약학회 지 제 43 권 제 6 호 743-750(1999) Yakhak Hoeji Vol. 43, No. 6
코카인 결함과 관련된 도파민 수송체의 아미노산 구조
장 미 윤 * • 전 대 준 * • 오 동 렬 ** • 이 용 성 * • 이상훈*우
*한양대학교 의과대학 생화학교실 및 정신건강연구소, **국럽정신병원 (Received July 15, 1999)
Amino Acid Structure of Dopamine Transporter Responsible for Cocaine Binding
Mi-Yoon Chang*, Dae-Joon Cheon*, Dong-Yul Oh**, Yong-Sung Lee* and Sang-Hun Lee*쌓
^Department of Biochemistry and Institute of Mental Health Hanyang University College of Medicine
**Seoul National Mental Hospital
Abstract - Human and bovine dopamine transporters (DAT) demonstrate discrete functional differences in the dopamine (DA) transport and cocaine binding. The functional analyses on the chimeras of human and bovine DAT have revealed that the region from the 133^ to 186*^ residue(encompassing the 3 ^ trans- membrane domain (TM)) is responsible for the substrate transport and cocaine binding. The present stud
ies have been done to find out the specific amino acid(s) which is essential for the binding of cocaine to DAT by interchanging the amino acids in that region between human and bovine DAT. When isoleucine, the 152™호 residue of chimera B3 (bovine DAT sequence), was transformed back to valine, the human DAT residue at the identical position, the cocaine binding was remarkably recovered to 98% of the human DAT values. In addition, the cocaine binding of the human DAT was decreased by 57% by substituting isoleucine for valine at position 152. When isoleucine at position 152 of the chimera B3 was converted to the other amino acids to provide an possible molecular basis for the functional role of the 152^^ residue, only the con
version to alanine among the amino acids tested significantly increased the cocaine binding by 34%, but these effects were not as much as those by the conversion to valine. In conclusion, valine at position 152 is a crucial amino acid for the interaction of cocaine to the DAT.
Keywords □ Dopamine transporter, cocaine binding, site-directed mutagenesis.
도파만(dopamine)은 대뇌에서 운동과 각종 호르몬 의 분비 및 행동양상 변화 등의 기능조절을 시행하고 있는 가장 중요한 신경견달물질 중의 하나이다. 신경 세포(presynaptic terminal)로의 도파민의 재롭수는 신 경언접 (synaptic)부위와 세포 외부의 도파민 농도틀 조 절하고, presynaptic 및 postsynaptic 수용체에서의 도 파민의 활성을 제한하는 역할을 하는 도파민 수송체 (DAT)라고 하는 특이적인 막단백질에 의해서 조절되 어진다(Kuhar and Zarbin, 1978; Kanner and Schul-
우 본 논문에 관한 문의는 이 저자에게로 (전화) 02-2290-0620 (팩스) 02-2294-6270
diner, 1987; Horn, 1990; Johnstone, 1990; Shima- da et a l, 1991; Uhl, 1992; Iversen, 1992). 도파민 수송체는 이와 같이 도파민의 신호 견달을 조절하는 정상 생리 기능 이외에도, 신경득, 항우울제, 및 코카 언 같은 정신홀분제의 작용부위가 된다. 그래서 코카 인 중득과 파킨슨씨병, 정신분열증, 및 Gilles de la Tourette syndrome 등의 정신질환과도 깊은 관련이 있는 것으로 보고되어 있다. 또한 여러 신경 득성 물 질들이 본 수송체를 통해 신경세포 내로 첨입되며, 항
^ 제 를 비롯한 자중 정신 신경계 약물들도 본 수송 체에 결합하식 그 작용을 나타내는 것으로 알려지고 있다(Ritz et al 1987: Bergman et al, 1989; Speal-
743
744 장미윤 ■ 전대준 ■ 오동■렬 • 이용성 • 상f
man et al, 1989). 특히 코카인은 도파민 수송체에 특 이적으로 결합하는데 이로 인해 도파민 흡수를 억제하 며 코카인 중독증을 유발한다(Rite et al., 1987:
Kuhar et al., 1991; Kuhar et al., 1992).
