 4.74, 95.6±4.64, 90.35±6.63, 94.06±2.97, 85.59±4.03%로 측정되었다(Fig. 3). 따라서 RLE에의한 triglyceride 축적억
제효과는 3T3-L1에대한독성효과는아닌것으로사료된다.
RLE에 의한 지방세포 분화 관련 전사인자의 발현 변화 지방세포의 분화는 peroxisome proliferation activated receptor gamma(PPARγ), CCAAT enhancer binding protein
(C/EBP) family라고불리는전사인자들이중추적인역할을
담당하며다양한상호작용을통해조절된다[18]. 지방세포 유도복합체인 MDI(insulin, dexamethasone, 3-isobutyl-1-
methylxanthine)는이러한전사인자의발현을촉진함으로
써지방전구세포를지방세포로분화시킨다. 바꾸어말하면 지방세포분화관련전사인자들의발현저해를통하여지방 세포로의분화를억제하고지방축적감소등을야기시킬수 있으므로, 이는항비만물질탐색의기작연구에가장많이
쓰이는방법중하나이다. 따라서, RLE에의한지방축적억
제효과가지방세포의분화를조절하는전사인자들의발현 변화와관련되는지알아보았다.
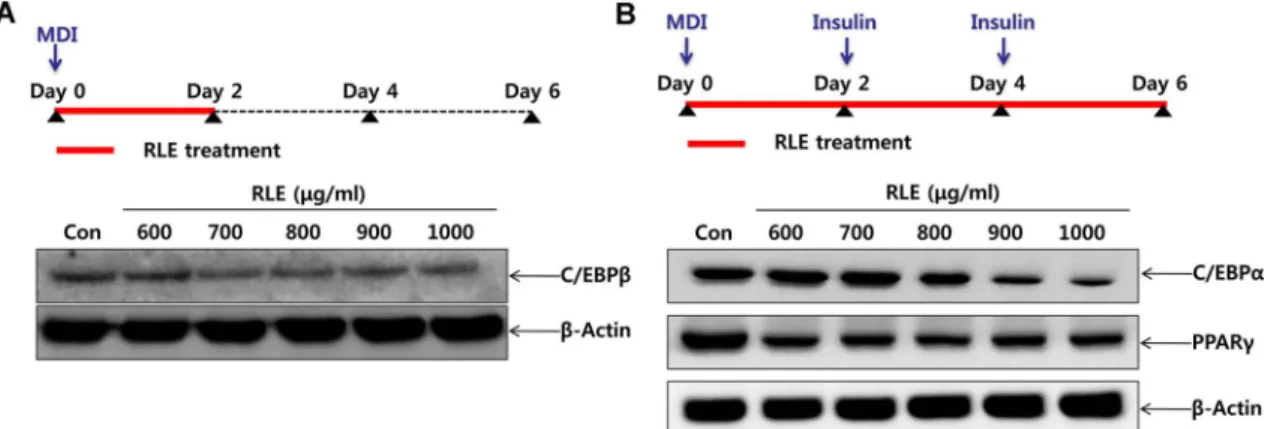
C/EBPβ는지방세포의분화초기에 MDI에의해발현되는
단백질로지방전구세포에 MDI와 RLE를 2일간복합처리한
후(Day 2) 세포를회수하여단백질발현양상을확인한결
Fig. 4. Effects of Rosae laevigatae fructus 30% EtOH extracts (RLE) on expression of adipogenic transcription factors in early stages (A) and late stages (B) of adipocyte differentiation of 3T3-L1. (A) 3T3-L1 pre-adipocytes were incubated in DMEM containing adipogenic incucers (insulin, dexamethasone and 3-isobutyl-1-methylxanthine) for 2 days with various concentrations (600−1000 µg/
ml) of RLE. (B) 3T3-L1 pre-adipocytes were cultured in DMEM containing adipogenic inducers (insulin, dexamethasone and 3-isobutyl- 1-methylxanthine) with RLE for 2 days, and then in DMEM containing insulin for 4 days with various concentrations (600−1000 µg/ml) of RLE. Cells were harvested and were lysed and then equal volumes of proteins determined by western blot analysis.
Fig. 3. Cytotoxic effects of Rosae laevigatae fructus 30%
EtOH extracts (RLE) on 3T3-L1 pre-adipocyte. 3T3-L1 pre-adi- pocytes were treated with various concentrations (600−1000 µg/
ml) of RLE for 2 days. And then, cell viability was evaluated by PreMix WST-1 Cell Proliferation Assay System. Values are repre- sented as the mean ± SD (n = 3) *p < 0.05 compared by control.
