서 론
어류의 생식기구에 관한 연구는 각 종의 생식소 및 생식세포의 구조, 성 분화와 성 성숙 그리고 정확한 생 식주기와 이를 지배하는 요인 등이 밝혀져야 한다. 이와 관련된 연구는 Cymatogaster aggregata배우자형성 과 정의 내분비조절(Wiebe, 1969), sand goby, Gobius mi- nutus의 생식소 발달과 포란수(Healey, 1971), Horai- chthys setnai의 정소구조 및 정포형성(Grier, 1984), 참 돔, Pagrus major 수컷의 성숙(Matsuura et al., 1987),
Solea lascaris와 S. impar의 성주기, 난자형성과정 및 산란의 비교(Deniel et al., 1989), Ammodytes marinus의 산란과 포란수(Gauld and Hutcheon, 1990), 노래미, Hexagrammos agrammus의 생식주기에 따른 정소소낭 및 간질세포의 구조∙조직화학적 변화(Chung and Lee, 1994), 잉어과 어류 수정관의 미세구조(Lahnsteiner et al., 1994), Anguilla anguilla의 성 분화 및 호르몬의 영 향(Colombo and Grandi, 1995), Atherina boyeri 암컷의 생식생물학적 연구(Tomasini et al., 1996) 그리고 불볼 락, Sebastes thompsoni의 정자 미세구조(Lee et al., 1997b)에 관한 보고 등 아주 많은 편이다.
─
─ 192 ──
주둥치Leiognathus nuchalis의 생식생물학적 연구
이 정 식∙허 성 회*
여수대학교 어병학과, *부경대학교 해양학과
Reproductive Biology of the Slimy, Leiognathus nuchalis (Teleostei: Leiognathidae)
Jung Sick Lee and Sung-Hoi Huh*
Department of Fish Pathology, Yosu National University, Yosu 550-749, Korea
*Department of Oceanography, Pukyong National University, Pusan 608-737, Korea
Reproductive biology of the slimy, Leiognathus nuchalis was investigated by means of histological methods. Sex ratio was 1.81 : 1 in female to male. Developmental pattern of oocytes was group-synchronous. Testicular structure was restricted spermatogonial testis-type of tubular testis. The size of f irst group maturity is 7.5 cm in total length. Gonadosomatic index (GSI) of female was the highest in July (12.83) and the lowest in September (1.08). GSI of male was the highest in June (19.0) and the lowest in October (0.24). Hepatosomatic index (HSI) of female showed to be positively correlated with GSI. Thoracic spot index (TSI) showed to be the minimum value from May to July when the maturation and ripe season of gonad. Reproductive cycle of female could be classif ied into the growing (March~May), maturation (May~June), ripe and spent (June~August), recovery (August~November), and resting stage (November
~March). Reproductive cycle of male could be classified into the multiplicative and growing (January~April), maturation (April~May), ripe and spent (June~August), recovery (August~
October), and resting stage (October~December).
Key words : Leiognathus nuchalis, f irst group maturity, TSI, gonad, reproductive cycle
주둥치는 농어 , 주둥치科에 속하는 경골어류로서 우 리 나라 남부와 일본 중남부의 수심이 얕은 내만에 주 로 서식하며(Chyung, 1977), 전장 10 cm 내외의 소형 어종으로 외부 형태적으로 암수의 구분은 어렵다.
주둥치의 생식에 관해서는 일본 연해에서 채 집된 개체들의 생식소중량지수와 생식소 외부형태의 변 화에 관한 보고가 있으나(� 과 , 1965), 우리 나 라 남해안 광양만 인근해역의 우점종인 주둥치의 생식 과 관련된 국내 연구는 전무한 상태이다.
본 연구는 주둥치의 군 성숙도, 생식소중량지수, 간중 량지수 및 가슴반점지수의 월 변화, 생식세포의 발달, 생 식주기 및 이에 따른 생식소 구조의 변화 등을 조사하 여 이들 어류의 생식생태에 관한 기초자료를 제공하기 위하여 수행되었다.
재료 및 방법
본 연구에 사용된 재료는 1998년 11월부터 1999년 10월까지 매월 광양만 인근 해역(34�52′~34�57′N, 127�46′~127�53′W)에서 채집한 암컷 379, 수컷 209개 체이다(Table 1). 채집된 재료는 전장 및 체중을 각각 0.1 mm, 0.1 g까지 측정하였다. 그 후 생식소 무게를 0.01 g 까지 측정한 다음, 생식소를 Bouin’s f luid에 고정 하여 파라핀 절편법에 의해 4~6µm 두께로 연속 절편 하여 조직표본을 만들었다. 제작된 조직표본은 Mayer’s hematoxylin과 0.5% eosin의 비교염색을 실시하였다. 생 식소중량지수 (gonadosomatic index: GSI)는 성어를 대 상으로 “생식소 무게×100/전중-생식소 및 내장 무게” 의 식으로 구하였다. 간중량지수(hepatosomatic index:
HSI)는 성어를 대상으로 “간 무게×100/전중-생식소
및 내장 무게”의 식으로 구하였다. 가슴반점지수(thora- cic spot index: TSI)는 가슴반점의 진한 정도를 세 단계 (1 =없음, 2 =미약, 3 =명확)로 구분하여 각각의 값을 월 평균으로 나타냈다. 난모세포의 발달단계는 Elorduy- Garay and Pamirez-Luna (1994)의 방법에 따랐다.
결 과
1. 성 비
채집된 개체들의 암∙수 성비는 시기별로 다소 차이 가 있었으나, 전체 평균에서는 1.81 : 1로 암컷의 비율이 높게 나타났다(Table 1).