사람과 흰쥐의 도파민 수송체의 도파민 수송능과 코 카인 결합력은 비교적 높게 나타나고 있으나, 소 도파 민 수송체에서는 그 값이 사림께 비해 매우 낮은 것으 로 알려져 있다(Lee et al., 1996). 따라서 도파민 수 송체 단백질의 chimeric 접근을 통한 중간 도파민 수 송체의 기능적 다양성 연구는 도파민 수송체의 구조적, 기능적인 관계와 코카인의 인식 위치률 밝혀줄 수 있 을 것으로 관단된다. 이러한 가설 하에 사람파 소의 도파민 수송체 사어의 재 조합된 chimera틀 기능적으 로 분석한 결과, 사람 도파민 수송체 기눙의 절대적 역할을 하는 곳이 세 번째 세포막 통과부위(the 3-^
TM)을 둘러싼 부위라는 것은 이미 증명되었다(Lee et al., 1998). 그러나 이러한 연구는 도파민 수송체의 구 조적 , 기눙적 중요한 실마리는 제공하였으나 도파민 수 송체의 보다 상세한 작용기전을 설명하기에는 미흡했 다. 그래서 도파민 수용체의 기능을 더욱 명확히 하기 위해서 세 번째 세포막 통과부위 (the TM)을 둘러 싼 영역의 기능과 구조 사의의 관계룰 명확히 밝혀야 한다는 것이 제안되었으며, 본 연구는 코카인 결합력 과 관련하여 세 번째 세포막 통과부위 (the TM>i:
둘러싼 부위의 기능적 중요성에 기여하는 아미노산을 결정하는데 방향을 두었다. 이 부위내에서 사람 도파 민 수송체와 소 도파민 수송체 간의 54개의 아미노산 염기서열 중 치의를 보이는 곳은 7개가 있었으며, 본 실험에서는 chimera B3에서 7개의 소 도파민 수송체 의 아미노산을 site-directed mutagenesis에 의해 사 람 도파민 아미노산으로 번환 시켰다(Fig. 1). 또한 사 람 도파민 수송체에서 이 7개의 특정 아미노산 중 하 나률 그 특정 아미노산에 해당되는 소 도파민 수송체 의 아미노산으로 치환한 사람 돌연번이 도파민 수송체 를 제조하H 활성번화를 관찰하고 이를 자연형태 수송 체 및 chimeric 수송체의 활성과 비교 검토하였다.
실험방법
들연변어 도파민 수송체 유전자의 제조 - 먼저 Lee 등(1998)의 방법으로 도파민 수송체 chimera B3, 죽 사람 도파민 수송체의 염기서열 133번째에서 186번까
지의 영역인 세 번째 세포막 통과부위(the 3"^ TM)을 둘러싼 부위를 소 도파민 수송체의 같은 부위로 치환 한 chimeric 도파민 수송체 cDNA률 제조하고, 이률 이용하여 진핵세포 발현 유전자 pcDNA3.1(+) (Invitrogen Co., San Diego, CA, USA)-based pla
smid# 만들었다.
이 부위 (the 3"^ TM) 내에서 사람 도파민 수송체와 소 도파민 수송체 간의 54개의 아미노산 염기서열 중 차이를 보이는 곳은 7개가 있었으며, chimera B3에서 7개의 소 도파민 수송체의 아미노산을 site-directed mutagenesis에 의해 사람 도파민 아미노산으로 변환 시켰다. 또한 사람 도파민 수송체에서 이 7개의 특정 아미노산 중 하나률 해당되는 소 도파민 수송체의 염 기로 치환한 돌연번이 사람 도파민 수송체룰 제조하였 다. 이때 site-directed mutagenesis 는 QuickChange*근 site-directed mutagenesis system(Stratagene, L^oUa, CA, USA>을 사용하식 시행하였다.
Site-directed mutagenesis 후, 돌연번이 부위률 644-base-pair HindIII-ScaI(nucleotide 1-644) DNA 조각에서 분리하였고, chimera B3 cDNA에 삽입하였 다. 그 후 돌연변이된 조각의 염기서열은 Cy5"™
Autoread™ automatic sequencing system(Phamada Biotech, Uppsala, Sweden)을 사용하여 DNA 염기서 열을 결정함으로 확인하였다.
본 연구에서 사용된 용어들은 Chi B3ai52V>틀 에 로 들면, chimera B3에서 152번째 아미노산인 isoleudne을 valine으로 돌연번이 시켰다는 뜻이고, hDAT(V152I)는 사람 도파민 수송체에서 152번째 아 미노산인 valine을 isoleudne으로 돌연변이 시켰다는 뜻으로 사용하였으며, 다른 돌연번이의 용어에도 이와 같은 방법으로 적용하였다.
