kfas
10
0
0
전체 글
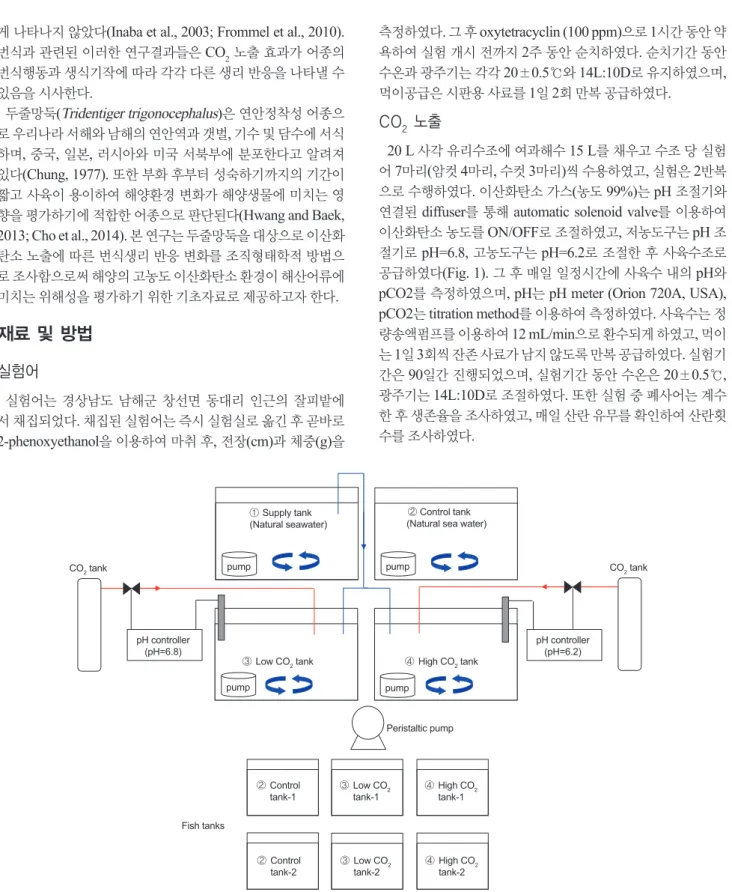
(2) 55. 이산화탄소 노출에 따른 두줄망둑의 번식특성. 게 나타나지 않았다(Inaba et al., 2003; Frommel et al., 2010). 번식과 관련된 이러한 연구결과들은 CO2 노출 효과가 어종의 번식행동과 생식기작에 따라 각각 다른 생리 반응을 나타낼 수 있음을 시사한다. 두줄망둑(Tridentiger trigonocephalus)은 연안정착성 어종으 로 우리나라 서해와 남해의 연안역과 갯벌, 기수 및 담수에 서식 하며, 중국, 일본, 러시아와 미국 서북부에 분포한다고 알려져 있다(Chung, 1977). 또한 부화 후부터 성숙하기까지의 기간이 짧고 사육이 용이하여 해양환경 변화가 해양생물에 미치는 영 향을 평가하기에 적합한 어종으로 판단된다(Hwang and Baek, 2013; Cho et al., 2014). 본 연구는 두줄망둑을 대상으로 이산화 탄소 노출에 따른 번식생리 반응 변화를 조직형태학적 방법으 로 조사함으로써 해양의 고농도 이산화탄소 환경이 해산어류에 미치는 위해성을 평가하기 위한 기초자료로 제공하고자 한다.. 재료 및 방법 실험어 실험어는 경상남도 남해군 창선면 동대리 인근의 잘피밭에 서 채집되었다. 채집된 실험어는 즉시 실험실로 옮긴 후 곧바로 2-phenoxyethanol을 이용하여 마취 후, 전장(cm)과 체중(g)을. 측정하였다. 그 후 oxytetracyclin (100 ppm)으로 1시간 동안 약 욕하여 실험 개시 전까지 2주 동안 순치하였다. 순치기간 동안 수온과 광주기는 각각 20±0.5℃와 14L:10D로 유지하였으며, 먹이공급은 시판용 사료를 1일 2회 만복 공급하였다.. CO2 노출 20 L 사각 유리수조에 여과해수 15 L를 채우고 수조 당 실험 어 7마리(암컷 4마리, 수컷 3마리)씩 수용하였고, 실험은 2반복 으로 수행하였다. 이산화탄소 가스(농도 99%)는 pH 조절기와 연결된 diffuser를 통해 automatic solenoid valve를 이용하여 이산화탄소 농도를 ON/OFF로 조절하였고, 저농도구는 pH 조 절기로 pH=6.8, 고농도구는 pH=6.2로 조절한 후 사육수조로 공급하였다(Fig. 1). 그 후 매일 일정시간에 사육수 내의 pH와 pCO2를 측정하였으며, pH는 pH meter (Orion 720A, USA), pCO2는 titration method를 이용하여 측정하였다. 사육수는 정 량송액펌프를 이용하여 12 mL/min으로 환수되게 하였고, 먹이 는 1일 3회씩 잔존 사료가 남지 않도록 만복 공급하였다. 실험기 간은 90일간 진행되었으며, 실험기간 동안 수온은 20±0.5℃, 광주기는 14L:10D로 조절하였다. 또한 실험 중 폐사어는 계수 한 후 생존율을 조사하였고, 매일 산란 유무를 확인하여 산란횟 수를 조사하였다.. ② Control tank (Natural sea water). ① Supply tank (Natural seawater). pump. pump. CO2 tank. pH controller (pH=6.8). ③ Low CO2 tank pump. CO2 tank. ④ High CO2 tank. pH controller (pH=6.2). pump. Peristaltic pump. ② Control tank-1. ③ Low CO2 tank-1. ④ High CO2 tank-1. ② Control tank-2. ③ Low CO2 tank-2. ④ High CO2 tank-2. Fish tanks. Fig. 1. Schematic diagram of CO2-exposure experiment for chameleon goby Tridentiger trigonocephalus. Control tanks were natural seawater flow (②), high CO2 tanks were pH 6.2 regulated by CO2 tank and pH controller (④), low CO2 groups pH 6.8 regulated by CO2 tank and pH controller (③). Water flows were supplied with peristaltic pump..
