관련 문서
In the current context, where the interconnectedness of the global economy has intensified greatly and the importance of a collective response to
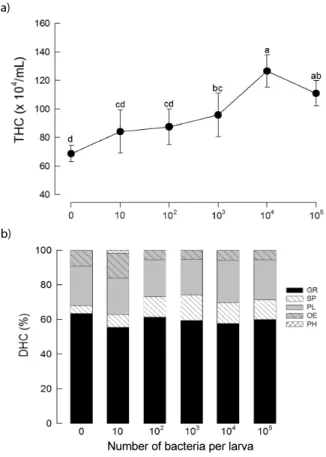
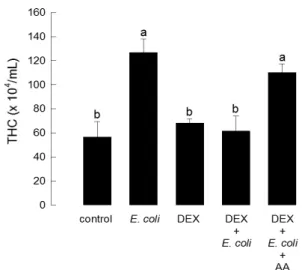
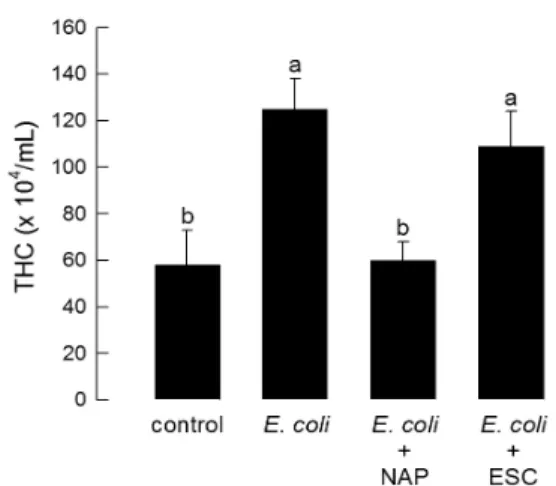
In the API 20NE system, negative reaction for nitrate reduction, indole production, glucose fermentation, arginine dihydrolase, urease acti vity, gelatin hydrolysis,
2재화 2요소 헥셔-올린 모형에서는 어느 한 경제에서 어느 한 요소의 양이 증가하면, 그 요소를 집약적으로 사용하는 산업의 생산량은 증가하고 다른
The index is calculated with the latest 5-year auction data of 400 selected Classic, Modern, and Contemporary Chinese painting artists from major auction houses..
It considers the energy use of the different components that are involved in the distribution and viewing of video content: data centres and content delivery networks
After first field tests, we expect electric passenger drones or eVTOL aircraft (short for electric vertical take-off and landing) to start providing commercial mobility
1 John Owen, Justification by Faith Alone, in The Works of John Owen, ed. John Bolt, trans. Scott Clark, "Do This and Live: Christ's Active Obedience as the
- 우선, 외부 전원이 없는 회로, 즉 초기조건에 의해서 전류, 전압이 결정되 는 회로의 응답 : natural response.. - 다음, 직류 전압이나 전류가