도파민 수송체의 코카인 결합력 측점 - 자연형태의 도 파민 수송체 cDNA와 돌연변이된 도파민 수송체 cDNA 률 각각 pCDNA3.1(+)에 삽입시키고 FuGENE™6 transfection reagent(Boehringer Mann- heim Co., Mannheim, Germany)를 사용하여 COS-7 세포에 발 현 시켰다. 그 후 24-well plate에 COS-7세포를 각 well당 3xi(y* 개씩 접종하고 10% fetal bovine serum이 함유된 Dulbecco's modified Eagle medium (DMEM)에서 24시간 동안 배앙했다. 각 well당 0.3 lag 의 DNA 률 0.5 |o/의 FuGENE™6 reagent 와 흔합하여 상온에서 15분간 배양하였다. 이 DNA-
J. Pharm . Soc. Korea
도파민 수송체의 야기노산 구조 745
FuGENE™ 6 복합체를 세포들에 첨가한 후 이틀간 배 양하고, 변형된 Krebs-Ringer HEPES(KRH) 완충액 [23 mM HEPES(pH 7.4), 125 mM NaCl, 4.8 mM KCl, 1.2 mM KH2PO4, 1.3 mM CaCl^, 1.2 mM MgS04, 5.6 mM glucose, 1 mM sodium ascorbate]
속에서 코카인의 유사물질인 2p-carbomethoxy-3p-(4- flurophenyDtropane ([^H]CFT, [크H]WIN35,428)의 결 합력을 측정하였다. 각 세포들은 변형된 KRH 완충액 으로 세척하고 lO nM 의 [크H]CFT(87 Ci/mmol, Du
pont-New England Nuclear Co., Boston, MA, USA)가 함유된 KRH 완충액에서 4°C로 2시간 동안 세포를 배양시켰으며, NaOH용액으로 세포률 용해시 키고 Liquid Scintillation Counter률 사용헤 방사선 양을 측정하였다. 이때 도파민 수송체 cDNA 가 삽입 되지 않은 PCDNA3.1 벡터를 동일한 조건하에 세포 내로 도입하고 동일한 방법으로 각 도파민 수송체의 각각의 수송능과 결합력 값으로부터 특이적 수송능과 결합력을 결정하였다.
실 험 결 과
Chimera B3의 단일 아미노산 치환 - 사람 도파민 수송체의 세 번째 세포막 통과부위 (the 3"* TM)를 포 함하는 부분(133번째부터 186번째 염기서열)을 소 도 파민 수송체로 치환시킨 chimera B3는 코?}인 결합력 이 사람 도파민 수송체의 20.3±5.5% 수준으로 감소
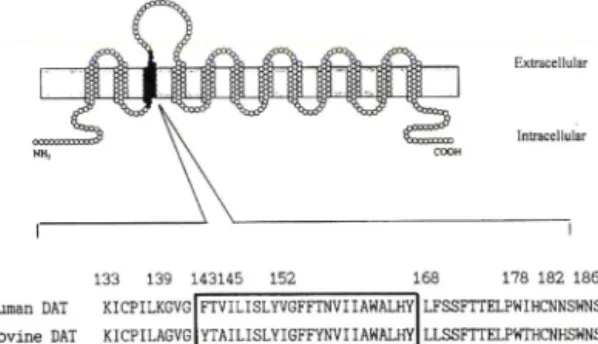
133 139 143145 Human DAT KICPILKGVG Bovine DAT KICPILAGVG
FTVILISLYVGFFTNVIIAWALHY miLISLYIGFFYNVI lAHALHY
168 178 182 186 LFSSFTTELPWIHCNNSWNS LLSSFTTELPWTHCNHSWNS
Fig. I - Schematic of the chimera B3 structure. The region consist of filled circles represents the bovine DAT sequences comprising from the 133"언to 186*^ posi
tion, whereas the portion of open circle represent the human DAT sequences. Comparison of primary amino acid sequences from the 133^^ to 186*^ amino acid residue in human and bovine DAT. The region in box indicate the 3*넌 transmembrane domain.
하였다(Lee et a l, 1998, Fig. 2). 이러한 코카인 결 합력의 감소에 중요한 역할을 하는 아미노산을 찾기 위 하여 chimera B3에서 소의 염기서열에 해당하는 각각 의 염기를 본래의 사람 도파민 수송체 염기로 되돌린 후 도파민 수송체의 활성도률 측정하였다.
Chimera B3 의 1st intracellular loop 139 번째 위처에 었는 alaninei: 같은 위처의 사람 도파민 수송 체 염기인 lysine으로 전환시켰을 때(Chi B3(A139K)), 코카인 결합력에 아무런 영향을 미처지 못하고 chimera B3 에서와 같이 낮게 관찰되었다(Fig. 2). 또 한 chimera B3의 2nd extracellular loop에 있는 아 미노산(168, 178, 182번째)둘을 위와 같은 방법으로 전환시켰을 때 코카인 결합력은 약간 상승하였으나 사 람 수송체의 활성에는 현저히 머처지 못하였다.
