http://bxdoi.org/10.7783/KJMCS.2012.20.5.381
대식세포 Raw 264.7에서 두충의 항염증효과
김영옥*†·이상원*·손상현*·김승유*·오명숙**·김수강***
농촌진흥청 국립원예특작과학원 인삼특작부, **경희대학교 약학대학 한약학과, ***경희대학교 의과대학 고항의학연구소
Anti-inflammatory Effects of Water Extract of Eucommia ulmoides O
LIVERon the LPS-induced RAW 264.7 Cells
Young Ock Kim*†, Sang Won Lee*, Sang Hyun Sohn*, Seung Yu Kim*, Myung Sook Oh** and Su Kang Kim***
*Department of Herbal Crop Research, NIHHS, RDA, Eumseong 369-873, Korea.
**Department of Oriental Pharmaceutical Science, College of Pharmacy Kyung Hee University, Seoul 130-701, Korea.
***Kohwang Medical Research Institute, College of Medicine, Kyung Hee University, Seoul 130-701, Korea.
ABSTRACT :Eucommia ulmoides OLIVER (EU) is a traditional Korean herbal used for the treatment of rheumatoid arthri- tis (RA). In the present study, the molecular pharmacology basis of its anti-inflammatory effect is revealed in this work, EU was studied in lipopolysaccharide (LPS)-activated macrophage cells (RAW 264.7) as an established inflammation model.
After activation, nitric oxide (NO) production and iNOS mRNA were measured by using a colorimetric assay (Griess reagent), and reverse transcription polymerase chain reaction (RT-PCR), respectively. The change in the content of PGE2, TNFα, and IL-6 was concurrently monitored by ELISA. In results, we found that in the concentration range without show- ing cytotoxicity, EU produced a remarkable anti-inflammatory effect and showed a dose-dependent inhibition of LPS- induced NO production. Compared with indomethacin, EU has more potency and a specific action of NO inhibition, PGE2, IL-6, and TNF-α inhibition. These results suggest that EU may be a suitable herbal medicine to yield the greatest anti- inflammatory activity for food additives and medicine.
Key Words : Eucommia ulmoides OLIVER, NO, iNOS, PGE2, IL-6, TNF-α
INTRODUCTION
Nitric oxide (NO), made by inducible NO synthase (iNOS) and excessive NO would produce injurious effects to individuals with chronic inflammation (Jeoung et al., 2009, Hesslinger et al., 2009). In 1998, Drs. Furchgott, Ingnarro and Murad received the Noble Prize in Physiology and Medicine for ‘the first discovery that a gas can act as a signal molecule’, emphasizing that nitric oxide (NO) produced by mammalian cells is an important signaling mediator in immunology, physiology, and neuroscience. The inhibitory activiry of NO production in lipopolysccharide (LPS) activated macrophage cell line, RAW 264.7 from the ethanolic extract of EU. Collectively, NO plays a essential role in endotoxin induced septic shock and death.
Thus, iNOS-derived NO overproduction appears to be a
ubiquitous mediator of a wide range of inflammatory conditions (Sacco et al., 2006).
Marcrophages are immune cells usually dispersed throughout the body. Marcrophage detect pathogenic substances control inflammatory responses (Medzhitov R et al., 1997) using pro-inflammatory mediators such as NO, TNF-α, interleukins (ILs), bacterial endotoxin, lipopolysaccharide (LPS), ionizing radiation, and carcinogens that are often associated with inflammatory diseases or tumorigenesis.
LPS is the primary component of endotoxin and is formed by a phosphoglycolipid (Lee and Cho, 2007) and prohibits macrophage proliferation and activates them to produce proinflammatory factors, which play principal roles in the immune response (Morrison et al., 1987).
