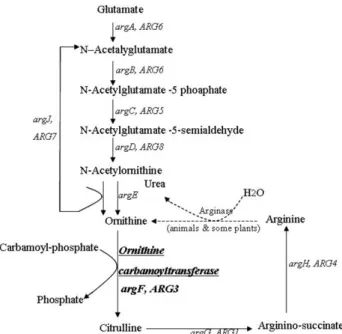
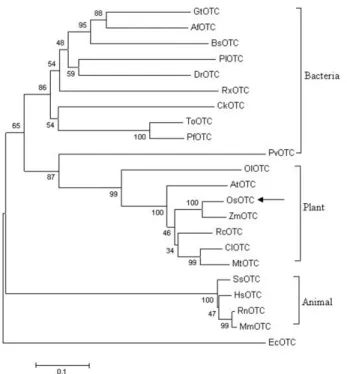
Characterization of a gene encoding ornithine carbamoyltransferase from rice
6
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
수치

관련 문서
For the dentists, 56 general practitioners and 60 residents, the affirmative responses were obtained from a survey, with a questionnaire composed by 52 values with regard to
mRNA expression of osteonectin, Runx2 and BSP(bone sialoprotein) in MC3T3-E1 cells cultured for 24 hours on 4 different titanium surfaces.. The mRNA were analyzed
‘small and medium-sized enterprises’ human resources development project for characterization high schools’ and ‘customized employment class’ based on
Quantitation of procollagen synthesis by Hs68 cells treated with fish collagen hydrolysates (FCHs), their <1 kDa fraction, and their >1 kDa fraction for 24 hrs.
The results show that the green seed extracts are only shown as antioxidants, but the others are shown as a prooxidants against DNA damage and Escherichia coli
The purpose of this study was to examine the effects of football on PAPS in sports club activity for 12 weeks in middle school boys.. The following
(1) PCR monitoring of bphC gene of Ralstonia eutropha H850 in soil Amount of bphC gene amplification of H850 cells was not generally proportional to the cell density
4 > Effects of Taro on COX-2 expression and iNOS expression(hot water) in human thyroid cancer cells. The cells were pretreated for 48hours with either