사람과 소의 도파민 수송체의 세번째 세포막 통과 부위에서 3가지의 아미노산에서 차이를 보이는데, 사 람 도파민 수송체의 143번째 phenylalanine과 소의 tyrosine, 145번째 valine과 alanine, 152번째 valine 과 isoleu- cine 등이 이에 해당된다. 이 중 , 143번째 와 145번째 chimera B3외 염기를 사람 도파민 수송체 아미노산으로 번환(Chi G (Y143F), Chi G(A145V)}시 켰을 때 코카인 결합력은 chimera B3과 벌 차어가 없
hOAT C h iB 3 A139K Y143F 시46 V L168F T 1 7 8 I H 18 2N
Fig. 2 - Cocaine binding activities of the chimera B3 mutants. Each of amino acids in the substituted region of chimera B3 (amino acid 133--186) was transformed back to that of the human DAT sequences, then followed by the functional assays.
The term 1152V as an example, refers to the mutation of 152"선 amino acid isoleucine to valine in chimera B3. The identical way of designation is applied to the other mutations. Cocaine bindings were determined by incubating with 10 nM [크H]CFT for 2 hours at 4'C, as described in the text. The column represent the means and the vertical lines represent SEM as determined from four to ten independent experiments performed in duplicate.
Vol. 43, No. 6, 1999
746 장미윤 • 견대준 이 • 이상훈
g .ED)
■D .E0Q
X
TT
rin
hDAT ChiB3 K139A F143Y V145A V152I F168L I178T N182H F ig . 3 - Effects of amino acid substitutions on the cocaine
binding activities of the human DAT. Cocaine binding assays were done on the human DAT mutants in which one of the seven amino acids in the region from the 13 3*언 to 186* residue was substituted by the respective one of bovine DAT.
The term V I 521 designate the mutation on the 152"크 residue of the human DAT from valine lo isoleucine.
The identical way of designation is applied to the other mutations. The column represent the means and the vertical lines represent SEM as determined from four to ten independent experiments performed in duplicate.
었다 (Fig. 2). 그러나 152번째 isoleudne을 valine으로 번환하였을 때 코카인 결합력이 사람 도파민 수송체의 활성도의 97.5±30.8%까지 회복하는 양상을 보였다 (Fig. 2). 이는 chimera B3의 152번째 아미노심<>1 기능 결함에 중요한 역할을 한다는 것을 보여주는 것이다.
사람 도파민 수송체의 단일 아미노산 치환 - 이번에 는 152번째 염기인 valine의 기능적 중요성을 재차 확 인하기 위하쉬 다른 방법의 돌연번이, 죽 사람 도파민 수송체 3"* TM 부위의 아미노산을 소 도파민 수송체 의 대응되는 아미노산으로 변환을 시켰다. Fig. 3에 나 타나•는 바와 같이, 152번째 valine을 isoleudne^-?.
번환한 hDAT(V152I)을 만들어서 코카인 결합력을 측 정하였을 때 사람 도파민 수송체의 42.7±11.1% 수준 으로 의미 있는 감소률 보였다. 한편 168번째 및 182 번째 아미노산의 처환시에는 약간의 코카인 결합력 감 소 현상을 관찰할 수 있었으나, 152번째 아미노산과 같은 현저한 효과는 관찰할 수 없었다. 그 외 사람 도 파민 수송체의 139, 143, 145, 178번째 아미노산을 소 도파민 수송체의 아미노산으로 번환 시에는 코카인 결합력에 의머 있는 변화가 없었다.
Chimera B3의 152번째 isoleudne의 처환 - 먼저, 도파민 수송체의 코카인 결합력과 관련된 152번째 아
hDAT Chi B3 1152V I152A I152G I152L I152D I152H
F ig . 4 - Effects of the substitutions of isoleucine to various amino acids at position 152 of the chimera B3. The conversions of isoleucine to the other monoamino-, monocarboxylic-amino acids or amino acid whose side groups are similar to that of leucine were followed by the functional assays for DAT. The column represent the means and the vertical lines represent SEM as determined from four to ten independent experiments performed in duplicate.
