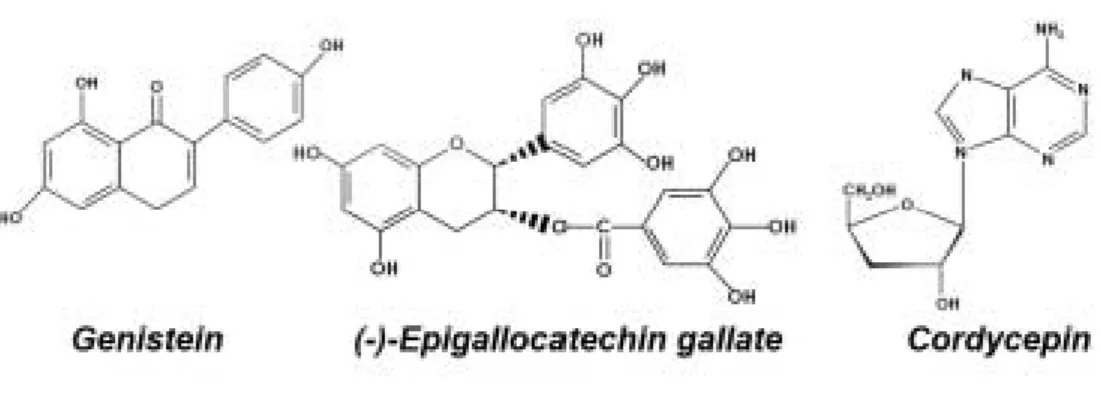
Anticancer effects of genistein, green tea catechins, and cordycepin on oral squamous cell carcinoma
10
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
(7)
(8)
(9)
(10)
수치


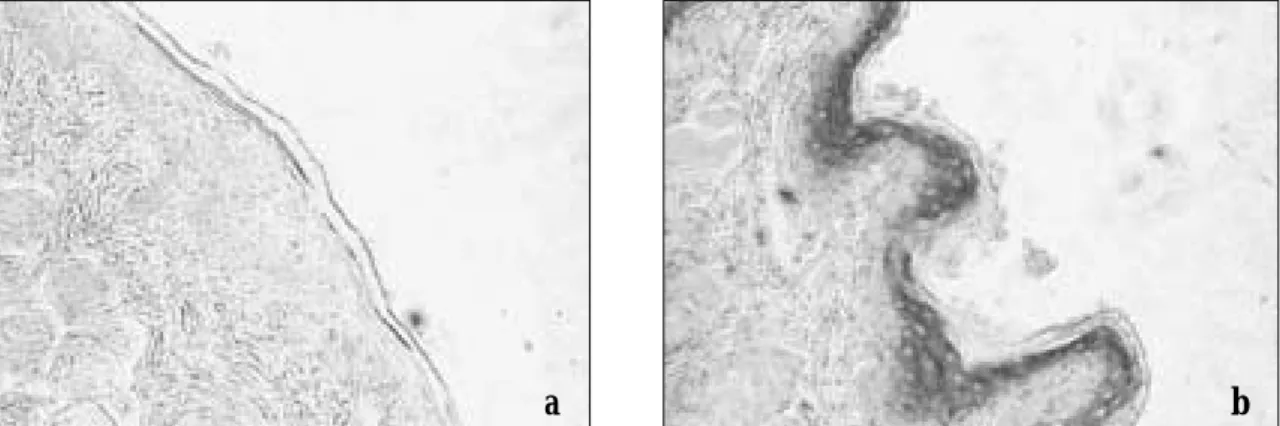
+2

관련 문서
Effects of Anthriscus sylvestris Hoffmann aqueous layer (ASAL) on cell apoptosis and PARP activity in NIH/3T3 fibroblast and KB, FaDu oral cancer cells by
In this study, the expression profiles of miRNAs were compared and analyzed for establishment of miRNAs related cancer cell growth inhibition in normal human oral
Anticancer activities of EF-24, a novel curcumin analog, on KB human. oral
Concentration-Dependent inhibition of cell viability by adenosine in human oral fibroblast and FaDu human head and neck squamous cell carcinoma · · ·
It also suggest that cell proliferation inhibits by a novel signal transduction for adenosine effect in human FaDu hypopharynx squamous cell carcinoma cells....
Differential Expression of Desmoglein1, Desmoglein3, Epithelial Membrane Antigen, Ber-EP4 and CD10 in Basal Cell Carcinoma and Squamous Cell Carcinoma
According to above, liquid based cytology is considered to be adequate for routine oral examination or diagnostic tool for oral dysplasia and carcinoma
Inhibitory effects of classified methanol extracts of Smilax china L on the COX-2 and iNOS expression of human colorectal cancer cell lines... Inhibitory