EU also called Du-zhong, a dioecious perennial angiosperm, is one of the oldest tonic herbs. The main
†Corresponding author: (Phone) +82-43-871-5585 (E-mail) [email protected] Received 2012 April 7 / 1st Revised 2012 April 18 / Accepted Revised 2012 October 5
components of EU Cortes are 6-10% gutta percha (plant gum), glycoside, alkaloid, pectin, chlorogenic acid, iridoid, cyclopentanoid and dilignan-glycoside (Deyama et al., 2001), The pharmacological studies of EU is various activities such as antioxidant (Hsieh and Yen, 2000) antimicrobial, anti- inflammatory (Kim et al., 2009), antihypertensive (Gu et al., 2011), and immunosuppressive (Zhu et al., 2009) neuroprotective (Kwon et al., 2011) or inhibitory of osteoclast and osteoblastic apoptosis (Lin et al., 2011). Du-zhong tea, the aqueous extract of EU leaves, has been already known as a functional food and commonly used for reduction of hypertension (Greenway et al., 2011). EU leaves have also been used in foods and contain the following active ingredients (Lie et al., 2005), geniposidic acid and asperu- loside as iridoid glycosides, and chlorogenic acid as a cof- feic acid derivative. Based on the traditional uses and known functionalities of EU, we examined for its potential anti-inflammatory activities in vitro assay models evaluating effects on NO, iNOS, PGE2 IL-6 and TNF-α production.
MATERIALS AND METHODS
1. Materials
Indomethacin, and LPS purified from Escherichia coli O111 : B4 were purchased from Sigma-Aldrich (St. Louis, MO, USA). 3-(4,5-dimethylthiazolyl-2)-2,5-diphenyltetrazoliumbromide (MTT, Sigma, USA), Dulbecco’s modified Eagle’s medium (DMEM) and fetal bovine serum (FBS) were from Invitrogen (Carlsbad, CA, USA). The prostaglandin E2 (PGE2) ELISA assay was from R & D Systems (Minneapolis, MN, USA).
Lipopolysacharide (LPS, Sigma, USA), Griess reagent (Sigma, USA), Dimethyl sulfoxide (DMSO, Sigma, USA), RIPA buffer (Pierce, Rockford, IL, USA), protein assay kit (Bio-Rad Laboratories, USA), all other reagents were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Filtered ethanol EU extracts were air dried using a rotary evaporator and powdered.
2. Plant materials
The leaves of EU was purchased from a store at the Gyung Dong herbal market in Seoul, Republic of Korea, and then specimens were taxonomically identified by a oriental doctor, S.W. Lee at the National Institute of Horticultural & Herbal Science, RDA. The voucher specimen (HPR-208) was deposited at the herbarium of Herbal Crop
Research Institute (Eumsung, Republic of Korea). We used water extraction because most traditional oriental herbal materials are decocted with boiling water and because the iridoid glycosides are more soluble in water than in organic solvents. The crushed plant materials (200 g for each) were extracted under reflux with distilled water three times. The combined water extracts were lyophilized, and yields were 23.5% (wt = wt) for EU in the dried state.
They were kept at 80℃ until use.
3. Cell culture
Murine macrophage RAW 264.7 cells (ATCC) were cultured at 37℃ in Dulbecco’s modified Eagles’s medium containing 10% fetal bovine serum, 2 mM glutamate, 100 unit/㎖ of penicillin, and 100 ㎍/㎖ of streptomycin in a humidified incubator with 5% CO2. Cells were incubated with 1㎍/㎖ LPS along with various concentrations of plant extracts for 24 hrs as indicated.
4. MTT cell viability assay
The cytotoxicity of EU was assessed using the microculture tetrazolium [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)]-based colorimetric assay. The remaining cells after the Griess reaction were used for MTT assay.
MTT solution (5㎎/mL) was added to each well (final concentration, 62.5㎍/mL). The degree of mitochondrial respiration, as an indicator of cell viability, was assayed by measuring the mitochondria dependent reduction of MTT to formazan crystals. The absorbance of each well was then read at 570㎚ using the microplate reader (model EL808, Bio-Tek Instrument Inc., Winooski, VT).
The details of this experiment were reported elsewhere (Park et al., 2004).
