대한 두경부 종양 학회지 제 30 권 제 2 호 2014
The Effect of Cancer Associated Fibroblasts(CAFs) and CD44 of CAFs on the Motility of Cancer Cells in Head
and Neck Squamous Cell Carcinoma
*Seon-Hui Shim, PhD1,2, J. Hun Hah, MD, PhD2,3, Soo Youn Cho, MD, PhD4, Tae-min Kim, MD, PhD5, Young-il Koh, MD5, Dong-Wan Kim, MD, PhD2,5, Choon-Taek Lee, MD, PhD6, Dae Seog Heo, MD, PhD1,2,5, Myung-Whun Sung, MD, PhD1,2,3 Department of Molecular Tumor Biology1 and Cancer Research Institute,2 Seoul National University College of Medicine,
Seoul, Korea
Department of Otorhinolaryngology-Head and Neck Surgery,3 Seoul National University College of Medicine, Seoul, Korea Department of Pathology,4 Korea Cancer Center Hospital, Seoul, Korea
Department of Internal Medicine,5 Seoul National University Hospital, Seoul, Korea
Department of Medicine and Respiratory Center,6 Seoul National University Bundang Hospital, Seongnam, Korea
두경부편평세포암에서 암-연관 섬유아세포가 암세포의 운동성에 미치는 영향과 암-연관 섬유아세포에서 과발현되는 CD44의 역할*
서울대학교 의과대학 분자종양생물학교실,1 암연구소,2 이비인후과학교실,3 원자력병원 병리과,4
서울대학교병원 내과,5 서울대학교분당병원 내과 및 폐센터6
심선희1,2·하정훈2,3·조수연4·김태민5·고영일5·김동완2,5·이춘택6·허대석1,2,5·성명훈1,2,3
= 국 문 초 록 =
배경 및 목적
암-연관 섬유아세포(Cancer-associated fibroblasts, CAF)는 종양미세환경의 가장 중요한 요소의 하나다. 그래서 두경부편평세포암에 대해 CAF가 암세포의 운동성에 미치는 영향을 평가하고, CAF에 과발현되는 CD44의 역할에 대해 평가하고자 하였다.
재료 및 방법
두경부암환자의 종양조직에서 CAF를 분리하고, 비종양성 조직으로부터 정상 섬유아세포(NHF)를 분리하였다.
창상치유분석과 상하 챔버를 이용한 3차원 세포 이동 분석을 이용하여, CAF가 암세포의 이동에 미치는 영향을 분석 하고, CAF에서 과발현되는 CD44를 중화항체로 CAF를 차단했을 때 암세포 이동의 변화를 관찰하였다.
결 과
NHF에 비해 CAF에서 CD44가 과발현되는 것을 관찰하였다. 창상치유분석에서 CAF와 같이 배양된 암세포는 NHF 와 같이 배양된 암세포에 비해 더 빠른 이동을 보였다. CD44 중화 항체를 처리했을 때는 암세포의 이동성이 저해되었다.
결 론
CAF는 종양미세환경에서 암세포의 운동성을 조절하는 중요한 인자의 하나일 것으로 사료된다. CD44는 CAF의 기능을 매개하는 중요한 표지자 중 하나로 생각된다.
중심 단어:두경부 편평세포암종·섬유아세포·CD44·암-연관 섬유아세포.
Received : September 29, 2014 / Accepted : October 2, 2014
*This work was supported by grant No. 0420090730 from the SNUH Research Fund.