2. 생식소의 형태 및 내부구조
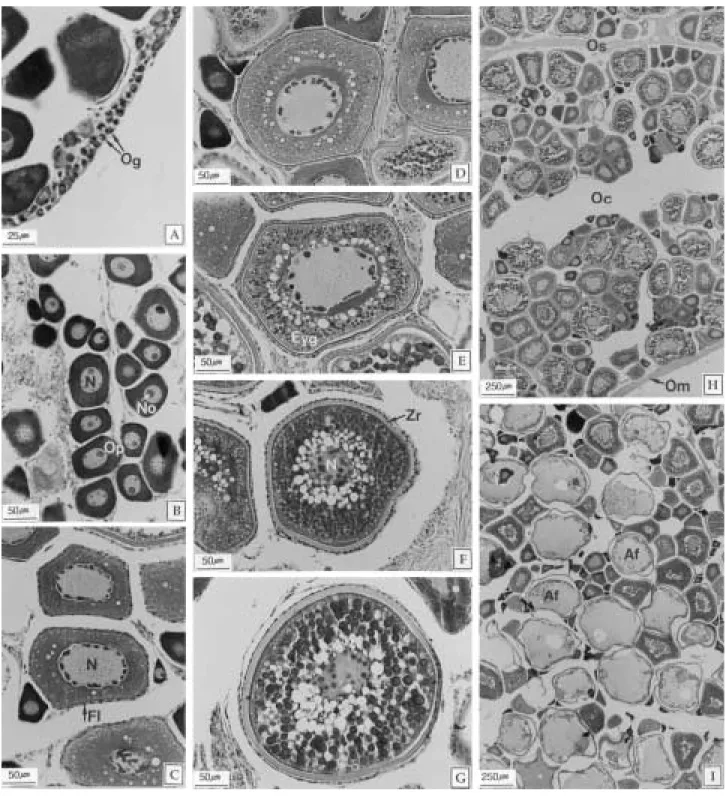
난소는 한 쌍의 둥근 삼각형의 낭상형으로 복강의 등 쪽 후방에 위치한다. 난소내강은 종축으로 형성된 두꺼 운 내부 격벽에 의해 두 부분으로 나누어지고 난소외막 에서 시작된 결체성조직의 난소 소엽으로 채워져 있으 며, 이곳에서 난원세포들이 유래한다. 성숙기의 난소조직 상으로 볼 때 난모세포의 발달양식은 난군동기발달형으 로 관찰되었다(Fig. 7-H).
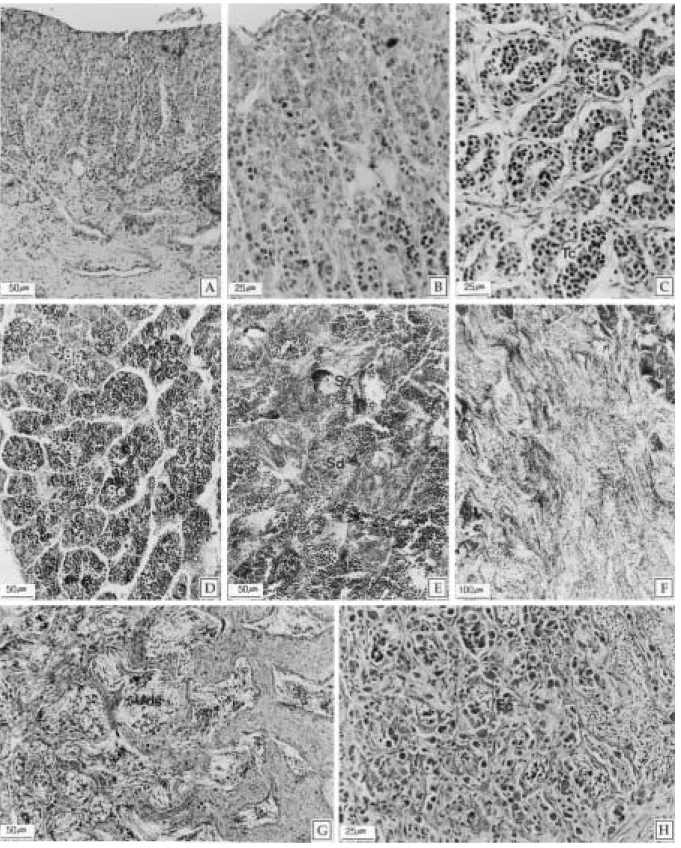
정소는 한 쌍으로 외부형태와 위치는 난소와 유사하 나 성숙하면서 난소보다 다소 크며, 난소는 외형적으로 과립상의 난모세포가 관찰되나 정소는 그렇지 않다. 정 소의 내부 조직상에서는 정세관 구조가 관찰되는데, 이 들 정세관의 형태는 생식세포의 성장기에 가장 뚜렷하 다(Fig. 8-C). 정세관은 다수의 소낭(cyst)을 가지며 각 소낭내의 생식세포들은 같은 발달 단계를 보인다. 정원 세포들은 주로 정소외막 부근의 피질부에 존재하고, 완 숙정자들의 대부분은 수정관 부위에서 관찰되나, 일부는 정소의 피질부와 수질부 사이에서도 관찰된다.
