Abstract
An abnormal mono-specific bloom of the cyanobacterium Microcystis aeruginosa had developed at a specific location (transitional zone, monitoring station of Hoenam) in Daecheong Reservoir from middle of July to early August, 2001. The maximum cell counts during the peak bloom reached 1,477,500 cells/mL, which was more than 6~10 times greater than those at other monitoring sites. The hypothesis of this study is that the timing and location of the algal bloom was highly correlated with the local environmental niche that was controled by physical processes such as hydrodynamic mixing and pollutant transport in the reservoir. A three-dimensional, coupled hydrodynamic and ecological model, ELCOM-CAEDYM, was applied to the period of development and subsequent decline of the bloom. The model was calibrated against observed water temperature profiles and water quality variables for different locations, and applied to reproduce the algal bloom event and justify the limiting factor that controled the Microcystis bloom at R3. The simulation results supported the hypothesis that the phosphorus loading induced from a contaminated tributary during several runoff events are closely related to the rapid growth of Microcystis during the period of bloom. Also the physical environments of the reservoir such as a strong thermal stratification and weak wind velocity conditions provided competitive advantage to Microcystis given its light adaptation capability. The results show how the ELCOM-CAEDYM captures the complex interactions between the hydrodynamic and biogeochemical processes, and the local environmental niche that is preferable for cyanobacterial species growth.
keywords : Algal bloom, Cyanobacteria, Daecheong Reservoir, ELCOM-CAEDYM, Microcystis
1. 서 론1)
저수지, 호수, 하구 같은 정체수역에서 식물플랑크톤이 과잉 번식하여 수면에 막(scum)을 형성하는 녹조현상(algal bloom)은 상수원 수질관리뿐만 아니라 수생태계의 건강성 측면에서도 매우 중요하게 다루어야 할 문제이다. 녹조발생 의 주요 원인종은 대부분 남조류(Cyanobacteria)이며, 우리 나라의 많은 댐 저수지에서는 여름철마다 남조류의 대량 증식에 의한 녹조현상이 자주 발생하고 있다(김범철 등, 2001; 김호섭 등, 2007; 박재충 등, 2005; 이정준 등, 2003;
허우명 등, 1991). 녹조현상은 저수지의 심미적인 경관을 훼손하고 상수원수에서 심한 이취미를 발생시키며, 일부 조 류종은 독성물질을 생성하는 등 수생태계의 균형을 파괴하 는 원인이 되고 있다.
대청호는 1981년에 준공되어 대전권과 청주권을 비롯한 인근 대도시와 중부권지역에 상수원수를 공급하는 매우 중 요한 수자원이다. 그러나 매년 하절기가 되면 남조류 녹조
†To whom correspondence should be addressed.
현상이 자주 발생하고 있어 수자원공급에 지장을 초래하고 있다. 대청호에 주로 출현하는 조류 종은 규조류, 갈색편모 조류, 남조류 등이다. 이 중 상수원수질관리에 가장 어려움 을 주는 것은 남조류인 Anabaena 와 Microcysits 이며, 주로 초여름에 출현하여 초가을까지 우점한다(천세억 등, 2006).
특히 적은 강우로 댐의 유입량이 급감한 2001년에는 평균 체류시간이 약 499일로 장기화됨에 따라 회남수역에 Microcysits 가 대량 번식하여 조류주의보(Chl-a 농도 15~25 μg/L 및 남조류 세포수 500~5,000 cell/mL 수준)가 35일, 조류 경보(Chl-a 농도 25 μg/L 이상 및 남조류 세포수 5,000 cell/mL 이상)가 47일, 조류 대발생(Chl-a 농도 100 μg/L 이상 및 남조류 세포수 1,000,000 cell/mL 이상)이 7 일간 발령되어 수질오염의 심각성이 본격적으로 대두되었 다(국립환경과학원, 2005).