교신저자 : 하정훈, 110-744 서울 종로구 대학로 101 서울대학교 의과대학 이비인후과학교실 전화 : (02) 2072-0215 ・ 전송 : (02) 745-2387 ・ E-mail : [email protected]
online©MLComm
Introduction
Cancer-associated fibroblasts(CAFs) came to be considered one of the major players that make cancer-favorable micro-en- vironments.1) CAFs are characterized as activated fibroblasts and are also known as myofibroblasts that express α-smooth muscle actin(α-SMA).2) Carcinomas bearing myofibroblasts correlate with a poorer prognosis.3,4) If fibroblasts acquire an activated phenotype, proliferative activity increases and many extracellular matrix(ECM) proteins, such as type I collagen, are secreted.5) The fibroblasts in invasive breast cancer were reported to promote tumor growth and angiogenesis through elevated SDF-1 secretion.6) In addition, significant numbers of activated fibroblasts were found at the site of wound healing, which implicates these cells as the main mediator of organ fibrosis.7) Fibroblasts in cancer tissue that have an activated phenotype are shown to have different characteristics from those found in normal tissues.5)
CD44 is a cellular adhesion receptor that is upregulated following tissue injury and is also implicated in many chron- ic inflammatory diseases8) As a major receptor for hyaluronan (HA), CD44 participates in the activation of leukocytes and parenchymal cells in areas of inflammation.9) The possible interaction between CD44 and cytoskeletal components may suggest a role for CD44 in tissue remodeling and fibrosis through modulation of the adhesion and motility of fibro- blasts.10,11)
Oncologically, CD44 is a multi-structural and multifunction- al cell-surface molecule modulating cell proliferation, cell dif- ferentiation, cell migration, and angiogenesis. Reports sug- gested that CD44 is involved in tumor progression.12,13) In head and neck squamous cell carcinoma, stronger expression of CD44 was found in metastatic lymph nodes, compared to that of primary tumors.14) Importantly, CD44 expression in surgical margins negatively influenced survival rate in laryn- geal cancer patients.14)
Furthermore, CD44 is a possible surface marker of cancer stem cells for head and neck cancers.15) Many studies have suggested markers by separating from a single tumor sample two distinct subpopulations of cancer cells that differ from each other in their cell-surface proteins and their ability to seed new tumors in vivo.16,17)
In a study with scar fibroblasts, the differences in CD44 ex- pression levels were observed between scar fibroblasts and normal fibroblasts.18) Scar fibroblasts expressed higher levels of CD44 than normal skin fibroblasts did. Scar fibroblasts and CAFs might share similar traits because both are categorized as activated fibroblasts. Thus, we hypothesized that CAFs may
overexpress CD44 and CAFs highly expressing CD44 may have an important role in the tumor microenvironment. In this study, we report the possibility that CD44 plays a signif- icant role in the interaction between cancer cells and CAFs in the tumor microenvironment.
Materials and Methods
1. Cell lines and cell culture
The human HNSCC cell lines SNU-1041, SNU-1066 and SNU-1076 were purchased from the Korea Cell Line Bank (Seoul, Korea). Cells were cultured in RPMI 1640 medium with 10% fetal bovine serum and gentamycin(Gibco BRL, Grand Island, NY). We extracted fibroblasts from tissues ob- tained from head and neck cancer patients. Tissues were di- gested with collagenase and placed into culture dishes in DMEM containing, gentamycin and Fungizone supplement- ed with 10% FBS. After 10-12 days, the fibroblasts were sep- arated by differential trypsinization with the fibroblast remain- ing through passage(Supplementary 1). CAFs from tumor tissues were cultured less than fifteen times. Normal human fibroblasts(NHFs) were obtained from muscle tissue at the resection margins, adequately apart from the tumors. NHF was not thought to be contaminated with tumor cells because all the resection margins were negative for the tumors by the pathology reports. NHF is cultured in the same method as were the CAFs. Collection of the tissue samples was approved by Institutional Review Board(IRB number : H-1001-063-307).
To verify the purity of fibroblasts in collected samples, West- ern-blot analysis was performed for fibroblast markers, such as vimentin(BioVision, CA) and α-SMA(DAKO, Denmark).
2. Wound healing assay
Cancer cells were seeded in the bottom chamber of a tran- swell dish(SPL, Seoul, Korea), and a straight scratch was made with a pipette tip the next day. CAFs or NHFs were then placed in the upper chambers of the transwell dish. After one or two days, the migration of cells from the wound edges was captured with an OLYMPUS IX51(Tokyo, Japan).
3. Blocking antibody treatment
CAFs or NHFs were incubated with 1 µg of normal mouse IgG(Santa Cruz Biotechnology, Santa Cruz, CA) or CD44 blocking antibody(Neomarker, Fremont, CA) in PBS for 30 min and washed two times before seeding.
4. Migration assay and colony formation assay The space between the upper and lower chambers in the tran- swell dish was filled with 0.5% soft agar in DMEM contain-
ing 10% FBS, and cells were seeded in the upper chambers of 24-transwell or 6-transwell dishes(SPL, Seoul, Korea). Two weeks after seeding, cells in the bottom chambers were stained with crystal violet(Sigma-Aldrich, St. Louis, MO).