미노산 valine의 역할이 side chain의 길이에 영향받 는지 여부률 검중하기 위하쉬 chimera B3의 152번째 isoleucinei: 다른' monoamino, monocarboxylic 아미 노산으로 변환시켰다. 그 결과 isoleudne을 alanine으 로 번환시켰을 때 chimera B3의 코카인 결합력이 사 람 수송체의 50% 이상 수준으로까지 증가하였으나 valine^ 의 변환했을 때보다는 크게 미처지 못하였다 (Fig, 4). 그 외 leucine이나 glycine으로 번환 시에는 chimera B3의 활성에 전혀 영향을 주지 못하였다.
또한 도파만 수송체의 기능을 걸정하는 것이 152번 째 염기의 side group의 크기에 영향받는지를 검증하 기 위하여 isoleudne을 valine과 크기가 비숫한 aspartic acid나 histidine으로 번환시켜 보았으나 chimera B3 기능에는 큰 영향을 주지 못하였다. 이러 한 결과는 152번째 위처의 아미노산의 크기가 도파민 수송체의 코카인 결합기능을 결정하는 요인이 아니라 는 것을 말해 주는 것이다.
고 찰
도파만 수송체는 presynaptic neuron의 세포막에 존재하는 당단백으로서 중추신경계에서 기억, 운동신 호전달, 약물중독 등과 깊은 관련이 있는 도파민성 신 호전달을 조절하는 주체로 알려져 있다(Kuhar and
/. Pharm . Soc. Korea
도 민 수송체의 아 "lie산 구조 747
Zarbin, 1978; Kanner and Schuldiner, 1987; John
stone, 1990: Iversen, 1975). 또한 임상적으로 코카 인 중득과 파킨슨써병, 정신분열증 등의 정신과적 병 번과의 직접적인 관련성이 입증(Ritz et al, 1987:
Bergman et al, 1989; Spealman et al, 1989)됨으로 써 의학적으로 가장 관심이 집중되어지고 있다. 그 구 조는 분자량이 60-80 kDa 내외로 추정되며 12개의 세포막 통과 부위와 5개의 세포막내 고러 및 6개의 세포막외 고리로 구성되어 있다. 또한 2번째 세포막외 고리가 다른 고리에 비해 크고 N-말단과 C-말딘이 모 두 세포막내에 존재한다는 것이 특징이다. 지금까지 도 파민 수송체의 기능을 결정하는 기본 구조를 확인하려 는 여러 연구(Kitayama et al., 1992b; Kitayama et al., 1993; Wang et al., 1995; Buck et al., 1995;
Giros et al., 1994; Pifl et al., 1996; Lee et al., 1998)가 시행되어 왔으나, 아직 까지 명확하게 관명되 지 않고 있다. 도파민 수송체와 같은 monoamine 수 송체 계열로서 코카인 결합력을 갖는, 세로토닌 수송 체 (serotonin transporter: SERT)의 경우, 3번째 세포 막 통과 부위중 172 번째와 179 번째 isoleudne이 기능을 결정하는데 중요한 역할을 하고(Chen et al., 1997), 이 두 monoamine 수송체의 1 번째 세포막 통과 부위의 aspartate(쥐 세로토닌 수송체의 98 번째 와 사람 도파민 수송체의 79 번째)의 돌연번이체에서 는 기질 수송력과 코카인 결합력의 결함이 생기는 것 으로 보고 되었다(Barker et al., 1999, Kitayama et al., 1992). 현재의 연구돌은 도파민 수송체의 기질 수 송능과 코카인 결합력의 기견과 분자 구조를 밝히는 쪽으로 접근하고 있다.
본 연구에서는 도파민 수송체의 TM을 포함하는 부위(아미노산 133-186)가 수송체의 활성 발현에 중요 한 역할을 한다는 것에 기초초1식 이 부위에서 도파민 수송체의 코카인 결합력에 관여하는 특정 아미노산을 찾아내고자 하였다. 소수성 분석(hydrophobic an- alysis)에 기초룰 둔 도파민 수송체의 가상 구조도에 따르면(Uhl and Hartig, 1992; Amara and Kuhar, 1993; Giros and Caron, 1993), 아미노산 133번째에 서 186번째까지의 지역은 the intracellular loop (133-142), the 3"* TM(143-166), 및 the 2™오 extra
cellular loop(167-186) 등을 포함한다(Fig. 1).
사람 도파민 수송체에 소 도파민 수송체 3"** TM 부위를 포함한 부위 (133번째에서 186번째 아미노산)틀
치환시키면 도파민 수송체의 도파민 수송능과 코카인 결합력이 감소한다는 연구 결과(Lee et al, 1998)로 이 부위의 어떤 아미노산이 도파민 수송체의 이러한 기능감소에 결정적인 역할을 할 것이라는 가설을 세울 수 있었다. 사람과 소의 도파민 수송체의 이 부위 염 기서열을 비교해보면 54개중에서 7개만 다른 것을 알 수 있다. 따라서 이 7개의 서로 다른 아미노산을 site-directed mutagenesis를 통해 상호 교차 시켜 3^ TM을 둘러싼 부위의 기능적 중요성에 기여하는 아미노산을 결정하고자 하였다.