대청호에서 남조류 녹조현상을 효과적으로 억제하기 위 해서는 녹조발생 기작에 대한 정밀한 원인분석이 우선되어 야 한다. 일반적으로 저수지에서 남조류 녹조발생의 원인은 영양염의 과도한 유입과 체류시간 장기화, 높은 표층 수온 과 성층화, 그리고 부력조절 능력과 같은 생리적 특성 등 으로 알려져 있다(Chaneva and Furnadzhieva, 2007; Rey-
nolds, 2006; Robarts and Zohary, 1987; Wetzel, 1983). 그 리고 질소와 인의 비, pH, CO2 농도도 중요한 영향인자로 보고된 바 있다(오광철 등, 1995). 한편, 성층화된 저수지에 서 녹조현상의 시공간적인 발생과 소멸 특성은 저수지의 지형적 특성, 강우와 같은 수문사상, 외부 및 내부 공급 영 양염류의 수리학적 이류확산 경로, 그리고 저수지 내에서 일어나는 다양한 물리적 혼합 기작과도 매우 밀접한 관계 가 있다(Chan et al., 2002; Donaghay and Osborn, 1997;
Robson and Hamilton, 2004; Spigel and Imberger, 1987).
대청호에서도 녹조현상이 발생하는 시기와 장소는 해마다 다르게 나타나며, 강우-유출의 규모와 높은 영양염을 포함 한 탁수 및 소옥천 유출수의 저수지내 수리학적 거동 특성 과 매우 깊은 관계가 있는 것으로 보고되고 있다(정세웅 등, 2007; 천세억 등, 2006; Ahn et al., 2003; Chung et al., 2008). 이렇듯 남조류 녹조발생에 미치는 영향인자는 다양 한 반면, 정기측정망 자료는 충분하지 못하여 녹조발생의 원인을 정확히 규명하기는 매우 어려운 실정이다. 특히, 저 수지의 수리현상과 수질환경의 상호작용은 다양한 물리적 현상에 의해 시시각각으로 변하므로 녹조발생을 정확히 예 측하기 위해서는 정밀한 현장계측이 요구되지만 비용 측면 에서 어려움이 있다. 따라서 이러한 문제점을 극복하고 현 장계측과 실험결과를 보완해석하기 위해서는 가상의 공간 에서 다양한 시나리오 분석이 가능한 3차원 수리・수질모델 링 기술의 활용이 효과적이다.
수리・수질모델링기술을 활용하여 저수지 수환경시스템을 해석하려는 시도는 지속적으로 이루어져 왔다(Ambrose et al., 1993; Cerco and Cole, 1993; Chapra, 1997; Chung et al., 2008; Cole and Buchak, 1995; Hamrick, 1992; Wallace and Hamilton, 2000). 그러나, 대부분의 경우 저수지를 연 직방향의 1차원(z 모델) 또는 연직-종방향 2차원(x-z 모델) 으로 가정하는 경우가 많았으며, 3차원(x-y-z) 모델을 적용 하는 경우에는 계산시간의 제약 때문에 수리와 수질을 분 리해서 모의하거나 단기해석에 그치는 경우가 많았다. 최근 들어 3차원 수리・수질모델링 기술은 컴퓨터기술의 발전과 함께 급속히 발전하고 있으며, 저수지의 수리-수질-생태계 시스템의 동적 연동해석(dynamic coupled simulation)을 가 능하게 하고 있다. 이들 모델 중 서호주대학교 물연구센터 에서 개발한 ELCOM(Estuary, Lake and Coastal Ocean Model)은 대청호와 같이 수심이 깊어 성층화가 있는 수체 에 적합한 범용 3차원 수리동력학 모델로써, 수질-생태계 해석 모델인 CAEDYM(Computational Aquatic Ecosystem Dynamic Model)과 연결되어 수리-수질-생태계의 동적 모의 가 가능하다(Botelho and Imberger, 2007; Chung et al., 2009; Hipsey et al., 2004; Hodges and Dallimore, 2006;
Wallace and Hamilton, 2000).