Table 1. Number of specimens and collection date of the slimy, Leiognathus nuchalis for analysis Collection Total number Number Number Sex ratio date of specimens of female of male (F : M)
Nov. 21, 1998 35 25 10 2.50 : 1
Dec. 19, 1998 50 23 27 0.85 : 1
Jan. 11, 1999 66 41 25 1.64 : 1
Feb. 22, 1999 36 22 14 1.57 : 1
Mar. 23, 1999 40 27 13 2.08 : 1
Apr. 24, 1999 41 31 10 3.10 : 1
May 23, 1999 40 26 14 1.86 : 1
June 26, 1999 41 28 13 2.15 : 1
July 16, 1999 80 54 26 2.08 : 1
Aug. 14, 1999 59 46 13 3.54 : 1
Sep. 19, 1999 51 31 20 1.55 : 1
Oct. 18, 1999 49 25 24 1.04 : 1
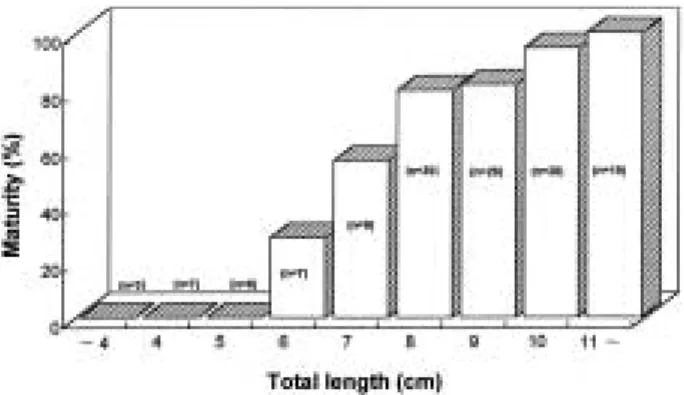
Total / Mean 588 379 209 1.81 : 1 Fig. 1. Maturity with body size of the female slimy, Leiog- nathus nuchalis.
3. 군 성숙도
4월부터 8월사이에 암컷 156개체와 수컷 66개체를 대상으로 생식소의 조직표본을 분석한 결과, 주둥치 암 컷(Fig. 1)과 수컷(Fig. 2) 모두 첫 번째 군 성숙을 나타 내는 크기는 전장 7.5 cm였다.
4. 생식소중량지수(GSI)의 월 변화
1998년 11월부터 1999년 10월까지 조사된 GSI의 월 변화는 Fig. 3과 같다. GSI는 4월부터 큰 폭으로 증가하 여 수컷은 6월에19.0, 암컷은 7월에 12.83으로 연중 가 장 높은 값을 나타냈다. 그후 감소하여 암컷은 9월에 1.08, 수컷은 10월에 0.24로 연중 가장 낮은 값을 나타 냈다.
5. 간중량지수(HSI)의 월 변화
Fig. 4에 나타낸 것과 같이 암컷의 HSI는 2월에 1.60 의 가장 낮은 값을 보인 후 3월부터 증가하기 시작하여
7월에는 3.48로 연중 가장 높은 값을 나타냈으며, 그후 차츰 감소하여 GSI와 유사한 변화양상을 나타냈다.
하지만 수컷의 경우에는 6월에 가장 낮은 값을 보였 으며, 8월과 9월에 2.33으로 가장 높은 값을 보여 GSI 와는 반대되는 변화양상을 나타냈다.
6. 가슴반점지수(TSI)의 월 변화
Fig. 5에서 볼 수 있듯이 주둥치는 암∙수 모두 가슴
지느러미 아래쪽에 노란색의 반점을 가지는데, 이들 가 슴반점의 진한 정도는 시기적으로 차이를 보여 5월부터 7월 사이에는 1.2 내외의 낮은 값을 보이고, 8월부터 이 듬해4월까지는 2.3 내외의 높은 값을 보였다(Fig. 6).
7. 난모세포의 발달단계 1) 난원세포기(Oogonial stage)
난소소엽 내부의 결체성조직에서 주로 관찰되는 난원 세포들의 크기는 직경 5~15µm이다. 핵은 세포의 대부 분을 차지하며, 세포질은 강한 호염기성을 나타냈다(Fig.
7-A).
2) 난황형성전기(Previtellogenic stage)
난모세포들의 크기는 직경 30~70µm이다. 핵이 차지 하는 비율은 세포의 약 50%로서 전 단계에 비해 감소 한 상태이며, 핵내에서는 커다란 한 개의 인과 여러 개의 작은 인들을 관찰할 수 있었다. 세포질은 균질화 되어있 는 상태이며, 여전히 호염기성을 나타냈다(Fig. 7-B).
3) 난황형성 개시기(Initial vitellogenic stage) 이 단계의 난모세포들의 크기는 직경 80~200µm이 다. 핵막을 따라 다수의 불규칙한 형태의 인이 관찰되었 으며, 여포세포층이 구별되었다. 세포질은 호염기성이 점 차 소실되며, 세포질에는 미세한 과립들과 소형의 공포 Fig. 4. Monthly change of hepatosomatic index (HSI) of
the slimy, Leiognathus nuchalis.
Fig. 2. Maturity with body size of the male slimy, Leiog- nathus nuchalis.
Fig. 3. Monthly change of gonadosomatic index (GSI) of the slimy, Leiognathus nuchalis.
상 난황구들이 일부 관찰되었다(Fig. 7-C).
4) 난황형성 활성기(Active vitellogenic stage) 난모세포들의 크기는 직경 200~300µm이며, 여포세 포층의 발달과 방사대의 구분이 가능하다. 이 시기는 두 단계로 나눌 수 있다. 초기에는 세포질의 대부분을 소형 의 공포상 난황구들이 차지하며(Fig. 7-D), 후기에는 공 포상 난황구의 수는 감소하나 크기는 증가된 상태이며, 세포질에서는 eosin에 진하게 염색되는 호산성 난황구 들이 관찰되었다(Fig. 7-E).
5) 성숙기(Mature stage)
성숙 난모세포들의 직경은 300µm 내외이다. 핵은 불 규칙한 형태이며, 잘 발달된 여포세포층과 방사대가 관 찰되었다. 핵 주위의 수질부 세포질에는 공포상의 난황 구가 집중되어 있으며, 피질부 세포질의 대부분은 호산 성 난황구들이 차지하고 있었다(Fig. 7-F).
6) 산란전기(Prehydrated stage)
산란전의 완숙 난모세포는 직경 350µm 내외이며, 잘 발달된 방사대를 가지고 있었다. 세포질에는 공포상의 난황구와 10~15µm 크기의 호산성 난황구들이 혼재하 고 있었다(Fig. 7-G).
