Protective Effect of the Ethyl Acetate-fraction of Methanol Extract of Ophiophogon japonicus on Amyloid beta Peptide-induced Cytotoxicity in PC12 Cells
Ja-Young Moon
1*, Eun-Sook Kim
1, Soo-Jin Choi
1, Jin-Ik Kim
1, Nack-Shik Choi
2, Kyoung Lee
1, Woo-Jin Park
3and Young-Whan Choi
41Department of Bio-Health Science, College of Natural Sciences, Changwon National University, Changwon, Gyungnam 51140, Korea
2KPROTEC INC. Youseong, Daejeon 34141, Korea
3School of Life Sciences, Kwangju Institute of Science and Technology, Kwangju 61005, Korea
4Department of Horticultural Bioscience, College of Natural Resource and Life Sciences, Pusan National University, Miryang, Gyungnam 50463, Korea
Received January 3, 2019 /Revised January 7, 2019 /Accepted February 12, 2019
Amyloid β-protein (Aβ) is the principal component of senile plaques characteristic of Alzheimer’s dis- ease (AD) and elicits a toxic effect on neurons in vitro and in vivo. Many environmental factors, includ- ing antioxidants and proteoglycans, modify Aβ toxicity. It is worthwhile to isolate novel natural com- pounds that could prove therapeutic for patients with AD without causing detrimental side effects.
In this study, we investigated the in vitro neuroprotective effects of the ethyl acetate fraction of meth- anol extract of Ophiophogon japonicas (OJEA fraction). We used an MTT reduction assay to detect pro- tective effects of the OJEA fraction on Aβ
25-35-induced cytotoxicity to PC12 cells. We also used a cell- based β-secretase assay system to investigate the inhibitory effect of the OJEA fraction on β-secretase activity. In addition, we performed an in vitro lipid peroxidation assay to evaluate the protective effect of the OJEA fraction against oxidative stress induced by Aβ
25-35in PC12 cells. The OJEA fraction had strong protective effects against Aβ
25-35-induced cytotoxicity to PC12 cells and was strongly inhibitory to β-secretase activity, which resulted in the attenuation of Aβ generation. In addition, the OJEA frac- tion significantly decreased malondialdehyde (MDA) content, which is induced by the exposure of PC12 cells to Aβ
25-35. Our results suggested that the OJEA fraction contained active compounds ex- hibiting a neuroprotective effect on Aβ toxicity.
Key words : Amyloid beta (Aβ) protein, b-site APP cleaving enzyme (BACE), malondialdehyde,
Ophiophogon japonicus, PC12 cell, β-secretase
*Corresponding author
*Tel : +82-55-213-3552, Fax : +82-55-213-3550
*E-mail : [email protected]
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Life Science 2019 Vol. 29. No. 2. 173~180 DOI : https://doi.org/10.5352/JLS.2019.29.2.173
Introduction
Alzheimer’s disease (AD) is characterized by senile pla- ques composed of amyloid beta (Aβ) peptide and neuro- fibrillary tangles [4, 23]. It has been hypothesized that this plaque is the primary cause of the AD disease [13]. Aβ pep- tide is a major and core component of amyloid plaques and has been reported to be toxic [16]. Aβ peptide is generated following the sequential cleavage of the amyloid precursor protein (APP) by β-and γ-secretase [32]. First, APP is cleaved by β-secretase, also known as β-site APP cleaving enzyme
(BACE), to produce the secreted BACE1-cleaved APP ecto- domain (APPsβ) and the membrane-bound C-terminal frag- ment C99. Next, C99 is cleaved by γ-secretase, which re- leases Aβ. The β-secretase is the key rate-limiting enzyme for the generation of Aβ [6, 21].
The plant Ophiopogon japonicus (Thunb.) Ker-Gawl, widely distributed in Southeast Asia, is known as an important tra- ditional Chinese herbal medicinal plant. Its rhizome is the primary medicinal portion and has been used as a traditional Chinese medicine to treat a wide range of disorders such as cardiovascular and chronic inflammatory diseases, throm- bosis, arrhythmias, respiratory disease, hyperglycemia and immunoregulation for thousands of years [19, 22, 34, 36].
Previous phytochemical studies showed that the rhizome of
O. japonicus contained a wide variety of bioactive compo-
nents including monoterpene glycosides, saponins, poly-
saccharide and homoisoflavonoids [2, 5, 15]. In recent years,
the polysaccharides isolated from the roots of O. japonicus
have drawn the attention of researchers and consumers due
to their nutritional and health protective values as an im- munostimulant, as anti-ischaemetic and hypoglycemic agent, and for their ability to inhibit platelet aggregation [10, 35].