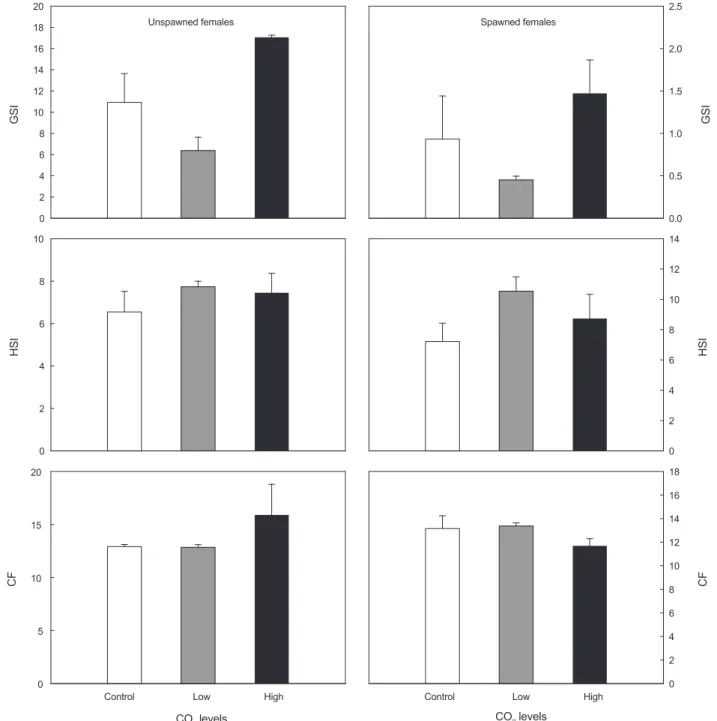
(3) 56. 8.0. 350. 7.8. 300. 7.6. 250. pH. 7.4. Concentrations of CO2 (ppm). 황인준ㆍ최상준ㆍ백혜자. 200. 7.2. 150. 7.0. 100. 6.8. 50. 6.6 Controls. Low. High. Controls. 100. CO2 treatment. Low. High. 0. CO2 treatment. GSI (%)=(gonad weight/body weight)×100 14 HSI (%)=(liver weight/body weight)×100 12 CF=(body weight/standard length3)×1000. 0. 10. 적출된8 조직은 Bouin's solution에 24시간 고정하였고, 수세 및 탈수6 과정을 거쳐 paraffin에 포매한 후, 4-6 µm의 두께로 4 하였다 제작된 조직 표본은 연속절편 . Mayer's hematoxylin 2 eosin (H-E)으로 비교염색을 실시한 후 광학현미경 과 0.5% 0 (OLYMPUS BX-50, Japan) 하에서 관찰하였다. 10. 통계분석. HSI. 실험어의 생체지수는 모두 평균±표준오차로 나타내었으며, 8 Kruskal-Wallis 검정과 Bonferroni 다수 비교 검정을 통해 95% 신뢰수준에서 CO2 노출에 따른 생체지수 변화의 유의차 검정 6 을 실시하였다.. 결. 40 0. 4. 과 2. 이산화탄소 노출에 따른 생존율 및 산란횟수 두줄망둑의 농도별 CO2 노출 기간 동안, 사육수의 pH는 대조 0 구에서 7.86±0.01, CO2 저농도구에서는 7.02±0.02 그리고 20 CO2 고농도구에서는 6.62±0.01의 범위로 나타났다(Fig. 2). CO2 농도는 대조구에서 51.18±2.56 ppm, CO2 저농도구에서는 15. 2.5. Controls females Spawned. Low. CF. 2.0. 1.5. Controls. Low. High. CO2 levels. 1.0. Fig. 3. Effects of CO2 on survival rates of chameleon goby Tridentiger trigonocephalus. Values are mean±SE from0.5 duplicate. 0.0. 7. 14 6 12 5 10. 7 4. 8. 6 3. 6. 5 2. 4. 4 1 3 0. 2 Control. 2. Low. High. CO2 levels. 0 18. 1. Fig. 4. Number of cumulative spawning of chameleon 16 goby Tridentiger trigonocephalus during exposure periods. 0 14 Control. Low. CO2 levels 10. High. CO2 levels. 20. Number of cumulative Number spawning of cumulative spawning. GSI. 16. 60 20. GSI. 18. 80 40. HSI. 이산화탄소 노출 90일 후, 생존한 개체를 채집하여 전장과 무게를 측정하였고 각각의 개체에서 간과 생식소를 적출 하여 0.01 g까지 무게를 측정 한 후 다음의 식에 의거하여 생식소 중량지수(gonadosomatic index, GSI), 간중량지수(hepatosomatic index, HSI), 그리고 비만도(condition factor, CF)를 산 20 출하였다. Unspawned females. 100 60. High. 12 10 8 6. CF. 생체지수, 간과 생식소의 조직학적 관찰. Survival rate (%) Survival rate (%). Fig. 2. Changes in pH and concentrations of CO2 during exposure period. Values 80 are means±SE..