8. 암컷의 생식주기
생식주기는GSI와 난소조직상의 월 변화(Fig. 9)를 기 초로 다음과 같이 연속적인5단계로 구분할 수 있었다.
1) 성장기(Growing stage)
이 단계는 3월부터 5월 사이로 이때의 GSI는 2.10~ 3.76이었다. 이 단계에 속하는 개체들의 난소 조직상에 서는 주로 난황형성 개시기(III)의 난모세포들과 일부 난황형성전 단계(II) 및 난황형성 활성단계(IV)의 난모 세포들이 관찰되었다.
2) 성숙기(Maturation stage)
5월부터 6월 사이로 GSI는 3.76~11.07이었다. 이 시 기에는 주로 성숙단계의 난모세포(V)들과 난황형성 활 성단계의 난모세포(IV)들을 가진 개체들이 관찰되었다.
3) 완숙 및 산란기(Ripe and spent stage)
이 단계는6월부터8월 사이로 이때의GSI는 11.07~ 3.80로 연중 가장 높은 값을 나타낸 후 급격히 감소하 는 시기로서 주 산란기는 7월말부터 8월초로 조사되었 다. 난소조직상에서는 산란전 단계의 난모세포(VI)들과 산란 후 폐쇄 여포들이 관찰되었다.
4) 회복기(Recovery stage)
8월부터 11월 사이로 GSI는 3.80~1.25이다. 이 시기 의 난소조직상에서는 주로 폐쇄 여포와 난황형성 개시 기 이후 난모세포들의 퇴화∙흡수가 관찰되었다(Fig.
7-I).
5) 휴지기(Resting stage)
11월부터 3월 사이로 GSI는 1.25~2.10로 조사되었 다. 휴지기 개체들의 난소조직 표본에서는 난원세포(I) 와 난황형성전 단계의 난모세포(II)들이 주로 관찰되었 다.
9. 수컷의 생식주기 및 정소조직상의 변화
주둥치 수컷의 생식주기는 GSI와 정소조직상의 월 변 화(Fig. 10)를 기초로 다음과 같이 구분할 수 있었다.
1) 분열증식 및 성장기(Multiplicative and growing stage)
이 단계는 1월부터 4월 사이로 이때의 GSI는 0.53~ 1.31로 낮은 편이었다. 분열증식기에 해당하는 개체들의 Fig. 5. External morphology and thoracic spot (TS) of the
slimy, Leiognathus nuchalis.
Fig. 6. Monthly change of thoracic spot index (TSI) of the slimy, Leiognathus nuchalis.
Fig. 7. Oocyte developmental stage of the slimy, Leiognathus nuchalis. A: Oogonial stage (I). B: Previtellogenic stage (II).
Note the homogeneous cytoplasm. C: Initial vitellogenic stage (III). Note the small yolk vesicle and follicle layer. D:
Early active vitellogenic stage (IVa). E: Section of lately active vitellogenic stage (IVb) showing the eosinophilic yolk globules. F: Mature stage (V). Note the germinal vesicle breakdown and large eosinophilic yolk globules. G:
Prehydrated stage (VI). H: Section of ripe stage showing the group-synchronous development of oocytes. I: Section of after spawning showing the degenerating oocytes and atretic follicles. Af: atretic follicle, Eyg: eosinophilic yolk globules, Fl: follicle layer, N: nucleus, No: nucleolus, Oc: ovarian cavity, Og: oogonia, Om: outer ovarian membrane, Op: ooplasm, Os: ovarian septum, Zr: zona radiata.
Fig. 8. Testicular developmental stage of the slimy, Leiognathus nuchalis. A: Section of resting stage showing the spermatogonia of interphase. B: Multiplicative stage. C: Initial growing stage. Note the seminiferous tubules, testicular cyst and spermatocytes. D: Active growing stage. E: Mature stage. Note the numerous spermatids. F:
Section of ripe stage showing the basophilic spermatozoa. G: Spent stage. Note the undischarged spermatozoa and abundant interstitial tissue. H: Recovery stage. Note the numerous eosinophilic cells. Ec: eosinophilic cells, Sc:
spermatocytes, Sd: spermatids, Sg: spermatogonia, St: seminiferous tubule, Sz: spermatozoa, Tc: testicular cyst.
Uds: undischarged spermatozoa.
정소 조직상에서는 인이 뚜렷한 간기의 정원세포들과 이형염색질이 관찰되는 분열증식중의 정원세포들이 주 로 관찰되었다(Fig. 8-B). 성장기 가운데 초기 성장기의 조직상에서는 정세관이 뚜렷하고, 정소소낭이 구별되며, 주로 정모세포들이 관찰되었다(Fig. 8-C). 그리고 성장 활성기의 정소 표본에서 정세관의 형태는 관찰이 어려 웠으나 정소소낭은 쉽게 구별되었으며, 주로 정모세포와 일부 정세포들이 관찰되었다(Fig. 8-D).
2) 성숙기(Maturation stage)
4월부터 5월 사이로GSI는 1.31~5.97로 차츰 증가하 는 시기이다. 이 시기의 정소조직상에서는 주로 정모세 포들과 염기성 색소인 헤마톡실린에 진하게 염색되는 일부 정자들이 관찰되었다(Fig. 8-E).
3) 완숙 및 방정기(Ripe and spent stage)
이 단계는 6월부터 8월 사이로 이때의 GSI는 19.0~ 3.45로 연중 가장 높은 값을 나타낸 후 급격히 감소하
는 시기이다. 완숙기 개체들의 정소조직상에서는 주로 염기성 색소에 진하게 염색된 정자들과 일부 정세포 및 정모세포들이 관찰되었다(Fig. 8-F). 방정기 개체들의 정소조직상에서는 정세관 내강에서 잔존 정자들이 관찰 되었으며, 간충조직은 완숙기에 비하여 매우 증가된 상 태였다(Fig. 8-G).