The study of the biological functions of O. japonicus has been limited largely to demonstrate antioxidant activities in vitro. Many studies have also been performed to explore the relationship between the bioactive constituents of O. japoni- cus and their anti-inflammatory properties [2, 5, 15, 19, 22, 34, 36]. However, there is little information regarding the effects of O. japonicus on the neuronal diseases such as Alzheimer’s disease. Moreover, there is no report on the mo- lecular mechanisms underlying anti-amyloid beta activities of O. japonicus. In this report, we investigated the effects of the OJEA fraction against toxicity induced by Aβ in the neu- ronal cells. We also evaluated the inhibitory potency of the fraction to β-secretase activity in the cell-based BACE assay system.
Materials and Methods
Chemicals
Cell culture reagents such as fetal bovine serum (FBS), horse serum, L-glutamine, non-essential amino acid, pen- icillin & streptomycin, 96 well plate, collagen, Phosphate Buffered Saline (PBS), trypsin, Dulbecco's modified Eagle's medium (DMEM) were purchased from Gibco BRL(Grand Island, NY, USA). Aβ
25–35(molecular formula: C
45H
81N
13O
14S, molecular weight: 1,060.27, purity: ³≥97%) was purchased from Sigma-Aldrich (St. Louis, USA). Aβ
25–35was diluted to 1 mg/ml with phosphate buffered saline (PBS). When us- ing, it was diluted to different concentrations with DMEM medium. 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazo- lium bromide (MTT), and dimethyl sulfoxide (DMSO) were purchased from Sigma-Aldrich (St. Louis, USA). All other chemicals were purchased from Sigma-Aldrich (St. Louis, USA) unless otherwise mentioned.
Preparation of the OJEA fraction
Fresh tubers of O. japonicus were purchased from farmers in Miryang (Miryang, Korea) and the species was identified by one of the study authors (Young Whan Choi). The fresh tubers were dried in an incubator at 50℃ for 2 days. The dried tuber of O. japonicus was pulverized using a blender.
The powdered tuber was extracted by maceration using 80%(v/v) methanol (powder: solvent ratio of 1:10 [w/v]) in a sonicator for 1 hr. The pooled extracts were filtered through
a cotton plug followed by Whatman No. 2 filter paper. The filtrate was then concentrated in a rotary evaporator (EYELA N-1200A, Japan). Subsequently, 40 g freeze-dried methanol extract of O. japonicus was suspended in 350 ml distilled wa- ter and sonicated for 30 min. The mixture was then trans- ferred to a 1-L separatory funnel (Schott Duran) and, using solvent-solvent extraction, the methanol extract of O. japoni- cus was sequentially fractionated with hexane (4x500 ml), chloroform (4x500 ml), ethyl acetate (4x500 ml), and n-buta- nol (4x500 ml) to separate the active compounds present in the plant extract. The resulting four fractions were then con- centrated on a rotary evaporator under reduced pressure (-760 mmHg) and the remaining residues were finally freeze-dried to obtain the dry extracts. The fractions were kept in refrigerator until further use. In this study, the ethyl acetate-fraction (OJEA fraction) was used and was dissolved in DMSO before use of cell culture.
PC12 Cell culture
PC12 (rat pheochromocytoma) cells were cultured in Dulbecco's modified Eagle's medium (DMEM) supple- mented with 5%(v/v) FBS, 10%(v/v) horse serum, 100 units/ml penicillin, and 100 μg/ml streptomycin onto the collagen type I (500 mg/10 cmΦ dish, Sigma)-coated culture dish. Cells were grown to confluence at 37℃ in 5% CO
2. The medium was changed two or three times per week. The cells at 80% confluency were maintained by sub-seeding in the same medium.
MTT assay for cell viability
Cell viability was determined colorimetrically by measur-
ing the reduction of the tetrazolium salt, 3-[4,5-dimethylthia-
zol-2-yl]-2,5-diphenyl tetrazolium bromide (MTT) to for-
mazan. Briefly, PC12 cells were seeded at a density of 2x10
4cells/well onto collagen type I-coated 96-well plates (Falcon,
Germany) in Dulbecco's modified Eagle's medium (DMEM)
supplemented with 5%(v/v) fetal bovine serum, 10%(v/v)
horse serum, 100 units/ml penicillin, and 100 μg/ml strepto-
mycin and cultured for 24 hr. After cell attachment, culture
media were freshly changed, and various concentrations of
Ab
25-35or OJEA fraction were added. Cells were additionally
cultured for 24 hr and then MTT solution (10 μl, 5 mg/ml
in PBS) was added to the wells. After 3 hr incubation, the
medium was removed, and 100 ml of DMSO was then added
to dissolve the formazan crystals produced by the cells. The
optical density of formazan solution was measured at 540
nm with a microplate reader (Biotek EL808, USA). Cell via- bility was expressed as a percentage of the value against the non-treated control group.
β-Secretase (BACE) enzyme activity assay β-Secretase activity was measured by using a cell-based BACE assay system that was developed by Oh et al [25].