5. RNA preparation and iNOS mRNA analysis by RT-PCR Cells (6 × 105cells/well) in 6-well plates were treated with and without EU and indomethacin in the presence of LPS for an optimal incubation time of 9 h. The total RNA was isolated by using a Total RNA Miniprep System (Viogene). One microgram of total RNA was used to produce the first strand cDNA pool using the Access RT-PCR System. PCR primer sequences for iNOS gene were 5-CCCTTCCGAAGTTTCTGGCAGCAGC-3 (sense) and 5-GGCTGTCAGAGAGAGCCTCGTGGCTTTGG-3 (antisense).
Primers for β-actin (as a house keeping gene) were 5-
ATGCCATCCTGCGTCTGGACCTGGC-3 (sense) and 5- AGCATTTGCGGTGCACGATGGAGGG-3 (antisense). The expected sizes of PCR products were 499 bp for iNOS gene and 607 bp for β-actin gene. The amplification profile consisted of denaturation at 94℃ for 45 s, annealing at 65℃
for 45 s, and extension at 75℃ for 2 min (15 cycles), and then another denaturation at 94℃ for 45 s, annealing at 67℃
for 45 s, and extension at 75℃ for 2 min (10 cycles). The products were analyzed by electrophoresis on 2% agarose gels containing ethidium bromide and visualized with an ultraviolet transilluminator.
6. NO, PGE2, TNF-α, and IL-6 assays
To investigate NO formation, nitrite (NO2-) is measured since it is a stable, nonvolatile breakdown product of NO.
The determination of nitrite relies on a diazotization reaction with the Griess reagent (1% sulfanilamide, 0.1%
naphthylethylenediamine dihydrochloride, and 2% phosphoric acid). Therefore, the nitrite accumulated in culture medium is a direct indicator of NO production (Green et al., 1982). A 100-µl aliquot of each supernatant from the 96- well plate was mixed with 100µl Griess reagent and incubated at room temperature for 15 min. The concentration of total nitrite was determined by reading the absorbance at 550㎚ and then calculated by a NaNO2 dilution standard curve. The culture medium was collected after LPS treatment for PGE2, TNF-α, and IL-6 assays, respectively.
The time points for PGE2, TNF-α, and IL-6 level were determined by a time course assay (data not shown).
Levels of PGE2, TNF-α, and IL-6 in the culture media were determined with commercial ELISA assay kits (Park et al., 2004).
7. Statistical Analysis
All data were expressed as mean ± SD values which were made in triplicate experiments. Statistical significance was tested by one-way ANOVA (version 20.0 Chicago, IL), and P < 0.05 was considered statistically significant.
RESULTS AND DISCUSSION
Numerous previous studies have shown that certain traditional herbal medicines have therapeutic effects on inflammation. Here, we explored the potential of UE as an anti-inflammatory herbal medicine. Our data demonstrate that EU exhibits pharmacological activity via significant inhibitory effects on the LPS-induced inflammatory mediators and cytokines such as NO, iNOS, PGE2, IL-6, and TNF-α, and in RAW 264.7 cells.
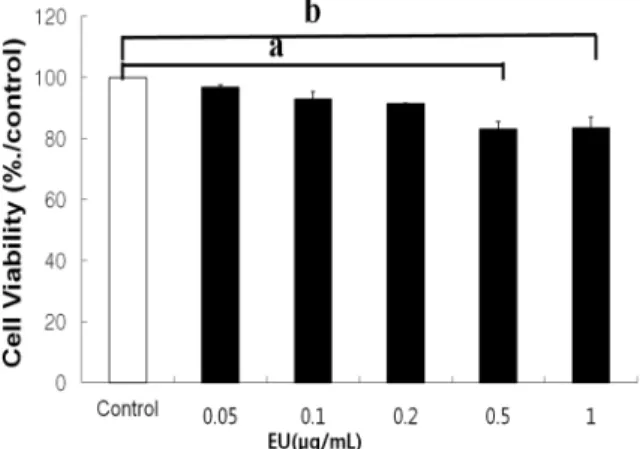
1. Effect of EU extracts on cellular cytotoxicity
Cell viability was tested throughout the experiments using the MTT assay. As shown in Fig. 1, EU did not decrease cell viability but rather protected the toxicity of
Fig. 1. Effect of EU on cell viability. RAW 246.7 cells macrophages were incubated with various concentrations of EU in the presence of 1㎍/mL LPS for 24 hours. Then cell viability was measured by the MTT assay as described in Materials and methods. Values represent the relative viability relative to the control group and are mean ± SD values of three independent experiments. Different letters indicate significantly different values.