4) 회복기(Recovery stage)
8월부터 10월 사이로 GSI는 3.45~0.24로 급격히 감 소하는 시기이다. 이 시기에는 잔존 정자들의 퇴화∙흡 수와 직경 약 10µm 크기의 세포질이 균질화 되어있는 호산성 세포들이 증가된 조직상(Fig. 8-H)을 보이는 개 체들이 많은 비율을 차지하였다.
5) 휴지기(Resting stage)
10월부터 12월 사이로 GSI는 0.24~0.41로 연중 가 장 낮은 값을 보이는 시기이다. 휴지기의 개체들은 주로 간기의 정원세포들과 간충조직이 풍부한 정소조직상을 보였다(Fig. 8-A).
고 찰
어류 생식소의 형태 및 내부 구조적 차이는 각 어류 의 계통학적 위치 및 이들의 생식생태에 따른 특징이라 할 수 있다.
주둥치 난소는 낭상형이며, 내부구조는 Encrasicho- lina heteroloba (Wright, 1992)와 Caulolatilus princeps (Elorduy-Garay and Pamirez-Luna, 1994)를 비롯한 대부분의 경골어류에서 볼 수 있는 것처럼 결체성 조직 인 다수의 난소소엽으로 구성되며, 이곳에서 난원세포들 이 유래된다.
Grier (1981)는 대부분 어류의 정소는 고등 척추동물 에서 볼 수 있는tubule type과 유사하다고 하였으며, 어 류의 정소형태를 내부구조, 정원세포의 분포 및 생식세 포의 발달 양식에 따라 정원세포 비국재형(unrestrict- ed spermatogonial testis-type)과 정원세포 국재형 (restricted spermatogonial testis-type)으로 구분하였다. 이 가운데 전자는 대부분의 경골어류가 가지는 전형적 인 형태로 정원세포는 정소 전체에 분포하며, 후자의 경 우에 정원세포들은 주로 정소의 피막 근처에 분포하고, 하나의 정소 소낭(cyst)내에서 생식세포는 동시에 발달 하며, 방정시기에 정자들은 수정관 부위로 모이는 형태 이다.
Billard et al. (1982)은 어류의 정소형태를 크게 세관형 (tubular type)과 소엽형(lobular type)으로 구분하였다. 세관형은 내강을 가지지 않으며, 소낭은 정자형성과정 Fig. 9. Monthly change in frequency of ovarian devel-
opment stage of the slimy, Leiognathus nuchalis.
Fig. 10. Monthly change in frequency of testicular devel- opment stage of the slimy, Leiognathus nuchalis.
동안 수정관 쪽으로 이동하는 형태이며, 소엽형은 완숙 된 정자들이 모여지는 내강을 가지는 형태이다. 그러나 여기서 언급하고 있는 세관형은 주로 정포(spermato- zeumata 또는 spermatophore)를 형성하는 체내수정형 어류에서 볼 수 있는 구조이므로 어류 정소의 구조를 세관형과 소엽형 두 가지로 나누는데는 추후 또 다른 기준들이 추가되어야 할 것으로 본다.
본 연구에서 관찰된 주둥치의 정소는 뚜렷한 관 구조 를 가지며, 정원세포들은 주로 정소외막 부근의 피질부 에 존재하고, 정소 소낭내의 생식세포들은 같은 발달 단 계에 속하며, 일부 완숙정자들이 여러 부위에서 관찰되 나 대부분은 수정관 부위에서 관찰된다는 점에서 Grier (1981)의 기준과 비교해 볼 때, 세관형 가운데 정원세포 국재형에 가깝다고 할 수 있다.
일반적으로 온대지역에 서식하는 어류의 계절에 따른 산란형태는 춘계산란형(spring spawner), 춘하계산란형 (spring to summer spawner), 추계산란형(autumn spaw- ner) 및 동계산란형(winter spawner)으로 구분된다(Sh- imizu and Hanyu, 1982; 1983).
본 연구에서 주둥치의 주 산란기는 7월말부터 8월초 로 조사되었으므로 계절에 따른 산란형태는 춘하계산란 형에 속한다.
춘하계산란형 어류 가운데 Solea lascaris와 S. impar (Deniel et al., 1989), Atherina boyeri (Tomasini et al., 1996) 그리고 참서대, Cynoglossus joineri (Lee et al.,
2000) 등에서 난소성숙은 비교적 짧은 기간에 이루어지
는데, 주둥치의 난소성숙도 이와 같은 특징을 보였다. 어류의 산란 및 방정시기를 추정할 수 있는 자료 중 의 하나로 생식소중량지수(GSI)가 사용되고 있는데, 이 미 보고된 그물코쥐치, Rudarius ercodes (Lee and Hanyu, 1984), 점망둑, Chasmichthya dolichignathus (Baek and Lee, 1985), pike, Esox lucius (Treasurer, 1990), Siganus sutor (Ntiba and Jaccarini, 1990), 짱둥 어, Boleophthalmus pectinirostris (Chung et al., 1991), Salmo trutta (Maisse et al., 1991) 그리고 쭈굴감펭, Scorpaena miostoma (Lee et al., 1997a)의 GSI는 수컷에 비해 암컷이 적게는 0.2배에서 많게는 20배 이상 높은 것으로 보고되고 있다.
하지만 본 연구에서 5월에서 7월 사이에는 주둥치 수 컷의GSI가 암컷의 GSI보다 약50 % 이상 높은 것으로 조사되어 � ∙ (1965)의 보고와 유사한 결과를 보였는데, 이와 같은 결과에 대해서는 추후 세부적인 연 구가 필요하리라 생각된다.