5. Gene expression measurement by Western blot and FACS analysis
Cultured cells were rinsed with PBS, suspended in lysis buffer(0.5% NP40, 50 mM Tris-Cl, 150 mM NaCl, 1 mM di- thiothreitol, 1% sodium deoxycholate, 0.1% SDS, 1 mM EDTA, 1 mM phenylmethylsulfonyl fluoride, 0.1 M aprotinin and 1 M pepstatin A) and incubated at 4℃ for 30 minutes. Cell ly- sates were then centrifuged at 13,000 r.p.m. for 20 minutes at 4℃. An appropriate amount of each supernatant(determined by protein assay) was mixed with 4X sample loading buffer and denatured for 10 minutes at 70℃. The denatured protein samples were fractionated on 4-12% NuPAGETM Bis-Tris gels(Invitrogen, Carlsbad, CA), transferred onto nitrocellulose membranes(Schleicher & Schuell, Dachen, Germany), and incubated with Tris-buffered saline containing 0.1% Tween- 20(TBS-T) and 5% nonfat dry milk. The membranes were then incubated with p-STAT3, STAT3, p-EGFR, EGFR, p-Src, Src, and α-tubulin antibodies(Cell Signaling, Danvers, MA). For surface staining of CD44, cells were harvested and washed with PBS supplemented 0.5% FBS. PE-conjugated anti-CD44 and PE- isotype control(BD Pharmingen, San Jose, CA) were used and incubated at 4℃ for 30min, after 3 washes and de- tected by FACSCalibur(BD Bioscience, San Jose, CA). Exper- iments were repeated at least three times.
Results
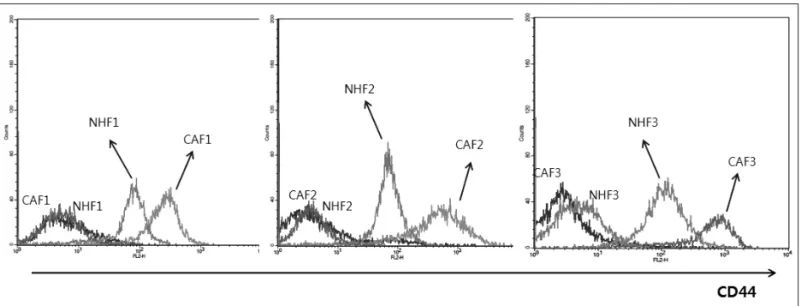
1. CAFs showed higher CD44 expression than did normal fibroblasts from corresponding patients After collecting tissues, primary cultures were performed to sort CAFs from tumor tissues and NHFs from normal tis- sues harvested from patients. To verify the identity of fibro- blasts, we examined the morphology of the grown cells un- der a microscope(Supplementary Data 1) and checked the expression of α-SMA and vimentin, the markers of CAFs (Supplementary Data 2). CD44 expression of CAFs was rela- tively higher than that of NHFs in all three sets of samples(Fig.
1). In addition, CD44 was also found to be expressed in the head and neck cancer cell lines SNU-1041, SNU-1066 and SNU-1076(supplementary data 3).
2. CAFs increased cancer cell motility
To investigate the role of CAFs, which have higher levels of CD44 expression, three cancer cell lines were co-cultured with
CAFs or NHFs. During co-culture, no significant differences were observed in cell growth(data not shown). However, the motility of cancer cells was significantly different in two con- ditions. As shown in the wound healing assay(Fig. 2), the gap made by the scratch was filled more efficiently when cancer cells were co-cultured with CAFs than with NHFs, as demon- strated in the case of SNU-1041 and SNU-1066. Since SNU- 1076 has a high preexisting motility, the motility of SNU-1076 did not show a difference in two conditions. SNU-1076 cells showed a prompt and complete filling of the gap, even with- out any fibroblasts.