(4) 57. 이산화탄소 노출에 따른 두줄망둑의 번식특성. 20. 2.5 Unspawned females. 18. Spawned females. 16. 2.0. 14 1.5. GSI. GSI. 12 10 8. 1.0. 6 4. 0.5. 2 0. 0.0. 10. 14 12. 8. 10 6. 6. 4. HSI. HSI. 8. 4 2. 2. 0. 0 18. 20. 16 14. 15. 10. 10. 8. CF. CF. 12. 6 5. 4 2. 0. Control. Low. High. CO2 levels. Control. Low. High. 0. CO2 levels. Fig. 5. Effects of CO2 on GSI, HSI and CF of female chameleon goby Tridentiger trigonocephalus. Values are mean±SE from duplicate. GSI, gonadosomatic index; HSI, hepatosomatic index; CF, condition factor.. 156.31±7.90 ppm 그리고 CO2 고농도구에서는 274.17±6.51 ppm의 범위로 나타났다. 90일간의 노출 기간 동안, 실험어의 생존율을 살펴보면, 대조구에서는 85.71 %, CO2 저농도구에서 는 64.29±7.14 % 그리고 CO2 고농도구에서는 42.86±42.86 %로 나타나, CO2 농도가 증가할수록 생존율이 감소하는 경향 을 보였으나 통계적 유의차는 나타나지 않았다(Fig. 3). 90일간 의 노출 기간 동안 일부 개체가 산란하였는데, 대조구에서는 단. 1회만 관찰되었고, CO2 저농도구에서는 총 6회 그리고 CO2 고 농도구에서는 총 4회의 산란이 관찰되었다(Fig. 4).. 두줄망둑의 생체지수 변화 90일간의 CO2 농도별 노출 종료 후, 암수별 GSI, HSI 그리 고 CF를 산출하였다. 암컷의 경우, 산란한 개체와 산란하지 않은 개체를 분리하여 나타냈다(Fig. 5). 산란하지 않은 암컷.
(5) 황인준ㆍ최상준ㆍ백혜자. 1.0. 0.8. GSI. 0.6. 0.4. 0.2. 0.0 8. 6. HSI. 의 GSI는 대조구에서 10.92±2.71이었으며, CO2 저농도구에 서는 6.38±1.53으로 다소 감소하였고 CO2 고농도구에서는 17.06±0.20으로 대조구에 비해 높은 값을 나타냈으나 통계 적 유의차는 나타나지 않았다. HSI는 대조구에서 6.55±0.97, CO2 저농도구에서는 7.74±0.32 그리고 CO2 고농도구에서 는 7.45±0.92로 대조구에 비해 다소 증가하였으나 통계적 유 의차는 나타나지 않았다. CF는 대조구에서 12.91±0.20이었 고, CO2 저농도구에서는 12.85±0.31로 대조구와 유사하였고 CO2 고농도구에서는 15.90±2.89로 대조구보다 증가하였으나 통계적 유의차는 나타나지 않았다. 산란한 암컷의 GSI는 대조구에서 0.93±0.51, CO2 저농도 구에서는 0.45±0.04 그리고 CO2 고농도구에서는 1.47±0.39 로 나타나 CO2 노출에 따른 GSI의 뚜렷한 변화는 나타나지 않았다. HSI는 대조구에서 7.22±1.21, CO2 저농도구에서 10.54±0.94 그리고 CO2 고농도구에서 8.73±1.60으로 CO2 노출에 의해 HSI가 대조구에 비해 다소 증가하였으나 통계적 유의차는 나타나지 않았다. CF는 대조구에서 13.15±1.09, CO2 저농도구에서는 13.37±0.27 그리고 CO2 고농도구에서 는 11.69±0.61로 나타났다. 수컷의 GSI는 대조구에서 0.76±0.13이었으나, CO2 저농도 구에서 0.43±0.16 그리고 CO2 고농도구에서 0.51±0.26으로 CO2 노출에 의해 GSI가 다소 감소하는 경향을 보였으나 통계 적 유의차는 나타나지 않았다. HSI는 대조구에서 5.90±0.46, CO2 저농도구에서 5.41±0.43이었으나 CO2 고농도구에서 3.12±0.72로 대조구에 비해 유의하게 감소한 것으로 나타났다 (P<0.05). CF 변화에서도 대조구에서 13.15±1.09, CO2 저농 도구에서 13.37±0.27이었으나 CO2 고농도구에서는 11.69± 0.61로 대조구에 비해 CF가 유의하게 감소한 것으로 나타났다 (P<0.05).. *. 4. 2. 0 16 14 12. *. 10. CF. 58. 8. 