Fig. 2. Inhibitory effect of EU on nitric oxide production in LPS- stimulated RAW 264.7 cells. The cells were stimulated with 1㎍/mL of LPS or with LPS plus various concentrations (1㎍/mL, 5 ㎍/mL, 25 ㎍/mL, 125 ㎍/mL, 625㎍/mL) of ethanol-based Eucommia ulmoides OLIVER
extract for 16 h. Nitric oxide production was determined using the Griess reagent method. Different letters indicate significantly different values.
LPS. This result suggests that EU inhibits NO and IL-6 release without affecting cell viability.
2. Effects of EU on NO inhibition
In the EU concentration range showing cytotoxicity from 0.5 to 2㎎/㎖ (Fig. 1), it produced a dose-dependent inhibition of LPS-induced NO production (Fig. 2). Clearly, the remarkable effect on NO inhibition indicates that EU has a potency against inflammation. Over NO production lead to serious diseases like septic shock, stroke, arthritis chronic inflammatory diseases, and autoimmune disease (Bak et al., 2011). It has been reported that NO synthesis is greatly diminished by neutralization with an anti-TNF-α antibody (Kim et al., 1999).
3. Effects of EU on LPS-induced iNOS expression levels in Raw 264.7 cells
LPS is the principal component of endotoxin, arrests macrophage proliferation and activates them to produce
pro-inflammatory factors (Morrison and Ryan, 1987).
Our results showed that LPS enhanced iNOS expression in Raw 264.7 cells and EU suppressed LPS-induced iNOS expression. Therefore, we assumed the effect EU on iNOS expression levels with LPS exposure. Raw 264.7 cells were incubated for 12 h with LPS (1㎍/mL) in the absence or presence of EU (10-100㎍/mL). Our results showed that EU suppresses LPS-induced iNOS expression levels in a dose-dependent manner (Fig. 3).
These results suggests that the inhibition of iNOS expression is important for alleviating inflammation (Lee et al., 2010). Therefore, the inhibition of iNOS expressions may establish an effective new therapeutic vision for the remedy of inflammation and the prevention of inflammatory disease.
Fig. 3. Inhibitory effect of EU on LPS-induced iNOS mRNA levels in Raw 264.7 cells. The cells were pretreated with the EU for 1 h before incubation with LPS (1㎍/mL) for 12 h and total RNA was extracted. iNOS mRNA were analyzed by RT-PCR using specific primers.
Fig. 4. Effect of EU on PGE2 inhibition. RAW 246.7 cells were treated with LPS (1㎍/mL) without and with EU (50, 100, and 200㎍/mL), and indomethacin (0.25 mM) for 12 h prior to PGE2 concentration being measured. Different letters indicate significantly different values.
Fig. 5. Effects of Eu on IL-6 (a) and TNF-α (b) production. RAW 246.7 cells were treated with LPS (1㎍/mL) with and without EU (50㎍/mL), or indomethacin (0.25 mM) for 3 h prior to TNF-α and IL-6 concentration being measured.
Different letters indicate significantly different values.
4. Effects of EU on the PGE2 inhibition
And 0.25 mM of indomethacin was used as a positive control in further studies to allow a comparison with EU anti- inflammatory activity (Shin et al., 2010). An abnormal level of PGE2 via COX activity is known to mediate inflammation.
LPS stimulation significantly increased PGE2 production (p < 0.05, Fig. 4). A dose of EU (0.05㎎/mL) did not affect LPS-induced PGE2 production. The induction of PGE2 was decreased by dose dependent of EU but can be mostly abolished by 0.25㎎/mL of indomethacin. Plainly, EU was less potent than indomethacin in the means of production.