이들 화합물의 planar aromatic moieties가 소수성 결합 및 반데르발스결합시의 표면과 유사하다고 믿어 지고 있기 때문에 monoamine의 암모니움 말단이 수 송체의 옴전하률 지닌 아미노산와 연관돼 있다는 것은 알려져 있다(Maxwell et al., 1976; DeF^ulis et al., 1978). 이러한 가설 하에서는 사람 도파민 수송체의 168번째 염기인 phenylalanine이 도파민 및 코카인과 효율적으로 반응할 수 있고, chimera B3의 168번째 염기인 leucine은 그렇지 못하다고 가정되고 있다. 사•
람과 소의 도파민 수송체의 143번째와 178번째 아미 노산은 그 둘의 소수성 (hydrophobicity)에 있어서 상대 적으로 다르다. 143번째 아미노산은 사람 도파민 수송 체에서는 phenylalanine이지만 소 도파민 수송체에서 는 tyrosine고, 178번째에서는 사람에서는 isoleudne 이지만 소에서는 threonine이다. 아미노산의 소수성 (hydrophobicity)이 단백질의 기능적 구조를 결정하는 중요한 요소가 된다는 것을 고려한다면, 3"* TM을 포 함하는 부위의 처환으로 생기는 기능적 결함미 이 아 미노신들 때문일 것이라고 가정될 수 있을 것이다. 그 러나 본 연구의 결과는 이 이론적 가설에 부함되지가 앉았다. 먼저 사람과 소의 143, 168, 및 178번째 위 치의 염기를 서로 처환하였을 때 비록 168번째 염기 의 치환에서만 코카인 결합력에 약간의 영향을 주었을 뿐 다른 도파민 수송체의 코카인 결합에는 아무런 번 화를 주지 못하였다. 반면에 chomera G의 152번째 염기인 isoleudne을 valine으로 치환하였을 때 코카인 결합력이 사람 도파민 수송체의 값만큼 극적으로 증가 하였다. 마찬가지로 사람 도파민 수송체의 기능적 활 성도는 valine에서 isoleudne으로 치환하였을 때 거의 chimera B3 수준으로 감소함을 보였다. 이러한 발견 은 도파민 수송체의 152번째 염기가 사람 도파민 수 송체의 TM 부위를 포함한 부위의 전환 시에 관
Vol. 43. No. 6, 1999
748 장미윤 • 전대준 ■ 오동렬 • 이용성 ■ 이상훈
찰되는 기능적 결함에 책임이 있다는 것을 밝혀 주었 고, 그래서 152번째 염기인 valine이 사람 도파민 수 송체의 코카인 결합에 있어서 절대적이고 핵심적인 역 할을 한다는 것이 입증되었다.
Chimera B3과 사람 도파민 수송체의 152번째 염기 인 isoleudne과 valine을 비교해 보면 둘 다 side group이 단지 hydrocarbon diain으^ 구성된 rnono- amino-, monocarboxylic-아미노심;이라는 것을 알 수 있다. 그들의 소수성 (hydrophobicity)을 포함한 물리 화 학적인 성향은 거의 동일하다. 그러므로 이러한 치환 이, 물론 예외도 있겠지만, 도파민 수송과 코카인의 결 합에 영향을 주는 수송체의 2차적 흑은 3차적 구조의 근본적인 번화를 주지는 못하는 것으로 보인다. 그 대 신 단지 기질을 인식하고 견환시키는데 있어서는 어떤 특이한 화학적 그룹이 관여 할 것으로 생각되어진다.
이들 chimera B3과 사람 도파민 수송체의 152번째 염기인 isoleudne과 valine의 차이점은 단지 hydro
carbon chain의 길이와 side group이 차지하는 크기 에서만 다르다. Chimera B3의 152번째 위치의 isoleucinei- alanine으■로 전환 시 기능적 활성도가 의 미 있게 중가하였으나, valine으로 전환시의 값에는 못 미처는 결과였다. 반면 isoleudne을 glycine이나 leucine으로 견환 시에는 기능적 활성도에 영향을 주 지 못하였다.