생식소중량지수(GSI)와 연관되어 밀접하게 변화하는 간중량지수(HSI)는 주둥치 암컷의 경우에는 GSI와 동
일한 양상으로 변화하였는데, 이러한 경향은 병어, Pampus argenteus와 덕대, P. echinogaster (Lee and Jin, 1989), 노래미, Hexagrammos agrammus (Chung and Kim, 1994), 망상어, Ditrema temmincki (Lee et al., 1995) 에서 볼 수 있다. 그러나 뱀장어, Anguilla japonica (Su- gimoto et al., 1976), 그물코쥐치, Rudarius ercodes (Lee and Hanyu, 1984) 및 문치가자미, Limanda yokohamae (Lee et al., 1985) 등에서는 HSI가 GSI와 역상관으로 변 화한다. 이처럼 변화 양상이 서로 다른 것은 난모세포에 축적되는 난황전구물질의 합성, 저장 및 이용시기가 서 로 다르며, 또한 성 성숙에 필요한 에너지의 소비가 어 종에 따라 서로 다르기 때문이다.
어류의 성숙 및 산란기를 추정하는 방법에는 GSI, 생 식소의 조직학적 관찰 등 직접적인 방법과 체색의 변화 및 이차성징의 발현 등을 이용한 간접적인 방법들이 있 다. 외형적인 특징을 수치화 하여 생식시기를 파악할 수 있는 자료는 각시붕어, Rhodeus uyekii에서 산란관의 길 이변화를 이용하여 성숙 및 산란시기를 파악할 수 있는 보고(An, 1995)와 망상어, Ditrema temmincki 수컷에서 이차성징의 일종인 뒷지느러미 연조 길이 변화에 의해 서 성숙 및 교미시기를 파악한 보고가 있다(Lee et al., 1996).
본 연구에서 주둥치는 가슴지느러미 아래쪽에 노란색 의 반점을 가지며 이들 반점의 진한 정도는 시기적으로 차이를 보이는데, 이를 수치화하여 가슴반점지수(TSI) 로 표시하고 TSI의 월 변화를 본 결과 5월에서 7월 사 이에는 다른 시기에 비해 약 1/2로 낮았다. 따라서 주둥 치에서 가슴반점의 소실은 성숙 및 완숙시기임을 알 수 있는 간접적인 자료로 이용될 수 있을 것이다.
경골어류의 난모세포의 발달양식은 종에 따라 다소 차이가 있는데, Wallace and Selmen (1981)은 이를 동기 발달형(synchronous), 난군동기발달형(group-synchron- ous), 비동기발달형(asynchronous)의 세 가지로 구분하 였다.
주둥치의 난모세포 발달양식은 6월의 성숙단계 난소 내에서 난원세포, 난황형성전 난모세포, 난황형성 개시기 난모세포, 난황형성 활성기 난모세포 및 성숙난모세포들 이 관찰되지만, 이 가운데 난황형성 활성기의 난모세포 와 성숙난모세포들이 많은 수를 차지하므로 Encrasi- cholina heteroloba (Wright, 1992) 등의 대부분 경골어류 에서 볼 수 있는 난군동기발달형에 속하는 것으로 판단 된다.
수서동물의 난모세포 발달과정에서 세포질 내 난황 축적과 관련 있는 구조로는 난병(egg stalk), 방사대 (zona radiata), 난황핵(yolk nucleus) 및 인(nucleolus)
등이 있다. 이 가운데 난병과 방사대는 난모세포의 외재 적 난황축적(exogenous vitellogenesis)과 밀접한 관계가 있으며, 난황핵과 인은 난모세포의 내재적 난황축적 (endogenous vitellogenesis)과 밀접한 관계가 있다 (Beams and Sekhon, 1966; Wallace and Selmen, 1981).
주둥치의 난모세포 발달과정에서 난병과 난황핵은 나 타나지 않았으나, 여포세포층 및 방사대의 발달 그리고 인의 활성화가 관찰되었으므로 주둥치 난모세포의 난황 축적은 대부분 경골어류와 마찬가지로 외재적 난황축적 과 내재적 난황축적 두 가지로 생각된다.
어류의 산란 후 잔존 난모세포의 퇴화∙흡수는 크게 4단계의 과정을 거치는데 각 각의 단계는 다음과 같다. 첫 번째는 난모세포의 파괴 개시 및 발달된 여포세포의 식세포 작용에 의한 난황의 흡수, 두 번째는 여포세포의 빠른 무사분열상의 증가 및 여포강의 침입, 난황의 계속 적인 흡수, 협막세포의 성장과 혈관의 확장, 세 번째는 오렌지색 색소의 출현과 퇴화작용, 네 번째는 난모세포 관련 구조들의 불규칙적 붕괴이다(Saidapur, 1978).
하지만 이러한 과정은 어종에 따라 각 단계의 소요시 간은 다소 차이가 있어서 광학현미경에서 주로 관찰되 는 단계는 다른 것 같다. 그 예로 참서대의 경우에는 위 에서 설명한 전 과정이 관찰되지만 이 가운데 네 번째 단계에 해당하는 조직상이 주로 관찰되었으며(Lee et al., 2000), 쭈굴감펭, Scorpaena miostoma (Lee et al.,
1997a)과 본 연구에서 주둥치의 경우에는 주로 세 번째
단계에 해당하는 조직상이 주로 관찰되었다.
잔존 난모세포의 운명은 난모세포의 발달정도에 따라 다소 차이가 있는데, 고등 척추동물과 메기류에서는 난 황형성전 난모세포(previtellogenic oocyte) 가운데 스테 로이드 합성이 시작된 경우에는 퇴화∙흡수가 확인되었 다(Saidapur, 1978). 하지만 보통 광학현미경적인 관점 에서 난황형성전 난모세포는 퇴화∙흡수되지 않고 다음 의 난소발달에 참여할 것으로 추측되어 왔는데, 이러한 추측은 광학현미경적 연구의 한계에서 온 것으로 생각 된다. 따라서 추후 난황형성전 난모세포의 퇴화∙흡수에 관해서는 미세구조 및 스테로이드 합성 여부를 확인하 는 연구가 함께 수행되어야 할 것이다.