3. Increased motility of cancer cells can be based on the action of CD44
To determine the effect of CD44 in the CAFs to the motil- ity of cancer cells, CD44 in the CAFs was blocked by a neu- tralizing antibody. When cancer cells were co-cultured with CAFs, CAFs were pre-treated with 1µg of CD44-neutraliz- ing Ab. As shown in Fig. 3, the scratch is hardly filled in cham- bers with CAFs pre-treated with the blocking antibody. CAF1 and CAF2 were shown similar results in co-culture with SNU- 1041 and SNU-1066. And then, the 3-dimensional motility of cancer cells were evaluated with colony forming assay using soft agar. Colony numbers were higher in the chambers with cancer cells co-cultured with CAFs than in the control cham- bers without CAFs(Fig. 4). Unanchored single cells cultured for two weeks on semi-solid agar should result in cell death by anoikis, however, SNU-1041 and SNU-1066 survived two weeks without cell death. These experiments clearly demon- strated that the invasiveness of cancer cells became stronger with coexistence of CAFs. To confirm this invasiveness is caused by CD44 expression on CAFs, we compared the mi- gration with or without a CD44-neutralizing antibody. The number of cancer cells migrated to the lower chamber were decreased when the CAFs treated with the CD44-neutraliz- ing antibody, compared with the control treated with control IgG(Fig. 5). In this experiment, CAFs were distinguishable from cancer cells because of the pale staining of CAFs. In Fig. 5A, two different areas can be distinguishable according to the staining strength. One is a region of strong staining colonies which are cancer cells and the other is pale staining region which is the CAF stained region. This observation strongly suggests that in mixed culture of CAFs and cancer cells, CD44 on CAFs may mediate cancer cell’s migration and aggressiveness.
4. Co-culture with CAF increase CD44 expression on cancer cells
Since we already found that CD44 was expressed on the
cancer cell themselves, we checked the change of CD44 ex- pression on cancer cells when co-cultured with CAFs. The longer cancer cells were co-cultured with CAFs, the more ex- pression of CD44 was observed on both cancer cells, SNU- 1041 and SNU-1066(Fig. 6A and B). Additionally, co-culture with CAFs lead to the change of other molecules associated with tumor microenvironment. Expression of p-STAT3 was significantly increased in all of the samples co-cultured with
CAFs. Expression of p-Src and p-EGFR showed increase in most of the samples when compared to the control cancer cells(Fig. 6C).
Discussion
This is the first study showing that CAFs have higher CD44 expression levels than do corresponding normal fibroblasts
Fig. 1. The expression of CD44 is higher in CAFs than in NHFs. Three sets of CAFs and NHFs from three patients were examined by FACS analysis to measure CD44 expression. In all cases, CAFs exhibited higher expression of CD44 than did corresponding NHFs.
CAF1, CAF2 and CAF3 annotates three CAFs from 3 head and neck cancer patients, whereas NHF1, NHF2 and NHF3 three normal fibroblasts from the corresponding patients. One hundred thousand cells were incubated with CD44 antibody for 1 hour in 4℃.
Fig. 2. The motility of cancer cells increased by co-culture with CAFs. A wound healing assay was performed to determine whether CAFs influence cancer cell motility. Three head and neck cancer cell lines, SNU-1041, SNU-1066 and SNU-1076, were used. When cancer cells were co-cultured with CAFs in transwell dishes, the scratch in the bottom chambers of transwells with CAFs were filled more efficiently than transwells with corresponding NHFs in SNU-1041 and SNU-1066. SNU-1076, which has a higher expression level of CD44 than the other cancer cell lines, showed no definite differences between CAFs and NHFs. Cells were seeded in the 24 well plate and a straight scratch was made with a pipette tip on the next day. CAFs or NHFs were then placed in the upper chambers of the transwell dish.
from the same patient. The expression of CD44 in the cancer cells was significantly increased by the presence of CAFs compared with the presence of NHFs from the same patient.
Blocking CD44 activity in CAFs reduced the motility and in- vasiveness of cancer cells.
With a modified migration assay using soft agar, we dem- onstrated distinct differences in cell migration and coloniza- tion between cells cultured with or without CAFs. In most ex- perimental systems, it is difficult to keep cancer cells alive in soft agar without attachment for two weeks. If a single can- cer cell is capable of invading this semi-solid, three-dimen- sional environment without attaching for two weeks and ar- rives at the bottom of the transwell plate, that cell can make a colony. Comparing the lower-chamber colony formation asso- ciated with cancer cells cultured with or without CAFs clearly
demonstrated the effect of CAFs on the mobility and invasive- ness of cancer cells.
A study reported that fibroblasts from normal breast tis- sues inhibited the proliferation of MCF-7 cells under co-cul- ture experiments.19) However, when we co-cultured cancer cells with fibroblasts from normal tissue, we did not observe an inhibitory effect of NHFs on cancer cells(data not shown).