간과 생식소의 조직학적 관찰. 6. 농도별 CO2에 90일간 노출 후, 산란하지 않은 성숙한 암컷의 간과 생식소의 조직학적 관찰을 살펴보면, 난소 내 난모세포의 경우 대조구와 농도별 CO2 처리구에서의 조직학적 관찰 결과, 뚜렷한 차이는 관찰되지 않았다(Fig. 7A, 7B, 7C). 간 조직 관 찰결과, 대조구를 포함한 모든 실험구에서 다소 팽창한 지방구 (lipid droplet)에 의해 간세포는 관찰이 잘 되지 않았으나 농도 별 CO2 노출에 따른 간세포 또는 지방구의 변화는 뚜렷하게 나 타나지 않았다(Fig. 7D, 7 E, 7 F). 산란한 암컷의 간과 생식소의 조직학적 관찰을 살펴보면, 난 소에서는 대조구와 실험구 모두에서 난소 내 성숙난이 방란 후 주변인기 단계의 어린 난모세포들이 드문드문 분포하였다 (Fig. 8A, 8B, 8C). 간 조직 관찰 결과, 대조구에서는 지방구들 이 타원형으로 비교적 균일하고 치밀한 배열을 보인 반면, CO2 처리구에서는 지방구들이 다소 일그러진 형태로 관찰되었다 (Fig. 8D, 8E, 8F).. 4 2 0. Control. Low. High. CO2 levels. Fig. 6. Effects of CO2 on GSI, HSI and CF of male chameleon goby Tridentiger trigonocephalus. Values are mean±SE from duplicate. Asterisks indicate significant differences compared to controls (P<0.05). GSI, gonadosomatic index; HSI, hepatosomatic index; CF, condition factor.. 수컷의 간과 정소의 조직학적 관찰을 살펴보면, 정소 조직에 서는 대조구와 실험구 모두에서 정모세포, 정세포, 정자무리들 이 관찰되었다. 그러나 CO2 처리구에서 정소소엽 내에 관찰되 는 정자들이 대조구에 비해 다소 적은 것으로 나타났다(Fig..
(6) 59. 이산화탄소 노출에 따른 두줄망둑의 번식특성. A. D. 600 µm. 50 µm E. B. 600 µm C. 50 µm F. 600 µm. 50 µm. Fig. 7. Histological observations of ovary and liver from unspawned females exposed to different levels of CO2. A; Ovary from controls, B; Ovary from low CO2 treatment, C; Ovary from high CO2 treatment, D; Liver from controls, E; Liver from low CO2 treatment, F; Liver from high CO2 treatment.. 9A, 9B, 9C). 간조직에서는 대조구에서 타원형의 균일한 크기 의 지방구들이 치밀한 구조로 관찰되었으나, CO2 처리구에서 는 지방구들의 형태와 배열이 다소 불규칙한 형태로 나타났으 며 일부 지방구들은 서로 융합된 형태로도 관찰되었다(Fig. 9D, 9E, 9F).. 고. 찰 A. 본 연구결과 CO2 농도 증가에 따른 두줄망둑의 생존율은 통 계적 유의차는 나타나지 않았지만 감소하는 경향을 보임으로 써 고농도의 CO2 노출은 두줄망둑에 독성 효과를 나타내는 것 으로 판단된다. Ishimatsu et al. (2004)에 의하면 HCl과 CO2. 50 µm. 를 각각 첨가후 동일한 pH로 조절하여 참돔의 난과 부화 자어 의 생존율을 관찰한 결과, HCl 처리구에서는 90% 이상의 생존 율을 보였으나, CO2 처리구에서는 10% 미만의 생존율을 나타 내어 어류의 초기 생활사에 대한 CO2의 독성을 제시하였다. 그 러나 CO2 농도 증가에 의해 감소된 생존율의 원인에 대해서는 아직까지 명확하게 밝혀진 바가 없으나 넙치(Paralichthys olivaceus)와 방어(Seriola quinqueradiata)에서 CO2 농도 증가에 의한 혈액 pH의 저하, CO2 노출에 따른 심장 수축과 산소 공급 D 저하가 그 원인이라고 하였다(Hayashi et al., 2004; Lee et al., 2004). 향후 연구에서 두줄망둑의 CO2 노출에 의한 혈액 pH 변 화, 혈액 내 산소 농도 분석 등이 요구된다.. 50 µm.