Briefly, after culturing in the 5 ml SD medium (Leu-, Trp-, 2% glucose), the recombinant yeast S2 cells stably trans- formed with pHS/GMII-β-sites-AP and pMT/BACE were sub-cultured in the 200 ml SD medium until absorbance at 600 nm reached to around 1.0. The sub-cultured recombinant yeast S2 cells were harvested by centrifugation at 3,000 rpm for 10 min at room temperature. After suspension in 1 ml SD medium the cells were mixed with samples with their different concentrations. The yeast recombinant cells mixed with samples were seeded into 96-well plates at a density of 5×10
5cells/well in 150 ml culture medium. The plates were preincubated for 12 hr at 10℃ in the presence or ab- sence of protease inhibitors, or samples and further in- cubated for 1 hr at 37℃ to drive the expression of the GMII- β-sites-AP fusion protein. The cells were centrifuged for 10 min at 1,000 rpm, and then 50 ml of supernatant from each sample was carefully transferred into a new 96-well plate and heated for 10 min at 65℃ to inactivate the endogenous alkaline phosphatase. One hundred ml of 20 mM L-homo- arginine solution containing AP substrate, pNPP (Sigma), were added to each well and incubated for 2-24 hr at room temperature. AP activity was determined by the measure- ment of absorbance at 405nm with a microplate reader (Molecular Devices). The peptidomimetic compounds, #190 and #192, were used as positive control for BACE inhibitor.
Another peptidomimetic compound, #202, which is not per- meable to yeast cell membrane, was used as negative control. Dose-dependent inhibition of BACE was analyzed using SigmaPlot (SPSS Inc.).
Assay for in vitro lipid peroxidation
Lipid peroxidation was determined by measuring the con- tent of thiobarbituric acid reactive substances (TBARS) by the double heating method [7], which was based on the col- orimetric determination of the purple color generated by the reaction between TBARS and malondialdehyde (MDA).
MDA was used as a standard. Briefly, PC12 cells were in- cubated with Aβ
25-35(1 mg/ml) alone with or without OJEA fraction (100 mg/ml) for 24 hr. Culture supernatants in cells
were collected by centrifugation. Supernatants were mixed with trichloroacetic acid (10%, w/v) solutions, followed by 15 min of boiling. Samples were then centrifuged at 3,000 rpm for 10 min, and the supernatant was transferred to new tubes to react with TBA (0.67%, w/v) solution. After boiling for 15 min again, samples were cooled to room temperature, and TBARS concentrations were determined by measuring absorption differences at 532 nm and 520 nm with a micro- plate reader, and by using a molar absorption coefficient of 1.56×10
5M
-1cm
-1.
Statistical analysis
Student’s t-test was used to determine the statistical sig- nificance of the difference between values for the various experimental and control groups. Experimental data were expressed as means ± standard deviation (SD), and the re- sults were obtained from at least three independent experi- ments performed in triplicate. A p-value of 0.05 or less was considered statistically significant.
Results
Cytotoxicity of Aβ
25-35on PC12 cells
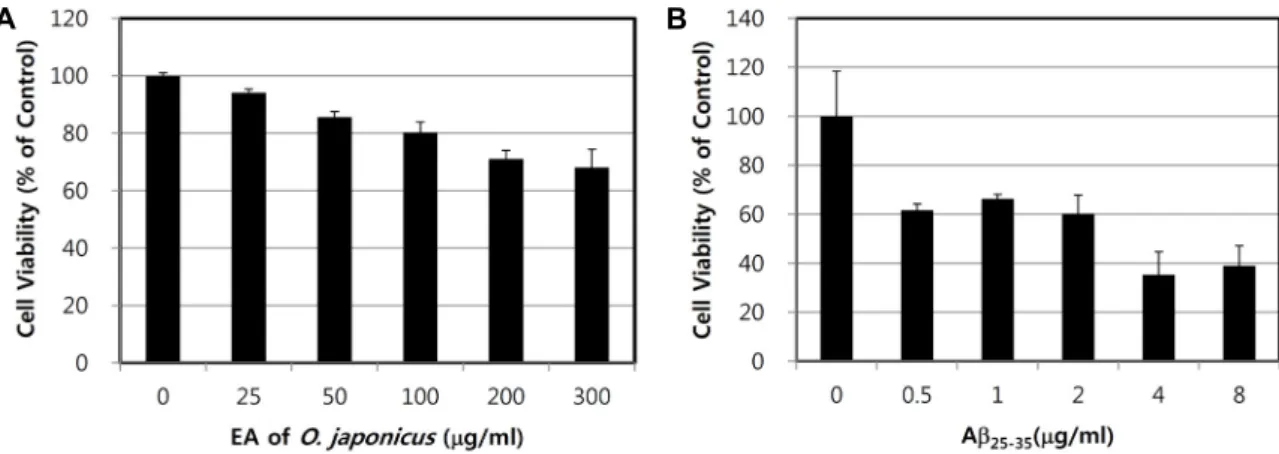
As a first step, in order to identify non-cytotoxic dose ranges of the OJEA fraction for rat PC12 cells, cytotoxicity was observed after exposure to the various concentrations of the OJEA fraction up to 300 μg/ml. When cultured for 24 hr, OJEA fraction itself had no or very low cytotoxic effect on PC12 cells up to 300 μg/ml (Fig. 1A), at which concen- tration cell viability was 70% of vehicle-treated control group.