어류 수컷의 생식주기 구분은 연구자 사이에 다소 차 이는 있으나, 보통 분열증식기와 성장기를 구분하고 있 는데, 이는 생식세포의 세포분열상의 차이 및 시기적으 로 이러한 조직상을 가지는 개체들의 출현빈도가 명확 히 구분되기 때문인 것으로 판단된다.
하지만 본 연구에서 분열증식기와 성장기를 같은 범 주에 포함시킨 것은 1월에서 4월 사이에는 분열증식을 나타내는 조직상을 가진 개체들의 출현빈도와 성장기의
조직상을 보이는 개체들의 출현빈도가 유사하기 때문이 었다.
일반적으로 회복기의 생식소에서 잔존 생식세포의 퇴 화∙흡수에 관한 연구는 주로 난소 조직을 대상으로 연 구되어 왔으며, 이러한 과정은 여포세포와 혈구 등의 식 세포 작용에 의해 수행되는 것으로 알려져 있으나 (Saidapur, 1978), 정소 조직에서 이러한 과정에 관한 연 구는 찾아보기 힘들다.
본 연구에서는 회복기 정소 조직상에서는 직경 약 10 µm 크기의 세포질이 균질한 호산성 세포들이 다수 관 찰되었는데, 이러한 세포들은 세포질이 매우 활성화되어 있는 것을 나타내는 특징 가운데 하나인 강한 호산성의 염색성을 보였다. 따라서 이들 세포의 출현시기와 활성 화라는 점에서 잔존 생식세포의 퇴화∙흡수와 관련이 있지 않나 의심해 볼 여지가 있으나 구체적인 연관성에 대해서는 추후의 연구가 필요하리라 본다.
적 요
주둥치의 암∙수 성비는 1.81 : 1로 암컷의 비율이 높 게 나타났다. 난모세포 발달양식은 난군동기발달형에 속 하며, 정소 조직상은 정원세포 국재형에 속한다. 암∙수 모두 첫 번째 군 성숙에 도달하는 크기는 전장 7.5 cm 로 조사되었다. 암컷의 생식소중량지수(GSI)는 7월에 12.83으로 가장 높았고, 9월에는1.08로 가장 낮았다. 수 컷의 GSI는 6월에 19.0으로 가장 높았고, 10월에 0.24 로 가장 낮았다. 암컷의 간중량지수(HSI)는 GSI와 같은 경향의 변화를 보였다. 가슴반점지수(TSI)는 생식소의 성숙 및 완숙시기인 5월부터 7월 사이에는 낮은 값을 보였다. 암컷의 생식주기는 성장기(3~5월), 성숙기 (5~6월), 완숙 및 산란기(6~8월), 회복기(8~11월) 그 리고 휴지기(11~3월)로 구분할 수 있었다. 수컷의 생식 주기는 분열증식 및 성장기(1~4월), 성숙기(4~5월), 완 숙 및 방정기(6~8월), 회복기(8~10월) 그리고 휴지기 (10~12월)로 구분할 수 있었다.
인 용 문 헌
An, C.M. 1995. Reproductive cycle of the spring-spawning bitterling, Rhodeus uyekii (Pisces : Cyprinidae). Korean J. Ichthyol., 7 : 33~42.
Baek, H.J. and T.Y. Lee. 1985. Experimental studies on the mechanism of reproductive cycle in the longchin goby, Chasmichthys dolichognathus (Hilgendorf ). Bull.
Korean Fish. Soc., 18 : 243~252.
Beams, H.W. and S.S. Sekhon. 1966. Electron microscope
studies on the oocyte of the fresh-water mussel (Ano- donta), with special reference to the stalk and mechani- sm of yolk deposition. J. Morphol., 119 : 477~502.
Billard, R., A. Fostier, C. Weil and B. Breton. 1982. Endo- crine control of spermatogenesis in teleost f ish. Can. J.
Fish. Aquat. Sci., 39 : 65~79.
Chung, E.Y. and K.Y. Lee. 1994. Structural and histoche- mical changes in the cyst cell and the interstitial cell in the testis of a teleost Hexagrammos agrammus asso- ciated with the reproductive cycle. Korean J. Ichthyol., 6 : 193~205.
Chung, E.Y. and S.Y. Kim. 1994. On the maturity and spawning of the greenling, Hexagrammos agrammus (Temmincki et Schlegel). Korean J. Ichthyol., 6 : 222~ 236.
Chung, E.Y., C.M. An and T.Y. Lee. 1991. Sexual matura- tion of the bluespotted mud hopper, Boleophthalmus pectinirostris (Linnaeus). Bull. Korean Fish. Soc., 24 : 167~176.
Chyung, M.K. 1977. The f ishes of Korea. Iljisa, Seoul, 727pp.
Colombo, G. and G. Grandi. 1995. Sex differentiation in the European eel: histological analysis of the effects of sex steroids on the gonad. J. Fish Biol., 47 : 394~413.
Deniel, C., C. Le Blanc, and A. Rodriguez. 1989. Compara- tive study of sexual cycles, oogenesis and spawning of two soleidae, Solea lascaris (Risso, 1810) and Solea impar (Bennet, 1831), on the western coast of Brittany.
J. Fish Biol., 35 : 49~58.
Elorduy-Garay, J.F. and S. Pamirez-Luna. 1994. Gonadal development and spawning of female ocean whitef ish, Caulolatilus princeps (Pisces: Branchiostegidae) in the Bay of La Paz, B.C.S., Mexico. J. Fish Biol., 44 : 553~ 566.