5. Effects of EU on IL-6 and TNF-α production
The cellular IL-6 and TNF-α levels were also assessed using a IL-6 and TNF-α specific ELISA experiment. IL-6 and TNF-α are early secreted cytokines (proinflammatory signals). IL-6 is related to a variety of inflammatory processes, particularly acute inflammation (Diehl et al., 2002) and mediates autoimmune conditions so IL-6 has become a promising new target for immunomodulatory anti-rheumatic therapy (Nishimoto et al., 2006). The inhibition LPS-induced increases in IL-6 and TNF-α has been used to assess the potential anti-inflammatory effects of drug (Kumar et al., 2004) and medicinal herb (Kim et al., 2005). Fig. 5 a, and b showed that LPS significantly stimulated IL-6 and TNF-α production.
EU (50㎍/mL) significantly inhibited the production of IL-6 and TNF-α in LPS-induced RAW264.7 cells (p < 0.05).
The administration of 50㎍/mL EU could produce a greater suppression of IL-6 production (p < 0.05 concentration of EU(50㎍/mL) required to lower IL-6 and TNF-α by 30%
and 20%, respectively, they could be very interesting for rheumatological inflammation treatments.
EU has long been used as traditional oriental herbs as well as ingredients for medicinal food like tea-type beverages.
Furthermore, when conducting in vivo animal experiments with the iridoid-containing herbal samples, the pharmacokinetic properties of iridoid glycosides should be considered.
Therefore, the extract of EU could be a new selective NO inhibitor in the future treatment of arthritis inflammation.
ACKNOWLEDGEMENTS
This research has been supported by the Biogreen 21 Program (Code # PJ007186), Rural Development Administration.
LITERATURE CITED
Bak JP, Son JH, Kim YM, Jung JH, Leem KH, Lee EY and Kim EH. (2011). Suppression of the inflammatory response in macrophages by Paeoniae Radix Rubra. Korean Journal of Medicinal Crop Science. 19:373-379.
Bowen WP and Jerman JC. (1995). Nonlinear-regression using spreadsheets. Trends in Pharmacological Sciences. 16:413-417.
Diehl S and Rincon M. (2002). The two faces of IL-6 on Th1/
Th2 differentiation. Molecular Immunology. 39:531-536.
Deyama T, Nishibe S and Nakazawa Y. (2001). Constituents and pharmacological effects of Eucommia and Siberian ginseng.
Acta pharmacologica Sinica. 22:1057-1070.
Greenway F, Lie Z, Yu Y and Gupta A. (2011). A clinical trial testing the safety and efficacy of a standardized Eucommia ulmoides Oliver bark extract to treat hypertension. Alternative Medicine Review. 16:336-347.
Green LC, Wagner DA, Glogowski J, Skipper PL, Wishnok JS and Tannenbaum SR. (1982). Analysis of nitrate, nitrite, and [15N] nitrate in biological fluids. Analytical Biochemistry.
126:131-138.
Gu J, Wang JJ, Y J, Cui CF, Wu WH, Li L, Wang ZS, Yu M, Gao N, Liu L and Ouyang DS. (2011). Effects of lignans extracted from Eucommia ulmoides and aldose reductase inhibitor epalrestat on hypertensive vascular remodeling. Journal of Ethnopharmacology. 133:6-13.
Hesslinger C, Strub A, Boer R, Urich WR, Lehner MD and Braun C. (2009). Inhibition of inducible nitric oxide synthase in respiratory diseases. Biochemical Society Transactions.
37:886-891.
Hsieh CL and Yen GC. (2000). Antioxidant actions of Du-zhong (Eucommia ulmoides OLIV.) toward oxidative damage in biomolecules. Life Sciences. 66:1387-1400.
Jeoung YJ, Choi SY, An CS, Jeon YH, Park DK and Lim BO.
(2009). Comparative effect on anti-inflammatory activity on the Phellinus linteus and Phellinus linteus grown in geminated brown rice extracts in murine macrophage RAW 264.7 cells.
Korean Journal of Medicinal Crop Science. 17:97-101.