Based on this result, the concentration of OJEA fraction at 100 μg/ml was applied for lipid peroxidation assay. We also tested the cytotoxicity of Aβ
25-35on PC12 cells by MTT reduc- tion assay. As shown in Fig. 1B, as the concentrations of Aβ
25-35(0.5, 1, 2, 4, and 8 mg/ml) were increased for 24h, the cell viability was decreased in a dose-dependent manner.
This result indicated that Aβ
25-35induced toxicity in PC12 cells. Based on this result, the concentration at 1 μg/ml of Aβ
25-35was applied for other experiments in this study.
Inhibitory effects of the OJEA fraction against cell death induced by Aβ
25-35A preliminary experimental result with the four organic
solvent-fractions, chloroform-, ethyl acetate-, n-hexane- and
butanol-fraction showed that, at concentrations of 25, 50, 100,
200, and 300 μg/ml, the OJEA fraction among the organic
A B
Fig. 1. Effects of the OJEA fraction (A) and Aβ25-35 (B) on viability of PC12 cells. The cells were cultured for 24 hr in DMEM supplemented with 10% FBS, 10% horse serum, 100 units/ml penicillin, and 100 μg/ml streptomycin containing different concentrations of OJEA fraction (A) or Aβ25-35 (B). Cell viability in PC12 cells was determined by MTT reduction assay as described in Materials and Method.
Fig. 2. Protective effects of OJEA fraction on Aβ25-35-induced cytotoxicity in PC12 cells. Data are presented as means
± standard deviation from independent experiments per- formed in triplicate (n=3). *, significantly different from Aβ25-25 only treated group, p<0.05.
solvent fractions possessed the strongest potency to attenu- ate the Aβ
25-35–stimulated cytotoxicity in PC12 cells (Data not shown).
To examine whether OJEA fraction inhibit the neuronal cell death induced by amyloid beta peptide, PC12 cells were simultaneously exposed to 1 μg/ml Aβ
25-35and the OJEA fraction at various concentrations for 24 hr. As shown in Fig. 2, when PC12 cells were cultured for 24 hr after treat- ment of 1 μg/ml Aβ
25-35alone, the cell viability was de- creased to about 60% compared with vehicle treatment.
When PC12 cells were cultured for 24 hr after co-treatment of 1 μg/ml Aβ
25-35with OJEA fraction, all the OJEA fraction applied at the range of the concentrations applied showed a strong potency to attenuate Aβ
25-35–stimulated cytotox-
icity. Especially OJEA fraction at 100 μg/ml significantly and strongly prevented the cell death induced by Aβ
25-35.
In vitro inhibitory activity of the OJEA fraction against BACE
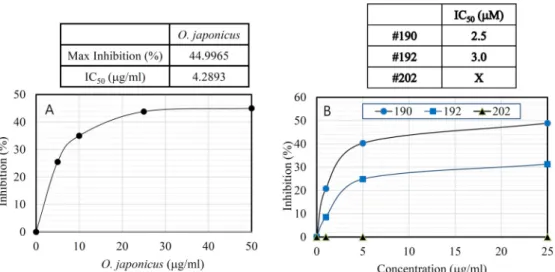
Since AD pathogenesis is potentially attributed to Aβ for- mation, the anti-Alzheimer effects of the OJEA fraction were evaluated by testing the inhibition of BACE (Fig. 3A). Our results showed the OJEA fraction had significant BACE in- hibitory activity in the cell-based BACE enzyme assay (Fig.
3A). The OJEA fraction exhibited BACE-inhibitory activities in a concentration-dependent manner with IC
50value of 4.29 μg/ml. This result indicates that the OJEA fraction may de- crease production of the Aβ through an efficient inhibition of β-secretase activity implicated in Alzheimer's disease.
As shown in Fig. 3B, the peptidomimetic compounds #190 and #192 inhibited BACE in a dose-dependent manner with IC
50values of 2.5 and 3.0 μM, respectively, as determined by in vitro assays using purified BACE and synthetic pep- tides as substrates. However, #202 did not inhibit BACE, probably because this compound is not permeable to cell membrane due to its negative charge (Fig. 3B). Compounds
#190 and #192 are uncharged compounds with slightly dif-
ferent conformations, and compound #202 is a carboxylated
derivative of #192. No cellular toxicity was observed with
the compounds, as judged by trypan blue exclusion assays
(data not shown). The total expression levels of BACE and
its substrate, GMII-β-site (Sw)-AP, were also unaffected by
the compounds (data not shown). Therefore, in fact, it im-
plied that the inhibitory effect of the compounds is due to
direct inhibition of BACE. The compounds inhibited BACE
Fig. 3. Dose-dependent inhibitory potency of the OJEA fraction (A) or three peptidomimetic compounds (B) to BACE. Various amounts of the OJEA fraction were tested for measuring its inhibitory potency. Three peptidomimetic compounds were tested as reference inhibitors for BACE. The assay was performed three times. The curves were fitted using Sigma Plot, yielding IC50 values of the fractions.