In a recent study, direct cell contacts between tumor cells and stroma dramatically enhanced proliferation in pancreatic cells and breast-cancer cells.20) When cells were treated with con- ditioned medium from CAFs, no proliferation of cancer cells was observed in our study(data not shown). However, when co-cultured in a transwell system, proliferation of cancer cells increased by about 5 percent(data not shown). These observa- tions strongly suggest that CAFs and cancer cells communi-
Fig. 3. Blocking CD44 activity in CAFs decreased cancer cell motility. In a transwell system, cancer cells were seeded on the bottom chamber of 24 well plate and CAFs, which were pre-treated with 1 µg of CD44-neutralizing antibody for 30 min, were placed on the upper chamber. Two or three days after making scratches, cell motility was checked by microscope. When CAFs were treated with CD44-neutralizing antibody, cell motility decreased below the levels exhibited by the group treated with IgG control antibody. All pictures used same magnification.
cate with each other when they physically associate.
Blocking CD44 with neutralizing antibody clearly demon- strated that CD44 may play a role in the interaction between cancer cells and the tumor microenvironment. Moreover, as
shown in Fig. 6, we found that increased periods of co-cultur- ing CAFs and cancer cells led to enhanced expression of CD44 from cancer cells. This might imply that cancer cells acquire traits of cancer stem cells through crosstalk with CAFs. To
Fig. 4. CAFs stimulated cancer cell migration and colony formation. By adding soft agar to the bottom chambers of transwell dishes, we checked both the migration ability and colony-forming ability of cancer cells. When we checked the formation of colonies in the bottom chambers by staining with crystal violet two weeks after seeding cells in the upper chamber, cancer cells mixed with CAFs exhibited a notably increased ability to migrate to the bottom chamber and form colonies. A : SNU-1041 itself has the ability to migrate to the bottom of the dishes when co-cultured with CAF1, CAF2 and CAF3, the ability of migration was increased. B : SNU- 1066 does not have the ability to migrate. However, when co-cultured with CAF1, CAF2 and CAF3, the SNU-1066 gained the ability to migrate which is demonstrated by colony formation.
B A
Fig. 5. Blocking CD44 on CAFs decreased cancer cell migration and colony formation. Using the same experimental approach as outlined in Fig. 4, we tried to block CD44 activity using neutralizing antibody. Before mixing cancer cells and CAFs, CAFs were pre-in- cubated either with 1 µg isotype IgG antibody, as a control, or with neutralizing anti-CD44 antibody. The formation of colonies in bottom chamber of 6 well plate after two weeks was reduced for chambers treated with anti-CD44 antibody.
suppress the aggressiveness of cancer cells and the rate of me- tastasis, CD44 should be monitored in CAFs as a significant target in cancer treatment.
Conclusions
CAFs play an important role in controlling the motility of cancer cells in the tumor microenvironment. CD44 can be an important marker of CAFs’ function and may mediate this interaction between the CAFs and cancer cells. The further
study on the biological role and mechanism of action of CD44 in CAFs and cancer cells are warranted.
References
1) Sherman L, Sleeman J, Dall P, Hekele A, Moll J, Ponta H, et al.
The CD44 proteins in embryonic development and in cancer.
Curr Top Microbiol Immunol. 1996;213(Pt 1):249-269.
2) Hinz B, Phan SH, Thannickal VJ, Galli A, Bochaton-Piallat ML, Gabbiani G. The myofibroblast: one function, multiple origins.
Fig. 6. CAFs influenced expression of CD44 and other tumor-promoting molecules from cancer cells. A and B : A half million cancer cells were gathered after co-culture for one day or seven days in transwell dishes. CD44 expression was measured by FACS analysis.
Increased periods of co-culturing CAFs and cancer cells led to increased expression of CD44 from cancer cells. Cells were incubat- ed with CD44 antibody and counted by FACS analysis. C : Co-cultured cancer cells were gathered to determine the change in sig- naling molecules by Western blot analysis. When co-cultured in 6 well transwell dishes, signaling molecules known to be favorable for tumor progression, such as p-EGFR, p-Src and p-STAT3, were found to be upregulated. The expression levels varied depending on cancer cell line or source of CAFs.
A B
C
Am J Pathol. 2007;170(6):1807-1816.
3) Bhowmick NA, Neilson EG, Moses HL. Stromal fibroblasts in cancer initiation and progression. Nature. 2004;432(7015):332- 337.