또한 본 연구에서는 chimera B3의 isoleucine을 valine과 크기가 비숫한 aspartic acid와 proline으로 전환 시켜보았으나 chimera B3의 152번째의 aspartic acid 와 proline은 도파민 수송체의 기능을 회복시키지 못하였다. 이런 결과들은 152번째 염기외 크기가 도파 민 수송체의 코카인 결합을 특이화시키는데 중요할지 는 모르나 유일한 요소가 아니라는 것을 제시해 준다.
152번째 염기의 기능적 중요성과 더불어 본 연구의 결과는 182번째 위치의 asparagine와 168번째 위치의 phenylalanine이 코카인 결합에 관여한다는 것을 암시 해 준다. 그러나 아직 이들 아미노산의 구체적 기눙이 나 구조적 모델에 대해서는 더욱 많은 연구가 진행되 어야 할 것으로 관단된다.
본 연구에서는 코?K 1 결합과 관련하식 도파민 수송 체의 핵심적인 특정 아미노산을 밝혀주었고, 이번 연 구에서 밝혀진 염기와 관련된 더 고차원적인 구조가 밝혀진다면 도파민 수송체로의 도파민 및 코카인 작용 기견에 대한 분자생물학적 기초가 확립되어질 것으로
^>료된다.
결 론
본 연구에서는 도파민 수송체의 코카인 결합에 중요 한 역할을 하는 특이 아미노산을 규명하기 위하뇌, 사 람 도파민 수송체와 chimera B3의 아미노산을 서로 교차 번환시켜 돌연번이된 수송체의 코카인 결합력을 서로 비교, 평가하식 다옴과 같은 결론을 얻었다.
1. 3*■선 TM 부위률 포함한 부위(133번째에서 186번 째 염기)가 소의 아미노산 서열로 치환된 사람의 chimera B3 수송체 코카인 결합력은 사람 도파민 수 송체의 20.3±5.5% 수준으로 감소되었다.
2. chimera B3의 152번째 아미노산 isoleudne을 valine으로 처환하였을 때 코카인 결합력은 사람 도파 민 수송체 활성의 97.5±30.8%까씨 희복되었다.
3. 사람 도파민 수송체의 152번째 valine을 isoleu- dne으로 처환하였을 경우, 코카인 결합력은 자연형태 사람 도파민 수송체의 42.7±11.1% 수준으로 의미 있 는 감소를 보였다.
4. chimera B3 의 152번째 isoleudne 을 다른 monoamino-, monocarboxylic 이•미노산인 alanine으로 번환시컸을 때 코카인 결합력은 약간 증가하였으나 valine과 크기가 비숫한 aspartic acid나 histidine으로 번 환시켰을 경우에는 의미 있는 번화가 관찰되지 앉았다.
이상의 걸과는 도파민 수송체의 코카인 결합에 152 번째 아미노산 valine의 역할이 중요함을 의미한다. 또 한 valine의 기능이 아미노산 side chain의 크기에 따 른 공간배치에 있기보다는 전하에 의한 코카인과의 걸 합특이성을 결정짓는데 있옴을 시사하는 것으로 관단 된다.
문 헌
1) Amara, S. G. and Kuhar, M. J .: Neurotxansmitter transporters. Recent progress. Annu. Rev. Neurosci., 16, 73 (1993).
2) Barker, E. L., Moore, K. R., Rakhshan, E and Blakely R. D .; Transmembrane domaine I contributes to the permiation pathway for serotonin and ions in the serotonin transporter. J. Neurosci. 19, 4705 (1999).
3) Bergman, J., Madras, B. K., Johnson, S. E., and Spealman, R. D .: Effect of cocaine and related drugs
J. Pharm . Soc. Korea
^파민 수송체의 이미노산 구조 749
in nonhuman primates HI. self-adminstration by squirrel monkeys.
J Pharmacol Exp Ther
251, 150(1989).
4) Buck, K. J. and Amara, S. G .: Chimeric dopamine- norepinephrine transporters delineate structural domains influencing selectivity for catechoramines and l-methyl-4-phenylpyridium.
Proc. Natl Acad.
Sci USA
91, 12584 (1994).5) Buck, K. J. and Amara, S. G .: Structural domains of catecholamine transporter chimeras involved in selective inhibition by antidepressants and psycho
motor stimulants.
Mol Pharmacol
48, 1030 (1995).6) Chen J. G., Sachpatzidis A. and Rudnick G .: The third transmembrane domain of the serotonin transporter contains residues associated with substrate and cocain binding.
J. Biol Chem.
272,28321 (1997).
7) DePaulis, T, Kelder, D., and Ross S. B .: On the topology of the norepinephrine transport carrier in rat hypothalamus. The site of action of tricyclic uptake inhibitors.