Fig. 4. Protective effects of the OJEA fraction against lipid per- oxidation induced by Aβ25-35 treatment in PC12 cells.
Data are presented as means ± standard deviation from independent experiments performed in triplicate (n=3).
*, significantly different from Aβ25-25 treated group, p<
0.05.
in a dose-dependent manner (Fig. 3B).
Effect of the OJEA fraction on lipid peroxidation To observe the protective effect of the OJEA fraction on the oxidative stress induced by Aβ
25-35, MDA production in PC12 cells was measured. As shown in Fig. 4, exposure of PC12 cells to 1 μg/ml of Aβ
25-35for 24 hr resulted in a sig- nificant increase in the level of MDA (6.29±1.00 nmol/mg protein) as compared to the vehicle-treated cells (1.19±0.50 nmol/mg protein, p<0.05). When PC12 cells were co-treated with 1 μg/ml of Aβ
25-35and 100 μg/ml of OJEA fraction, the content of MDA (2.63±1.24 nmol/mg protein, p<0.05)
was significantly decreased as compared to the 1 μg/ml of Aβ
25-35treated cells, as well as compared to the cells treated with 100 μg/ml of OJEA fraction (1.63±0.18 nmol/mg pro- tein, p<0.05). However, OJEA fraction itself did not induce lipid peroxidation. This result suggests that the OJEA frac- tion protects the cytotoxicity induced by Aβ
25-35through its anti-oxidative stress.
Discussion
A key event in AD is the conversion of the β-amyloid
peptide into amyloid plaques in the brain. Preventing Aβ
aggregation is pursued as a therapeutic strategy for treating
AD. Evidence shows that an abnormal deposition of amyloid
beta peptide was the primary cause of the pathogenesis of
AD. It has been shown that Aβ
1-42led to neurotoxicity and
increased protein oxidation [9]. Moreover, it has been pro-
posed that Aβ
25-35represents the biologically active region
of Aβ
1-42. Because it represents the shortest fragment that
exhibits large β-sheet aggregated structures and retains the
toxicity of the full-length peptide [28], Aβ
25-35-induced cyto-
toxicity is recognized as a typical model of Alzheimer’s dis-
ease [31, 33]. The elimination of Aβ is also considered an
important target for the treatment of AD. In agreement with
this, our present results also showed that Aβ
25-35-treatment
induced cytotoxicity in a concentration dependent manner
in PC12 cells. Therefore, we investigated the protective activ-
ity of OJEA fraction against Aβ
25-35-treatment.
Recent studies have focused on APP proteolysis and Aβ generation as potential targets for an AD therapy [12]. In the amyloidogenic process, amyloid precursor protein (APP) is sequentially cleaved by β- and γ-secretases to generate insoluble Aβ. Alternatively, APP is cleaved by α- and γ-sec- retases in a non-amyloidogenic manner, which produces soluble amyloid precursor protein a (sAPPα) [8]. Since Aβ is generated from the APP by BACE and γ-secretases, BACE inhibitors should reduce Aβ levels which may counter the progress of AD pathogenesis. Development of BACE in- hibitors targeting the fibrillary aggregates of Aβ peptide takes, therefore, an active interest as potential therapeutic agents for AD [11, 30]. In this study, we report that the neu- roprotective effect of OJEA fraction on Aβ
25-35-induced cell injury in PC12 cells might be from its potency to inhibit β- secretase activity.
Many studies have indicated that Aβ plaques are accom- panied by neurotoxicity and oxidative stress, which gen- erates free radicals with lipid peroxidation in the brain.
During the oxidative stress by Aβ, MDA, which is the final product of membrane lipid peroxidation, is an important in- dex for determining the level of oxidative stress. In this study, we investigated the protective effect of OJEA fraction against lipid peroxidation induced by Aβ
25-35-treatment in PC12 cells. Our result indicated that Aβ
25-35-treatment in- creased the level of MDA significantly in the cells. In con- trast, OJEA fraction strongly suppressed the generation of MDA induced by Aβ
25-35-treatment. This result suggests that OJEA fraction exerts protective effect of lipid peroxidation induced by Aβ
25-35in the PC12 cells. The reason for using PC12 cell, a nervous-like cell derived from rat adrenal me- dulla pheochromocytoma is that PC12 cell is a widely used cell line for the study of the nervous system in vitro [29].