(7) 60. 황인준ㆍ최상준ㆍ백혜자. A. D. 50 µm. 50 µm E. B. 50 µm. 150 µm C. F. 150 µm. 50 µm. Fig. 8. Histological observations of ovary and liver from spawned females exposed to different levels of CO2. A; Ovary from controls, B; Ovary from low CO2 treatment, C; Ovary from high CO2 treatment, D; Liver from controls, E; Liver from low CO2 treatment, F; Liver from high CO2 treatment.. 어류는 해양생물 중에서도 비교적 잘 발달된 산-염기 균형 조 절 시스템을 가지고 있어 고농도의 CO2 노출 조건에서도 체내 pH를 복원할 수 있다(Brauner and Baker, 2009; Esbaugh et al., 2012). 그러나 이온 수송을 통한 pH 조절은 에너지 소비가 큰 생리반응이며, 산-염기 균형을 유지하기 위해 사용된 에너지 의 증가는 번식을 포함한 다른 활동에 이용할 수 있는 에너지의 양을 저감시킬 수 있다는 것을 암시한다(Portner et al., 2004; Ishimatsu et al., 2008;A Sokolova et al., 2012). 따라서 pH 항 상성에 대한 증가된 에너지 요구량은 암컷의 번식에 영향을 미 치거나, 산란량 감소, 부화자어의 질에 잠재적인 결과를 초래 할 수 있다. 그러나 본 연구결과, 노출 기간 동안 CO2 처리구에 Sz. 서 대조구에 비해 두줄망둑의 산란횟수가 더 증가하였으며, 저 농도 CO2 처리구에서 산란횟수가 가장 높았다. Hoffman et al. (2010)과 Nielson and Holman (2012) 등은 이와 관련하여 하 나의 가설을 제시하였다. terminal investment 기작 즉, terminal investment를 수행하는 개체는 항상성 유지나 성장보다는 번식을 위한 모든 가용 에너지를 사용한다는 것이다. Terminal investment는 2가지의 큰 특징이 있는데, 1) 성체가 생존하지 못 하고 2) 성체가 열악한 환경에 서식함에 따른 자손의 질(qualD ity)이 저하되는 경우이다(Bonneaud et al., 2003). 본 연구결 과, 두줄망둑의 산란횟수는 뚜렷한 증가를 보였지만 이 결과는 점진적인 생존율 감소 또는 성체의 생체지수의 변화와는 관련.
(8) 이산화탄소 노출에 따른 두줄망둑의 번식특성. A. 61. D. Sz. E. B. Sz. C. F. Sz. Fig. 9. Histological observations of testis and liver from males exposed to different levels of CO2. A; Testis from controls, B; Testis from low CO2 treatment, C; Testis from high CO2 treatment, D; Liver from controls, E; Liver from low CO2 treatment, F; Liver from high CO2 treatment. Scale bars are 20 µm. Sz, spermatozoa; solid arrow head, spermatid; blank arrow head, spermatocyte.. 이 없는 것으로 판단된다. 특히 CO2 노출에 따른 수컷의 HSI와 CF가 감소하였으나 암컷에서는 뚜렷한 차이가 나타나지 않아 CO2 노출에 따른 산란횟수 증가에 terminal investment 기작을 적용하기에는 무리가 있을 것으로 사료된다. CO2 노출에 의한 산란횟수 증가는 hormesis 효과로도 해석 가능하다(Constantini et al., 2010; Schreck CB, 2010). Hormesis는 생물 반응이 외부물질(예, 독성물질)의 투여량에 따라 달 라지는 경우 발생하며, 특정 반응을 나타내는 투여량 이하의 농 도에서 반응이 증가하는 현상이다. 이러한 관점에서 고려해 볼 때, CO2 저농도구에서 증가된 산란횟수는 hormesis 효과에 의 한 것으로 사료된다. 그러나 특정 에너지 소요 또는 소비없이 산. 란을 포함한 일련의 번식행동이 극적으로 증가할 수 있는 기작 을 파악하기 위해서는 관련 내분비 연구가 요구된다. 또한 CO2 노출에 의한 산란횟수 증가와 같은 긍정적인 기능과 함께 부 화자어의 난황 감소(Miller et al., 2013)도 보고되어 있으므로, CO2 노출 이후 F1 세대를 대상으로 한 향후 연구도 필요할 것 으로 생각된다. 90일간의 CO2 노출 종료 후 암컷의 GSI, HSI 그리고 CF 등 은 뚜렷한 변화를 보이지 않았으며, 난소와 간의 조직학적 관찰 에서도 뚜렷한 변화는 나타나지 않았다. 그러나 수컷의 HSI와 CF는 유의하게 감소하였다. 탄산과잉(hypercapnia), 즉 고농도 의 pCO2에 노출된 어류는 호흡계통, 순환계통, 그리고 대사활.