Mol Pharmacol
14, 596 (1978).8) Giros, B., Mestikawy, S., Godinot, N., Zheng, K , Han, H., Yang-Feng, T. and Caron, M. G .: Cloning, pharmacological characterization, and chromosome assignment of the human dopamine transporter.
Mol Pbarmacol,
42(3), 383 (1992).9) Giros, B., and Caron, M. G .: Molecular characteriza
tion of dopamine transporter.
Trend in Pharm. Sci,
14, 43 (1993).
10) Giros B., Wang Y. M., Suter S., McLeskey S. B,, Pifl C., and Caron M. G .: Delineation of discrete domains for substrate, cocaine, and tricyclic antidepressant interactions using chimeric dopamine-norepinephrine transporters. /.
Biol. Chem,
269, 15985 (1994).11) Hom, A. S .: Dopamine uptake: a review of progress in the last decade.
Prog. Neurobiol
34, 387 (1990).12) Iversen L. L., In
Handbook of psychopharmacology.
New York, Plenum press, p. 381 (1975).
13) Iversen L. L .; Role of transmitter uptake mecha
nisms in synaptic neurotransmitter.
Crit Rev.
Biochem.,
22, 1 (1992).14) Kitayama S., Shimada S., Xu H., Markham L., Donoban D. M., and Uhl G. R .: Dopamine trans
porter site-directed mutations differentially alter substrate transport and cocaine binding.
Proc. Natl
Acad. Sci. USA
89, 7782 (1992).15) Kitayama S., Wang J. B., and Uhl G .: Dopamine transporter mutants selectively enhance MMP"^
transport.
Synapse
15, 58 (1993).16) Kuhar, M. J, and Zarbin, M. A .: Synaptosomal transporter, a chloride dependence for choline, GABA, glycine, and several other compounds.
J.
Neurochem.,
31, 251 (1978).17) Kuhar M. J., Ritz M. C., and Boja J, W.: The dopamine hypothesis of reinforcing properties of cocaine.
Trends. Neurosci.
14, 299 (1991).18) Kuhar, M, J ,: Molecular pharmacology of cocaine : a dopamine hypothesis and its implication.
Ciba Found Symp.,
166, 81(1992).19) Lee, S. H., Rhee J., Koh, J. K., and Lee, Y.
S .: Species differences in functions of dopamine transporter: paucity of MPP'^ uptake and cocaine binding in bovine dopamine transporter.
Neurosci.
Lett.
214, 199 (1996).20) Lee, S. H., Kang, S. S., Son, H., and Lee, Y. S .: The region of dopamine transporter encompassing the 3"언 transmembrane domain is crucial for function.
BBRC
246, 347-352 (1998).21) Maxwell, R. A., Ferris, R. M., and Burscu, J.
E .:
Mechanism of neuronal and extraneuronal tran
sport of catecholamines,
p. 95. Raven Press, New York (1976).22) Pifl, C,, Giros, B., and Caron, M. G .: The dopamine transporter, the cloned target site of Parkinsonism- inducing toxins and of drugs of abuse.
Adv. Neurol.
69. 235 (1996).
23) Ritz, M. C., Lamb, R. J., Goldberg, S. R. and Kuhar, M. J .: Cocaine receptors on dopamine transporters are related to self-administration of cocaine.
Science,
237, 1219 (1987).
24) Shimada, S., Kitayama, S., Lin, C. L., Patel, A., Nanthakumar, E., Gregor, R, Kuhar, M., and Uhl, G ,: Cloning and expression of a cocaine-sensitive dopamine transporter complementary DNA.
Science
254, 576 (1991).
25) Spealman, R. D., Madras, B. K. and Bergman, J .: Effect of cocaine and related drug in nonhuman primates H, Stimulant effects on schedule-controlled behavior. /,
Pharcol Exp Ther
261, 142 (1989).26) Uhl, G. R .: neurotransmitter transporters (plus)
Vol. 43, No. 6, 1999
정미윤 • 전대준 • 오동렬 • 이용성 • 이상훈
promissing new gene family. Trends PhrmacoL Sci 13, 421 (1992).
27) Uhl, G. R., and Hartig, E R .: Transporter explosion : uptake on uptake. Trends Pharmacol Sci. 13, 421 (1992).
28) Wang J. B., Moriwaki A., and Uhl, G. R .: Dopamine transporter cysteine mutants : second extracellular loop cysteines are required for transporter expres
sion. /. Neurochem. 64, 1416 (1995).
J. Pharm . Soc. Korea