Further detailed studies are also necessary to confirm its pro- tective effect at a mechanistic level.
Recently, many research groups are trying to isolate novel natural compounds possessing anti-oxidative activity from medicinal plants. Among the medicinal plants, Curcumi- noids isolated from Curcuma longa L [27], extracts of Gingko biloba (Egb761) [3, 20, 26], Huperzia serrata Travis [1], Magnolia officinalis [14] and walnut extract [24] were re- cently reported as the potential therapeutics of AD. It was also reported that berberine and ginseng contain the com- pounds inhibiting toxicity of metabolites of amyloid beta peptide, increase viability of neuronal cells, and delay the aggravation of AD symptom [18]. We previously screened
the protective effects of Liliaceae root extracts on amyloid be- ta protein-induced death in neuronal SK-N-MC and PC12 cells [17]. During the screening, we found that methanol-ex- tracts of Polygonatum falcatum, O. japonicus, Liriope platyphylla, and Hemerocallis fulva strongly attenuated cytotoxicity in- duced by the three Aβ fragments (Aβ
25-35, Aβ
1-42Aβ
1-43). We reported that, among the methanol-extracts of Liliaceae roots prescreened, O. japonicus was especially the most effective in protecting PC12 cells from the Aβ
25-35-induced oxidative cell death. Our present results further indicate that the ethyl acetate fraction of the methanol extract, OJEA, showed the protective effect on Aβ
25-35-induced cell death. The effect might be from its anti-oxidative property as well as its in- hibitory potency to BACE activity. Thus, our results suggest that OJEA fraction contain compound(s) that possess strong protective potencies to the Aβ-induced cytotoxicity by de- creasing at least lipid peroxidation.
Ethyl acetate as a polar aprotic solvent is widely used for extraction of many biological compounds to evaluate their biological activities. So far, numerous bioactive polar and non-polar compounds have been reported from O. japo- nicus showing various pharmacological activities [2, 5, 15].
Based on the literature, some bioactive components such as alkaloids, steroidal saponins and homoisoflavonoids might be dissolved in the OJEA fraction. Although no reported as anti-Alzheimer’s disease activities yet, the compounds may possess multifunctional pharmacological activities, poten- tially valuable for anti-Aβ toxicity based on their anti-oxida- tive activities. The present findings appear useful in leading to further experiments on the identification and character- ization specific compounds that are responsible for the rela- tively strong protective potencies to the Aβ
25-35-induced cyto- toxicity in OJEA fraction.
Acknowledgments
This study was supported by a grant from Changwon National University in 2017~2018. The author thanks Prof Woo Jin Park of Kwangju Institute of Science and Technol- ogy for graciously providing Cell-based BACE assay system.
References
1. Andrea, Z. 2003. The psychopharmacology of huperzine A:
an alkaloid with cognitive enhancing and neuroprotective properties of interest in the treatment of Alzheimer’s disease. Pharmacol. Biochem. Behav. 75, 675-686.
2. Anh, N. T. H., Sung, T., Porzel, A., Franke, K. and Wessjo- hann, L. A. 2003. Homoisoflavonoids from Ophiopogon ja- ponicus Ker-Gawler. Phytochemistry 62, 1153-1158.
3. Bastinetto, S., Ramassamy, C., Dore, S., Christen, Y., Poirier, J. and Quirion, R. 2000. The ginkgo biloba extract (EGb 761) protects hippocampal neurons against cell death induced by β-amyloid. European J. Neurosci. 12, 1882-1890.
4. Braak, H., de Vos, R. A., Jansen, E. N., Bratzke, H. and Braak, E. 1998. Neuropathological hallmarks of Alzheimer's and Parkinson's diseases. Prog. Brain Res. 117, 267-85.
5. Chen, X. M., Jin, J., Tang, J., Wang, Z. F., Wang, J. J., Jin, L. Q. 2011. Extraction, purification, characterization and hy- poglycemic activity of a polysaccharide isolated from the root of Ophiopogon japonicus. Carbohyd. Polym. 83, 749-754.
6. Cole, S. L. and Vassar, R. 2008. The role of amyloid pre- cursor protein processing by BACE1, the beta-secretase, in Alzheimer disease pathophysiology. J. Biol. Chem. 283, 29621- 29625.
7. Draper, H. H. and Hadley, M. 1990. Malondialdehyde deter- mination as index of lipid peroxidation. Methods in Enzymol.
186, 421-431.
8. Durand, D., Carniglia, L., Beauquis, J, Caruso, C., Saravia, F. and Lasaga, M. 2014. Astroglial mGlu3 receptors promote alpha-secretase-mediated amyloid precursor protein cleavage.
Neuropharmacology 79, 180-189.
