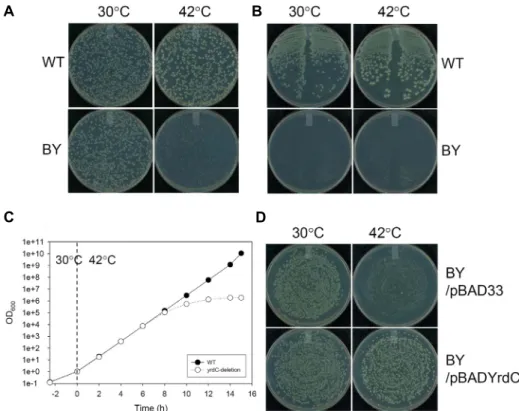
Isolation of Temperature-sensitive Mutant Escherichia coli YrdC Involved in Universal t<sup>6</sup> A tRNA Synthesis
8
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
(7)
(8)
수치



+2

관련 문서
Thus, boiling occurs at a given pressure acting on the fluid by raising the temperature, or at a given fluid temperature by lowering the pressure.. Cavitation phenomena in
1 John Owen, Justification by Faith Alone, in The Works of John Owen, ed. John Bolt, trans. Scott Clark, "Do This and Live: Christ's Active Obedience as the
Relationship of temperature at the mature stage, Atmospheric pressure distribution, airflow, convective cells.. The vertical distribution analyzed by K-LAPS at
First, there was no significant difference as a result of analyzing the differences in psychological capital, achievement goal orientation, and game roots
Comparison of the monthly average difference between surface temperature and sea surface temperature at Chilbaldo on foggy(f_avg) and no foggy(nf_avg) day cases from
The results show that the green seed extracts are only shown as antioxidants, but the others are shown as a prooxidants against DNA damage and Escherichia coli
The used output data are minimum DNBR values in a reactor core in a lot of operating conditions and the input data are reactor power, core inlet
Snow accumulation in January and the mean snowfall at each station in Jeollanam-do region from 1991 to 2008 · ·