(9) 62. 황인준ㆍ최상준ㆍ백혜자. 동과 같은 생체 내 생리적 기능에 영향을 받고, 이러한 기능들의 변화는 성장 둔화, 번식력 저하, 유영력 감소 등의 결과를 낳는 다(Brauner and Baker, 2009; Ishimatsu et al., 2005). Fivelstad et al. (1999)는 대서양연어를 대상으로 이산화탄소에 노출시 킨 결과, 대조구와 비교하여 저농도(pCO2=19 ppm)와 고농도 (pCO2=32 ppm)에 노출 시킨 실험구에서 유의적인 성장 둔화 및 CF의 감소를 보여 본 연구에서의 두줄망둑 수컷 결과와 유사 하였으며, Smart (1981)와 Fivelstad et al. (2003)은 CF의 감소 는 먹이 섭취의 감소와 관련이 있을 것으로 보고하였다. 수컷의 간 조직 관찰 결과에서도 대조구에서는 지방구들이 균 일하고 치밀한 배열을 보인 반면, CO2 처리구에서는 지방구들 이 다소 팽창하고 불규칙하게 일그러진 형태로 관찰되었다. 따 라서 CO2 노출이 두줄망둑 수컷에서 독성으로 작용하여 간 내 지방구들이 불규칙적인 형태로 다소 팽창하고 이로 인해 HSI와 CF가 감소한 것으로 생각된다. 어류의 간은 담즙의 대사, 유기 화합물과 중금속 해독, 영양 대사 및 영양물질의 체내 저장 그리 고 성호르몬 대사 등의 기능을 담당한다(Health, 1995; Hinton et al., 2001). 어류의 간조직이 오염물질 노출에 따른 가장 일반 적인 반응은 조직 내 글리코겐과 지질의 축적이며, 더 심할 경우 괴사현상이 나타난다고 알려져 있다(Myers et al., 1987; Wolf and Wolfe, 2005). 또한 남극지역에 서식하는 어류의 간세포에 1%의 이산화탄소를 노출시킨 결과, 간세포의 세포 내 단백질 동화작용이 급격히 떨어지고, 이로 인해 간의 정상적인 기능에 제한을 받는 것으로 보고되어 있다(Langenbuch and Pörtner, 2003). 그러나 두줄망둑 암컷에서는 뚜렷한 변화가 나타나지 않았는데, 이 부분에 있어서는 CO2 노출에 대한 민감도가 동 일한 어종이라도 성별에 따라 다르게 나타나는 것으로 판단된 다. 이와 관련한 향후 생리·생화학적 기법을 이용한 실험 분석 이 요구된다. 이산화탄소 농도 증가가 어류의 번식내분비 기작에 미치는 영 향은 현재까지 거의 알려진 바가 없다(Pankhurst and Munday, 2011). 본 연구에서 정소의 조직학적 관찰 결과, 대조구의 경우 정모세포와 정세포의 활발한 분열 증식이 관찰되었으며, 정소 소낭 상피세포벽이 얇아지면서 내강에는 많은 정자들이 관찰 되었다. 그러나 이산화탄소 노출구에서는 정모세포와 정세포 의 분열증식이 이루어지고 있었고, 일부 정자들도 관찰되었으 나, 대조구와 비교해 볼 때 정세포에서 정자로의 형성과정이 다 소 지연되는 것으로 판단되었다. 대구류(Gadus morhua)의 정 액은 CO2 노출에 의한 정자 속도, 운동성 등에 뚜렷한 효과가 나타나지 않았다고 보고되어 있다(Frommel et al., 2010). 한 편, CO2 노출에 의한 가자미류 정액의 운동성 저해가 보고되 어 있는데, 가자미류 정액은 CO2를 HCO3-로 전환하는 효소인 carbonic anhydrase가 다량 함유되어 있으며 이 효소가 정자의 운동성 저해인자라고 제시하였다(Inaba et al., 2003). 그러나 가 자미류를 제외한 다른 어종에서는 정액에 carbonic anhydrase 가 거의 없기 때문에 이 효소에 의한 정자 운동성 저해는 없을. 것이라고 보고하였다(Inaba et al., 2003). 본 연구의 실험어종인 두줄망둑은 수정 시에 수중에 방정하지 않고 암컷이 산란하면 수컷이 산란된 난에 정액을 묻혀서 수정하는 번식특성(Hwang et al., 2011)으로 인해 CO2 노출이 정액에 직접적인 영향을 미 치지는 않을 것으로 판단된다. 그러나 앞서 간 조직 관찰결과와 HSI, CF 감소 등을 고려했을 때, CO2 처리구에서 수컷의 정자 형성이 지연되는 것은 이산화탄소에 노출된 수컷의 생리상태 저하에 따라 정소의 성숙이 지연된 것으로 생각된다.. 사. 사. 이 논문은 부경대학교 자율창의학술연구비(2016년, C-D2016-0262)에 의하여 연구되었음.. References Bonneaud C, Mazuc J, Gonzalez G, Haussy C, Chastel O, Faivre B and Sorci G. 2003. Assessing the cost of mounting an immune response. Amer Nat 161, 367-379. https://doi. org/10.1086/346134. Brauner CJ and Baker DW. 2009. Patterns of acid-base regulation during exposure to hypercarbia in fishes. In: Cardi-Respiratory Control in Vertebrates. Glass ML and Woods SC, eds. Springer, Berlin, Germany, 43-63. Browman HI. 2016. Applying organized scepticism to ocean acidification research. ICES J Mar Sci 73, 529-536. https:// doi.org/10.1093/icesjms/fsw010. Caldeira K and Wickett ME. 2005. Ocean model predictions of chemistry changes from carbon dioxide emissions to the atmosphere and oceans. J Geophys Res 110, C09504. https:// doi.org/10.1029/2004JC002671. Cho HC, Hwang IJ and Baek HJ. 2014. Histological analysis of early gonadal development and sex differentiation in chameleon goby, Tridentiger trigonocephalus. Dev Reprod 18, 51-56. https://doi.org/10.2717/DR.2014.18.1.051. Chung MK. 1977. The fishes of Korea. Inji-sa, Seoul, Korea. Costantini D, Metcalfe NB and Monaghan P. 2010. Ecological processes in a hermetic framework. Ecol Lett 13, 14351447. https://doi.org/ 10.1111/j.1461-0248.2010.01531.x. Dupont