9. D'Ursi, A. M., Armenante, M. R., Guerrini, R., Salvadori, S., Sorrentino, G. and Picone, D. 2004. Solution structure of amyloid beta-peptide (25-35) in different media. J. Med.
Chem. 47, 4231-4238.
10. Fan, J. and Zhang, X. 2006. Research progress in pharmacol- ogy of Ophiopogon japonicus polysaccharides. Chinese Archives of Traditional Chinese Medicine 24, 626-627.
11. Guo, T. and Hobbs, D. W. 2006. Development of BACE1 inhibitors for Alzheimer's disease. Curr. Med. Chem. 13, 1811-1829.
12. Haass, C. 2004. Take five--BACE and the gamma-secretase quartet conduct Alzheimer's amyloid beta-peptide generation.
EMBO J. 23, 483-488.
13. Hardy, J. and Selkoe, D. J. 2002. The amyloid hypothesis of Alzheimer's disease: progress and problems on the road to therapeutics. Science 297, 353-356.
14. Hoi, C. P, Ho, Y. P., Baum, L. and Chow, A. H. 2010.
Neuroprotective effect of honokiol and magnolol, com- pounds from Magnolia officinalis, on beta-amyloid-induced toxicity in PC12 cells. Phytother. Res. 24, 1538-1542.
15. Hung, T. M., Thu, C. V., Dat, N. T., Ryoo, S. W., Lee, J.
H., Kim, J. C., Na, M., Jung, H. J., Bae, K. and Min, B. S.
2010. Homoisoflavonoid derivatives from the roots of Ophiopogon japonicus and their in vitro anti-inflammation activity. Bioorg. Med. Chem. Lett. 20, 2412-2416.
16. Kihara, T., Shimohama, S., Sawada, H., Honda, K., Nakami- zo, T., Shibasaki, H., Kume, T. and Akaike, A. 2001. Alpha 7 nicotinic receptor transduces signals to phosphatidylinosi- tol 3-kinase to block A beta-amyloid-induced neurotoxicity.
J. Biol. Chem. 276, 13541-13546.
17. Kim, E .S., Choi, S. J., Lee, C. W., Park, K. T. and Moon,
J. Y. 2005. Protective effect of Liliaceae root extracts on amy- loid β-protein-induced death in neuronal cells. Cancer Pre- vention Res. 10, 242-250.
18. Kim, M. H., Kim, S. H. and Yang, W. M. 2014. Mechanisms of action of phytochemicals from medicinal herbs in the treatment of Alzheimerʼs Disease. Planta Med. 80, 1249-1258.
19. Kou, J., Tian, Y., Tang, Y., Yan, J. and Yu, B. 2006. Anti- thrombotic activities of aqueous extract from Radix Ophiopogon japonicus and its two constituents. Biol. Pharm.
Bull. 29, 1267-1270.
20. Le Bars, P. L., Katz, M. M., Berman, N., Itil, T. M., Freedman, A. M. and Schatzberg, A. F. 1997. A placebo-controlled, dou- ble-blind, randomized trial of an extract of Ginko biloba for dementia, North American Egb Study Group. J. Am. Med.
Assoc. 278, 1327-1332.
21. Lin, X., Koelsch, G., Wu, S., Downs, D., Dashti, A. and Tang, J. 2000. Human aspartic protease memapsin 2 cleaves the beta-secretase site of beta-amyloid precursor protein. Proc.
Natl. Acad. Sci. USA. 97, 1456-1460.
22. Li, Y. N., Zhu, D. N., Qi, J., Qin, M. J. and Yu, B. Y. 2010.
Characterization of homoisoflavonoids in different culti- vation regions of Ophiopogon japonicus and related anti- oxidant activity. J. Pharm. Biomed. Anal. 52, 757-762.
23. Masters, C. L., Multhaup, G., Simms, G., Pottgiesser, J., Mar- tins, R. N. and Beyreuther, K. 1985. Neuronal origin of a cerebral amyloid: neurofibriliary tangles of Alzheimer's dis- ease contain the same protein as the amyloid of plaque cores and blood vessels. EMBO J. 4, 2757-2763.
24. Muthaiyah, B., Essa, M. M., Chauhan, V. and Chauhan, A.
2011. Protective effects of walnut extract against amyloid beta peptide-induced cell death and oxidative stress in PC12 cells. J. Neurosci. 36, 2096-2103.
25. Oh, M. S., Kim, S. Y., Oh, Y. S., Choi, D. Y., Sin, H. J., Jung, I. M. and Park, W. J. 2003. Cell-based assay for β-secretase activity. Anal. Biochem. 323, 7-11.
26. Oken, B. S., Storzbach, D. M. and Kaye, J. A. 1998. The effi- cacy of Ginkgo biloba on cognitive function in Alzheimer disease. Arch. Neurol. 55, 1409-1415.