Gauld, J.A. and J.R. Hutcheon. 1990. Spawning and fecun- dity in the lesser sandeel, Ammodytes marinus Raitt, in the north-western North Sea. J. Fish Biol., 36 : 611~ 613.
Grier, H.J. 1981. Cellular organization of the testis and spermatogenesis in f ishes. Amer. Zool., 21 : 345~357.
Grier, H.J. 1984. Testis structure and formation of sper- matophores in the atherinomorph teleost Horaichthys setnai. Copeia, 1984 : 833~839.
Healey, M.C. 1971. Gonad development and fecundity of the sand goby, Gobius minutus. Trans. Amer. Fish.
Soc., 3 : 520~526.
Lahnsteiner, F., R.A. Patzner and T. Weismann. 1994.
Testicular main ducts and spermatic ducts in some cyp- rinid f ishes I. Morphology, f ine structure and histo- chemistry. J. Fish Biol., 44 : 937~951.
Lee, J.S., C.M. An and P. Chin. 1995. Sexual maturation of viviparous teleost surfperch, Ditrema temmincki.
Korean J. Ichthyol., 7 : 150~159.
Lee, J.S., C.M. An and P. Chin. 1996. Copulation and em- bryonic development of the viviparous teleost surfper- ch, Ditrema temmincki. J. Korean Fish. Soc., 29 : 26~ 34.
Lee, J.S., J.C. Kang and S.H. Huh. 1997a. Gonad structure and reproductive cycle of the smallmouth scorpionfish, Scorpaena miostoma (Teleostei: Scorpaenidae). J.
Korean Fish. Soc., 30 : 627~633.
Lee, J.S., S.Y. Kim, K.H. Ma and S.H. Huh. 2000. Repro- ductive cycle of the female red tongue sole, Cynoglossus joyneri (Teleostei: Cynog-lossidae). J. Korean Fish. Soc., 33 : (unpublished).
Lee, J.S., Y.K. Oh and S.H. Huh. 1997b. Fine structural observations on spermatogenesis of the goldeye rock- f ish, Sebastes thompsoni (Teleostei: Scorpaenidae). J.
Korean Fish. Soc., 30 : 1005~1012.
Lee, T.Y. and I. Hanyu. 1984. Reproductive cycle of small f ilef ish, Rudarius ercodes. Bull. Korean Fish. Soc., 17 : 423~435.
Lee, T.Y. and J.J. Jin. 1989. Studies on the f ishery biology of pomfrets, Pampus spp. in the Korean waters. 2.
Gonadal maturation and spawning. Bull. Korean Fish.
Soc., 22 : 266~280.
Lee, T.Y., Y.J. Kang and B.D. Lee. 1985. Reproduction and population dynamics of marbled sole, Limanda yoko- hamae. I. Reproduction. Bull. Korean Fish. Soc., 18 : 253~261.
Maisse, G., B. Mourot, B. Breton, A. Fostier, O. Marcuzzi, P.Y. Le Bail, J.L. Bagliniere and A. Richard. 1991.
Sexual maturity in sea trout, Salmo trutta L., running up the River Calonne (Normandy, France) at the ‘fin- nock’ stage. J. Fish Biol., 39 : 705~715.
Matsuura, S., M. Matsuyama, Y. Ouchi and T. Hidaka.
1987. Maturity classif ication and group maturity of the red sea bream Pagrus major. II. Male maturity. Mar.
Biol., 96 : 169~172.
Ntiba, M.J. and V. Jaccarini. 1990. Gonad maturation and spawning times of Siganus sutor off the Kenya coast:
evidence for def inite spawning seasons in a tropical f ish. J. Fish Biol., 37 : 315~325.
Saidapur, S.K. 1978. Follicular atresia in the ovaries of no- nmammalian vertebrates. Int’l Rev. Cytol., 54 : 225~ 244.
Shimizu, A. and I. Hanyu. 1982. Environmental regulation of annual reproductive cycle in a spring-spawning bit- terling Acheilognathus tabira. Bull. Jap. Soc. Sci. Fish., 48 : 1563~1568.
Shimizu, A. and I. Hanyu. 1983. Environmental regulation of spawning-period in an autumn-spawning bitterling Pseudoperilampus typus. Bull. Jap. Soc. Sci. Fish., 49 : 895~900.
Sugimoto, Y., Y. Takeuchi, K. Yamauchi and H. Takahashi.
1976. Induced maturation of female Japanese eels, An- guilla japonica by administration of salmon pituitari- es, with notes on changes of oil droplets in eggs of matured eels. Bull. Fac. Fish. Hokkaido Univ., 27 : 107
~120.
Tomasini, J.A., D. Collart and J.P. Quignard. 1996. Female reproductive biology of the sand smelt in brackish lagoons of southern France. J. Fish Biol., 49 : 594~612.
Treasurer, J.W. 1990. The annual reproductive cycle of
pike, Esox lucius L., in two Scottish lakes. J. Fish Biol., 36 : 29~46.
Wallace, R.A. and K. Selmen. 1981. Cellular and dynamic aspects of oocyte growth in teleosts. Amer. Zoo., 21 : 325~343.
Wiebe, J.P. 1969. Endocrine controls of spermatogenesis and oogenesis in the viviparous seaperch Cymatogaster aggregata Gibbons. Gen. Comp. Endocrinol., 12 : 267~ 275.
Wright, P.J. 1992. Ovarian development, spawning fre- quency and batch fecundity in Encrasicholina hetero- loba (Ruppell, 1858). J. Fish Biol., 40 : 833~844.
� ∙ . 1965. ヒイラギの と につ
いて. , 13(2) : 45~49.
Received : August 2, 2000 Accetped : September 12, 2000