S, Dorey N and Thorndyke M. 2010. What meta-analysis can tell us about vulnerability of marine biodiversity to ocean acidification? Estuar Coast Shelf Sci 89, 182-185. https://doi.org/10.1016/j.ecss.2010.06.013. Esbaugh AJ, Heuer R and Grosell M. 2012. Impacts of ocean acidification on respiratory gas exchange in a marine teleost, Opsanus beta. J Comp Physiol B 182, 921-934. https://doi. org/10.1007/s00360-012-0668-5. Esbaugh AJ. 2017. Physiological implications of ocean acidification for marine fish: emerging patterns and new insights. J Comp Physiol B 188, 1-13. https://doi.org.10.1007/s00360017-1105-6..
(10) 이산화탄소 노출에 따른 두줄망둑의 번식특성. Fivelstad S, Olsen AB, Kloften H, Ski H and Stefansson S. 1999. Effects of carbon dioxide on Atlantic salmon (Salmo salar L) smolts at constant pH in bicarbonate rich freshwater. Aquaculture 178, 171-187. https://doi.org/10.1016/ S0044-8486(99)00125-8. Fivelstad S, Olsen AB, Asgard T, Baeverfjord G, Rasmussen T, Vindhelm T and Stefansson S. 2003. Long-term sublethal effects of carbon dioxide on Atlantic salmon smolts (Salmo salar L.): ion regulation, haematology, element composition, nephrocalcinosis and growth parameters. Aquaculture 215, 301-319. https://doi.org/10.1016/S0044-8486(02)00048-0. Forsgren E, Dupont S, Jutfelt F and Amundsen T. 2013. Elevated CO2 affects embryonic development and larval phototaxis in a temperate marine fish. Ecol Evol 3, 3637-3646. https://doi. org/10.1002/ece3.709. Frommel AY, Stiebens V, Clemmesen C and Havenhand J. 2010. Effect of ocean acidification on marine fish sperm (Baltic cod: Gadus morhua). Biogeosci Discuss 7, 58595872. https://doi.org/10.5194/bg-7-3915-2010. Hwang IJ and Baek HJ. 2013. Reproductive cycle of chameleon goby, Tridentiger trigonocephalus in the southern coastal waters of Korea. Dev Reprod 17, 353-361. https://doi. org/10.12717/DR.2013.17.4.3513. Health AG. 1995. Water Pollution and Fish Physiology. 2nd ed. CRC Lewis Publishers, Boca Raton, Florida, U.S.A. Heuer RM and Grosell M. 2014. Physiological impacts of elevated carbon dioxide and ocean acidification on fish. Am J Physiol-Reg I 307, R1061-R1084. https://doi.org/ 10.1152/ ajpregu.00064.2014. Hinton DE, Segner H and Braunbeck T. 2001. Toxic responses of the liver. In: Target Organ Toxicity in Marine and Freshwater Teleosts. Schlenk D and Benson WH, eds. Taylor & Francis, London. UK, 224-268. Hoffman CL, Higham JP, Mas-Rivera A, Ayala JE and Maestripieri D. 2010. Terminal investment and senescence in rhesus macaques (Macaca mulatta) on Cayo Santiago. Behav Ecol 21, 972–978. https://doi.org/10.1093/beheco/arq098. Inaba K, Dreanno C, and Cosson J. 2003. Control of flatfish sperm motility by CO2 and Carbonic anhydrase. Cell Motil Cystoskeleton 55, 174-187. https://dor.org/10.1002/cm. 10119. Ishimatsu A, Kikkawa T, Hayashi M, Lee KS and Kita J. 2004. Effects of CO2 on marine fish: Larvae and adults. J Oceanogr 60, 731-741. https://doi.org/10.1007/s10872-004-5765-y. Ishimatsu A, Hayashi M, Lee KS, Kikkawa T and Kita J. 2005. Physiological effects on fishes in a high-CO2 world. J Geophys Res 110, C09S09. https://doi.org/10.1029/2004JC002564. Ishimatsu A, Hayashi M and Kikkawa T. 2008. Fishes in highCO2, acidified oceans. Mar Ecol Prog Ser 373, 295-302. https://doi.org/ 0.3354/meps07823. Langenbuch M and Po¨rtner HO. 2003. Energy budget of hepatocytes from Antarctic fish (Pachycara brachycephalum. 