27. Park, S. Y. and Kim, D. S. 2002. J. Nat. Prod. 65, 1227-1231.
28. Pike, C. J., Walencewicz-Wasserman, A. J., Kosmoski, J., Cribbs, D. H., Glabe, C. G. and Cotman, C. W. 1995.
Structure–activity analyses of beta-amyloid peptides: con- tributions of the beta 25-35 region to aggregation and neurotoxicity. J. Neurochem. 64, 253-265.
29. Qian, X., Cao, H., Ma, Q., Wang, Q., He, W., Qin, P., Ji, B., Yuan, K., Yang, F. and Liu, X. 2015. Allopregnanolone attenuates Ab25-35-induced neurotoxicity in PC12 cells by re- ducing oxidative stress. Int. J. Clin. Exp. Med. 8, 13610-13615.
30. Salloway, S., Mintzer, J., Weiner, M. F. and Cummings, J.
L. 2008. Disease-modifying therapies in Alzheimer's disease.
Alzheimers Dement 4, 65-79.
31. Varadarajan, S., Kanski, J., Aksenova, M., Lauderback, C.
and Butterfield, D. A. 2001. Different mechanisms of oxida- tive stress and neurotoxicity for Alzheimer’s A beta (1-42) and A beta (25-35). J. Am. Chem. Soc. 123, 5625-5631.
32. Vassar, R. 2004. BACE1: the beta-secretase enzyme in
초록:소엽맥문동-에틸아세테이트 분획물의 아밀로이드 베타단백질-유발 세포독성에 대한 억제 효능
문자영
1*․김은숙
1․최수진
1․김진익
1․최낙식
2․이 경
1․박우진
3․최영환
4(1창원대학교 생명보건학부, 2케이프로텍, 3광주과학기술원 생명과학부, 4부산대학교 원예생명과학과)
Amyloid β-단백질(Aβ)은 알츠하이머 질병의 특징인 노인성 반점의 주요 성분이며 in vivo와 in vitro에서 신경 세포를 대상으로 독성효과를 유발한다. 항산화물질과 프로테오글리칸을 포함한 많은 환경인자들에는 Aβ의 독성 을 완화하는 물질들이 존재한다. 특히, 천연물질들 중에서 자신은 독성이 없으며, 알츠하이머 환자에게 치료효능 을 나타내는 천연화합물들을 순수 분리하는 것은 매우 가치가 있다. 본 연구에서는 소엽맥문동의 메탄올 추출물 로부터 에틸아세테이트 유기용매로 분획한 물질(OJEA)을 대상으로 in vitro상에서 신경세포독성 제어효능을 탐색
하였다. 본 실험을 위해 PC12 세포주에 Aβ
25-35로 유발한 독성에 대한 OJEA 분획물의 억제효능을 MTT 환원법
분석으로 측정하였으며, β-secretase 활성에 대한 OJEA 분획물의 억제효능을 세포기반 β-secretase assay system
으로 측정하였다. 또한 PC12 세포에서 Aβ
25-35에 의해 유도된 산화적 스트레스에 대한 OJEA 분획물의 억제효과를
지질과산화 분석법으로 수행하였다. 본 연구의 결과는 OJEA 분획물이 PC12 세포에서 Aβ
25-35에 의해 유도된 세포
독성을 강하게 예방 또는 억제하는 효과가 있음을 확인하였으며, 또한 β-secretase의 활성을 억제함으로써 Aβ의
생성을 완화하는 효과를 예상할 수 있었다. OJEA 분획물은 또한 PC12 세포에서 Aβ
25-35에의 노출에 의하여 유도
되는 malondialdehyde (MDA)의 생성을 강하게 억제하였다. 결론적으로, 본 연구의 결과에 의하면 OJEA 분획물 에는 Aβ 독성에 대한 신경세포의 보호효능을 함유하는 생리활성물질이 함유되어 있음을 제시한다.
Alzheimer's disease. J. Mol. Neurosci. 23, 105-114.
33. Yankner, B. A., Duffy, L. K. and Kirschner, D. A. 1990.
Neurotrophic and neurotoxic effects of amyloid β protein:
reversal by tachykinin neuropeptides. Science 250, 279-282.
34. Yu, X. W., Du, H. Z., Sun, L. and Yuan, S. T. 2014. Research progress on the pharmacological effects of Ophiopogonins.
Prog. Pharm. Sci. 38, 279-284.
35. Zheng, Q., Feng, Y., Xu, D. S., Lin, X. and Chen, Y. Z. 2009.
Influence of sulfation on antimyocardial ischemic activity of Ophiopogon japonicus polysaccharide. J. Asian Nat. Prod.
Res. 11, 306-321.
36. Zhou, Y. H., Xu, D. S., Feng, Y., Fang, J. N., Xia, H. L. and Liu, J. 2003. Effects on nutrition blood flow of cardiac mus- cle in mice by different extracts in radix Ophiopogonis. Chin.
J. Exp. Tradit. Med. Form. 9, 22-23.