과 RLE를처리한실험군의 C/EBPβ의발현량이대조군보 다감소되는것을확인할수있었다(Fig. 4A). 또한 PPARγ
와 C/EBPα는 C/EBPβ의자극을 받아발현하는단백질들
로, adipogenesis에서 중요한 transcription factor로 널리 알려져있고, 지방세포완전분화의지표로사용된다. RLE에 의한 PPARγ와 C/EBPα의발현량의변화를확인하기위해, MDI와 RLE를복합처리하여 2일간반응시키고, insulin과
RLE를 4일간복합처리한후단백질발현양상을확인한결
과(Day 6), RLE의농도의존적으로 PPARγ와 C/EBPα의발 현이감소하였다(Fig. 4B). 따라서, RLE는지방세포의분화
초기에 C/EBPβ의발현을저해시킬뿐아니라, 이후분화전
체단계에서중요한역할을하는 PPARγ및 C/EBPα의발현 또한저해시켜지방세포의분화를억제시키는것으로사료
된다. 즉, RLE는지방세포의분화를유도하는핵심전사인
자들의발현을저해하여지방세포의분화를억제하는것으 로사료된다.
RLE에 의한 분화 초기 세포 증식 억제(Clonal expansion 억제) 지방세포로의분화초기단계에서 contact inhibition에의 해증식이정지되어있던세포는지방세포유도복합체에의 해다시세포증식을시작하는데, 이과정에서핵심적인역 할을 하는 인자가 바로 C/EBPβ이다[34]. Fig. 4A에서 C/
EBPβ의발현이 RLE에의해저해됨을확인하였기에, RLE
가 지방세포 유도 복합체에 의한 세포 증식, 즉 clonal
expansion 단계를저해시킬것이라판단하고, 이를확인하
기위해 trypan blue exclusion assay를통해생세포수를확
인하였다. 그결과대조군의세포증식은 0시간일때 100%
라고봤을때 24, 48시간에는 154.77, 225.1%의증식을보였 지만 RLE 900 μg/ml 처리군은 119.6, 158.03%, RLE 1000
μg/ml 처리군은 101.59, 127.85%로증식이억제됨을확인하 였다(Fig. 5). 이러한증식억제효과는 RLE를처리하기전 confluent 상태, 즉 Day 0에서의 세포수와비교하였을 때 RLE 처리이후생세포수의감소는유도되지않는것으로보 아세포독성에의한효과는아닌것으로사료된다. 따라서, RLE는 3T3-L1 세포의분화초기 clonal expansion 단계에서 세포증식억제를통해지방세포로의분화를억제하는것으 로사료된다.
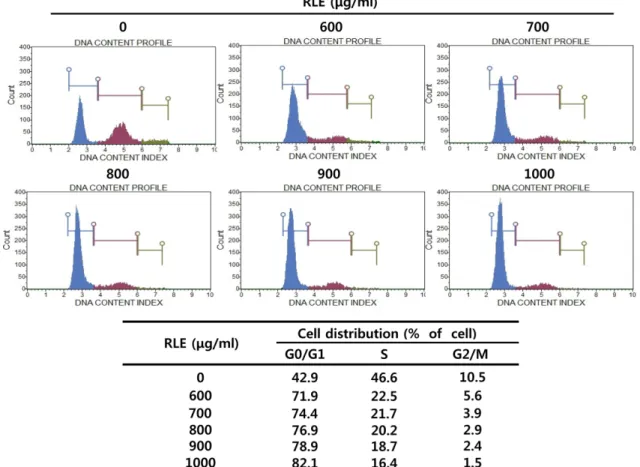
RLE에 의한 3T3-L1 지방전구세포의 G1기에서의 세포주기 정 지 효과
위의결과에의하면, RLE는지방전구세포가지방세포로
분화하는초기단계에서세포분열을억제시킨다. 즉, 정지 된세포주기가 MDI로인해다시진행될때, RLE의작용에
의해세포주기진행의변화가있을것으로생각되어 MuseTM
Cell Analyzer를이용하여세포주기분포도를확인하였다. Fig. 6에서와 같이 RLE를 처리하지 않고 MDI와 0.5%
DMSO를처리한대조군의 G1기의세포비율은 42.9%로나 타났으나, RLE를처리한군들의 G1기세포비율은 RLE 처 리농도가높아질수록점점증가하여최고농도인 1000 μg/
ml에서는 82.1%로나타났다. 이러한결과는, RLE가 clonal
expansion 단계에있는지방세포의세포주기를 G1기에서정
지시킴으로써, 지방세포로의분화를억제시킨다는것을나 타낸다.