4) Ostman A, Augsten M. Cancer-associated fibroblasts and tu- mor growth--bystanders turning into key players. Curr Opin Genet Dev. 2009;19(1):67-73.
5) Kalluri R, Zeisberg M. Fibroblasts in cancer. Nat Rev Cancer.
2006;6(5):392-401.
6) Orimo A, Gupta PB, Sgroi DC, Arenzana-Seisdedos F, Delau- nay T, Naeem R, et al. Stromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogen- esis through elevated SDF-1/CXCL12 secretion. Cell. 2005;121(3):
335-348.
7) Aruffo A, Stamenkovic I, Melnick M, Underhill CB, Seed B.
CD44 is the principal cell surface receptor for hyaluronate.
Cell. 1990;61(7):1303-1313.
8) Acharya PS, Majumdar S, Jacob M, Hayden J, Mrass P, Weninger W, et al. Fibroblast migration is mediated by CD44-dependent TGF beta activation. J Cell Sci. 2008;121(Pt 9):1393-1402.
9) Foster LC, Arkonac BM, Sibinga NE, Shi C, Perrella MA, Haber E. Regulation of CD44 gene expression by the proinflammatory cytokine interleukin-1beta in vascular smooth muscle cells. J Biol Chem. 1998;273(32):20341-20346.
10) Legg JW, Lewis CA, Parsons M, Ng T, Isacke CM. A novel PKC- regulated mechanism controls CD44 ezrin association and di- rectional cell motility. Nat Cell Biol. 2002;4(6):399-407.
11) Morrison H, Sherman LS, Legg J, Banine F, Isacke C, Haipek CA, et al. The NF2 tumor suppressor gene product, merlin, me- diates contact inhibition of growth through interactions with
CD44. Genes Dev. 2001;15(8):968-980.
12) Marhaba R, Zoller M. CD44 in cancer progression: adhesion, migration and growth regulation. J Mol Histol. 2004;35(3):211- 231.
13) Godar S, Ince TA, Bell GW, Feldser D, Donaher JL, Bergh J, et al. Growth-inhibitory and tumor- suppressive functions of p53 depend on its repression of CD44 expression. Cell. 2008;134(1):
62-73.
14) Wang SJ, Wong G, de Heer AM, Xia W, Bourguignon LY. CD44 variant isoforms in head and neck squamous cell carcinoma progression. Laryngoscope. 2009;119(8):1518-1530.
15) Prince ME, Ailles LE. Cancer stem cells in head and neck squa- mous cell cancer. J Clin Oncol. 2008;26(17):2871-2875.
16) Ward RJ, Dirks PB. Cancer stem cells: at the headwaters of tu- mor development. Annu Rev Pathol. 2007;2:175-189.
17) Jenq RR, van den Brink MR. Allogeneic haematopoietic stem cell transplantation: individualized stem cell and immune ther- apy of cancer. Nat Rev Cancer. 2010;10(3):213-221.
18) Messadi DV, Bertolami CN. CD44 and hyaluronan expression in human cutaneous scar fibroblasts. Am J Pathol. 1993;142(4):
1041-1049.
19) Dong-Le Bourhis X, Berthois Y, Millot G, Degeorges A, Sylvi M, Martin PM, et al. Effect of stromal and epithelial cells de- rived from normal and tumorous breast tissue on the prolifer- ation of human breast cancer cell lines in co-culture. Int J Can- cer. 1997;71(1):42-48.
20) Fujita H, Ohuchida K, Mizumoto K, Egami T, Miyoshi K, Mori- yama T, et al. Tumor-stromal interactions with direct cell con- tacts enhance proliferation of human pancreatic carcinoma cells. Cancer Sci. 2009;100(12):2309-2317.
Supplementary
Supplementary 2. Western blot analysis showed that CAFs express α-SMA and vimentin, which are representative markers of CAFs.
However, no expression was observed in the SNU-1041 and SNU-1066 head and neck cancer cell lines.
Supplementary 1. We confirmed the morphology of fibroblasts during primary cell culture. The CAFs were characterized by their long and pointed shape.
Supplementary 3. Three head and neck cancer cell lines were tested by FACS analysis to characterize CD44 expression. While high expression was observed in all cell lines, SNU-1076 exhibited higher expression than the other cell lines. 5×105 cells were incubated with CD44 antibody for 1 hour in 4℃.