Kim BH, Park KS and Chang Il-Moo. (2009). Elucidation of anti-inflammatory potencies of Eucommia ulmoides bark and Plantago asiatica seeds. Journal of Medicinal Food. 12:764-769.
Kim HM, Moon EJ, Li E, Kim KM, Nam SY and Chun CK.
(1999). The nitric oxide-producing activities of nitric oxide in mouse peritoneal macrophages after priming with interferon- gamma by the stem of Sinomenium acutum. Journal of Ethnopharmacology. 66:311-317.
Kim SJ, Jeong HJ, Moon PD, Lee KM, Lee HB, Jung HJ, Rhee HK, Yang DC, Hong SH and Kim HM. (2005). Anti- inflammatory activity of gumiganghwaltang through the inhibition of nuclear factor-kappa B activation in peritoneal macrophages. Biological & Pharmaceutical Bulletin. 28:233-237.
Kumar P, Paul B, Kumar S, Ali M and Sexana A. (2004).
Correlation of cytokine and mobility in mice with arthritis and during therpy with Swertia chirayita. Journal of Herbal Pharmacotherapy. 4:33-45.
Kwon SH, Lee HK, Kim JA, Hong SI, Kim SY, Jo TH, Park YI, Lee CK, Kim YB, Lee SY and Jang CG. (2011).
Neuroprotective effects of Eucommia ulmoides Oliv. bark on amyloid beta25-35-induced learning and memory impairments in mice. Neuroscience Letters. 487:123-127.
Lee YG and Cho JY. (2007). Inhibitory effects of curcumin on nitric oxide production in lipopolysaccharide-stimulated RAW 264.7 cells and its suppressive mechanism. Korean Journal of Medicinal Crop Science. 15:451-456.
Lie H, Chen B and Yao S. (2005). Application of ultrasonic tech- nique for extracting chlorogenic acid from Eucommia ulmodies Oliv. Ultrasonics Sonochemistry. 12:295-300.
Lin J, Fan YJ, Mehl C, Zhu JJ, Chen H, Jin LY, Xu JH and Wang HM. (2011). Eucommia ulmoides Oliv. antagonizes H2O2-induced rat osteoblastic MC3T3-E1 apoptosis by inhibiting expressions of caspases 3, 6, 7, and 9. Journal of Zhejiang University Science B. 12:47-54.
Lee SE, Lee JH, Kim JK, Kim GS, Kim YO, Choi JH, Lee ES, Noh HJ, Kim SY and Soe JS. (2011). Anti-inflammatory activity of medicinal plant extracts. Korean Journal of Medicinal Crop Science. 19:217-226.
Medzhitov R and Janeway CA Jr. (1997). Innate immunity: the virtues of a nonclonal system of recognition. Cell. 91:295-298.
Morrison DC and Ryan JL. (1987). Endotoxins and disease
mechanisms. Annual Review of Medicine. 38:417-432.
Nishimoto N and Kishimoto T. (2006). Interleukin 6: from bench to bedside. Nature clinical practice. Rheumatology. 2:619-626.
Park HJ, Jung wt, Basnet P, Kadota S and Namba T. (2004).
Syringin 4-O-b-glucoside, a new phenyprosynthase inhibitor, from the stem bark of Magnolia sieboldii. Journal of Natural Products. 59:1128-1130.
Sacco RE, Waters WR, Rudolph KM and Drew ML. (2006).
Comparative nitric oxide production by LPS-stimulated monocyte-derived macrophages from Ovis canadensis and Ovis aries. Comparative Immunology, Microbiology and Infectious Diseases. 29:1-11.
Shin M, Chen YD, Shen CR and Cherng. (2010). A molecular pharmacology study into the anti-inflammatory actions of Eluphorbia hirta L. on the LPS-induced RAW 264.7 cells through selective iNOS protein inhibition. Journal of Natural Medicine. 64:330-335.
Zhu HW, Di HY, Zhang YY, Zhang JW and Chen DF. (2009).
A protein-bound polysaccharide from the stem bark of Eucommia ulmoides and its anti-complementary effect. Carbohydrate Research. 344:1319-1324.