G1 arrest 관련 단백질의 발현변화
RLE가분화초기지방세포유도복합체인 MDI 처리에의
한세포의증식유도단계에서, G1기에서세포주기를정지 시킴으로써세포의증식을억제함을확인하였다. RLE에의 한 G1기로의세포주기정지에대한분자기전을확인하기위 해 세포주기 조절에 관여하는 단백질들의 발현 변화를 western blot analysis를 통해 확인하였다. 세포주기는 Cyclin에의한 cyclin-dependent kinase(Cdk)의활성화및 Cdk inhibitor의발현에의해조절된다[5]. 그중 G1기에서 S기로진입하는단계에서는 Cyclin E가발현되어 Cdk2를활 성화시켜세포주기가진행된다. 그러나어떠한요인에의해 Cyclin과 Cdk 발현이정상적으로유도되지않으면, 필요한 단백질의합성이충분히진행될때까지세포주기가정지된 다고알려져있다. 또한 Cyclin E와 Cdk2가활성화되면 Rb 를인산화시키게되는데, Rb가인산화되면 DNA 복제를진 행시키는전사인자가발현되어 S기가시작된다[27, 30]. Cdk
는다양한증식억제신호들에의해발현되는 Cdk inhibitor
인 p21에의해그활성이억제된다고알려져있다[19]. 3T3- L1 지방전구세포에 MDI와다양한농도의 RLE를동시에처 리하여배양한후, 회수하여 G1기에서 S기로의전이에관여 Fig. 5. Inhibitory effects of Rosae laevigatae fructus 30%
EtOH extracts (RLE) on cell proliferation in clonal expansion phase of adipocyte differentiation period. 3T3-L1 pre-adipocytes were treated in DMEM containing adipogenic inducers (insulin, dexamethasone and 3-isobutyl-1-methylxanthine) with indi- cated concentration (600−1000 µg/ml) of RLE. Viable cells were calculated by trypan blue exclusion assay in 0, 24, 48 h.
하는단백질들인 Cdk2, Cyclin E, Rb, phosphor-Rb 및 p21 의발현변화를검토하였다. 그결과 RLE의농도의존적으 로 Cdk inhibitor인 p21의발현이증가하였고, Cyclin E와 Cdk2, phosphor-Rb의발현은감소됨을확인하였다(Fig. 7).
이는 RLE에의해 증가된 Cdk inhibitor인 p21이 CyclinE/
Cdk2 complex에결합하여 CyclinE/Cdk2 complex의활성이 저해되고따라서, 이후단계의 Rb의인산화가억제되어 S기 로의진행이저해되고 G1 기에서세포주기정지가유도되었 을것으로사료된다.
본연구에사용된금앵자는장미과의금앵자나무열매를 건조한것으로, 유기산, 탄닌, 정유, 비타민 C, 수지, 사포닌 등의성분이함유되어있는것으로알려져있다. 금앵자가 속해있는장미과식물중비파엽, 용아초, 복분자, 산사등에
의한지방세포분화억제효능에대한보고가있다[12, 22, 28,
37]. 그중비파엽은 3T3-L1 cell에서 PPARγ와 C/EBPα의 발현억제를통해지방세포의분화를억제시키는것으로, 금 앵자추출물의효능및작용메커니즘이유사한것으로사 Fig. 6. Induction of G1 cell cycle arrest on clonal expansion phase of adipocyte differentiation process in Rosae laevigatae fruc- tus 30% EtOH extracts (RLE) treated 3T3-L1 adipocyte. 3T3-L1 pre-adipocytes were incubated in DMEM containing adipogenic inducers (insulin, dexamethasone and 3-isobutyl-1-methylxanthine) with various concentrations (600−1000 µg/ml) of RLE for 24 h. Cells were harvested and then fixed in 70% EtOH. Cells were stained with Muse™ Cell Cycle Reagent and analyzed by flow cytometer.
Fig. 7. Alteration of G1 phase-related proteins expression in 3T3-L1 adipocyte treated with Rosae laevigatae fructus 30%
EtOH extracts (RLE). 3T3-L1 pre-adipocytes were cultured in DMEM containing adipogenic inducers (insulin, dexamethasone and 3-isobutyl-1-methylxanthine) for 2 days with various con- centrations of (600−1000 µg/ml) RLE. Cell were harvested and lysed and then equal volumes of proteins were determined by western blot analysis.