63. and Lepidonotothen kempi) as a function of ambient CO2: pH-dependent limitations of cellular protein biosynthesis?. J Exp Biol 206, 3895-3903. https://doi.org/10.1242/ jeb.00620. Milazzo M, Cattano C, Alonzo SH, Foggo A, Gristina M, Rodolfo-Metalpa R, Sinopoli M, Spatafora D and Stiver KA. 2016. Ocean acidification affects fish spawning but not paternity at CO2 seeps. Proc R Soc Lon B Biol 283, 20161021. https://doi.org/10.1098/rspb.2016.1021. Melzner F, Gutowska MA, Langenbuch M, Dupont S, Lucassen M, Thorndyke MC, Bleich M and Portner HO. 2009. Physiological basis for high CO2 tolerance in marine ectothermic animals: pre-adaptation through lifestyle and ontogeny?. Biogeosciences 6, 2313-2331. https://doi.org/10.5194/bg-62313-2009. Miller GM, Watson SA, McCormick MI and Munday PL. 2013. Increased CO2 stimulates reproduction in a coral reef fish. Glob Change Biol 19, 3037-3045. https://doi.org/10.1111/ gcb.122. Myers MS, Rhodes LD and McCain BB. 1987. Pathologic anatomy and pattern of occurrence of hepatic neoplasms, putative preneoplastic lesions and other idiopathic hepatic conditions in English sole (Parophrys vetulus) from Puget Sound, Washington. J Natl Cancer Inst 78, 333-363. https:// doi.org/10.1093/jnci/78.2.333. Nielsen ML and Holman L. 2012. Terminal investment in multiple sexual signals: immune-challenged males produce more attractive pheromones. Funct Ecol 26, 20-28. https://doi.org/ 10.1111/j.1365-2435.2011.01914.x. Pankhurst NW and Munday PL. 2011. Effects of climate change on fish reproduction and early life history stages. Mar Freshwater Res 62, 1015-1026. https://doi.org/10.1071/MF10269. Portner HO, Langenbuch M and Reipschlager A. 2004. Biological impact of elevated ocean CO2 concentrations: lessons from animal physiology and earth history. J Oceanogr 60, 705-718. https://doi.org/10.1007/s10872-004-5763-0. Schade FM, Clemmesen C and Wegner KM. 2014. Within- and transgenerational effects of ocean acidification on life history of marine three-spined stickleback (Gasterosteus aculeatus). Mar Biol 161, 1667-1676. https://doi.org/10.1007/ s00227-014-2450-6. Schreck CB. 2010. Stress and fish reproduction: The roles of allostasis and hormesis. Gen Comp Endocrinol 165, 549-556. https://doi.org/10.1016/j.ygcen.2009.07.004. Sokolova IM, Frederich M, Bagwe R, Lannig G and Sukhotin AA. 2012. Energy homeostasis as an integrative tool for assessing limits of environmental stress tolerance in aquatic invertebrate. Mar Environ Res 79, 1-15. https://doi. org/10.1016/j.marenvres.2012.04.003. Wolf JC and Wolfe M. 2005. A brief overview of nonneoplastic hepatic toxicity in fish. Toxicol Pathol 33, 75-85. https://doi. org/10.1080/01926230590890187..
(11)
수치


+4



관련 문서
③ 분자가 반응하기 위해서는 반드시 충돌이 필요하며 그 충돌은 분자가 활성화에너지 이상 의 에너지를 갖고 화학적 반응에 필요한 방향이 알맞을 때 반응을 일으킬
따라서 계산
재무제표
기함수는 x에서의 함수값과 –x에서의 함수값의 부호가 반대 à 원점에 대칭.. 2차 함수의 그래프와
- 축산업으로 인한 환경부담을 낮추고, 사회로부터 인정받아야 중장기적으로 축산업 성장 가능 - 주요과제: 가축분뇨 적정 처리, 온실가스 저감, 축산악취 저감
Our analysis has shown that automation is already widespread among both domestic and foreign investors in Vietnam, and that both groups plan to continue investing
이는 아직 지부지사에서 확인 및 승인이 완료되지 않은 상태. 지부지사에서 보완처리 및 승인처 리 시
[r]