본 연구에서는 대청호에서 남조류의 녹조발생 시기에 일 어나는 수리학적 현상과 생물화학적인 현상의 상호작용에 대한 정밀 해석을 위해 ELCOM-CAEDYM 모형을 적용하 였다. 연구의 목적은 ELCOM-CAEDYM 모형을 이용하여 2001년에 발생한 저수지의 수문・수리현상을 재현하고, 이
러한 수리현상과 연계하여 남조류의 녹조현상에 영향을 미 치는 환경인자인 수온과 영양염류의 공간적 분포특성을 분 석한 후, 회남수역에서의 국부적인 남조류 대발생의 인과관 계를 규명하는데 있다. 본 연구결과는 대청호에서 빈번히 일어하는 남조류 녹조발생 기작에 대한 새로운 정보를 제 공함으로써 향후 효과적 제어대책 수립을 위한 기초자료로 활용될 수 있을 것으로 기대한다.
2. 연구방법
2.1. 연구대상지역
본 연구의 대상인 대청댐은 금강수계 중류에 위치하고 있으며 댐 하류지역의 홍수피해 경감, 용수공급 및 수력발 전 등 다목적으로 1975년 3월에 착공하여 1981년 6월에 준공되었다. 총 유역면적은(상류 용담댐 유역제외) 3,204.0 km2이고, 저수지 표면적은 72.8 km2로서 유역면적/저수면 적의 비는 44.0이다. 저수지의 평균 유입량은 95.6 m3/s, 강 우량은 1,230.0 mm/yr, 총 저수용량은 1,490×106 m3이며, 유효저수량은 790×106 m3에 해당한다. 대청호의 취수를 위 한 수리구조물은 추동과 문의에 위치한 취수탑(EL. 57.0 m) 2개와 댐의 발전 방류구(EL. 52.0 m)가 있으며, 그리고 홍수를 조절하기 위한 여수로(EL. 64.5 m)가 있다(한국수 자원공사, 2007).
금강물환경연구소에서는 저수지 내에서 조류예보제 시행 을 목적으로 대전과 청주의 상수원수를 취수하는 추동(A1) 과 문의(A2) 취수탑 및 회남수역(R3)에서 월간, 주간 및 필 요시 일간조사를 실시하고 있다. 또한, 한국수자원공사에서 는 조류감시와 강우시 유입하는 탁수의 거동을 파악하고자 월간 또는 주간조사를 장계교(R1), 대정리(R2), R3, 댐 앞 (R4), A1 및 A2에서 실시하고 있다(Fig. 1(a)).
조류대발생이 발령되었던 2001년 대청댐 유역의 연간 총 강수량은 794.6 mm이었고, 홍수기인 6~9월동안 강우량은 500.6 mm(연 강수량의 63.0%)이었다. 일평균 유입량은 34.6 m3/s이었고, 홍수기에는 52.1 m3/s이었으며, 저수위 일 변동의 최대값과 최소값은 각각 68.1 EL. m, 62.8 EL. m로 나타났다(Fig. 1(b)). 2001년 동안 대청호 유역의 총 강우량 과 일평균 유입량은 각각 예년 평균의 64.6% 및 36.2%에 해당하였고, 강우로 인한 유입량이 작아 홍수조절을 위한 여수로 수문방류는 없었다(국가수자원관리종합정보시스템, 2009).
2.2. ELCOM-CAEDYM 모형
ELCOM의 지배 방정식에는 운동량 방정식, 연속방정식, 자유수면 방정식, 물질 이류-확산방정식이 포함된다. 운동 량 방정식은 Reynolds-averaged Navier-Stokes (RANS) 방정 식을 사용하며, 기본적으로 유체의 특성에 대해서는 Bous- sinesq 가정과 수직방향의 운동량 방정식은 정수압(Hydro- static pressure) 이론을 가정한다. 그러나 비정수압(Non- hydrostatic)이 중요한 경우(예, 수면경사에 따른 동수압 작 용, 내부파 영향 등)에는 ELCOM의 비정수압 코드 옵션을
(a) (b)
Fig. 1. Locations of monitoring stations (a) and daily variations of hydrological factors in 2001 (b).
사용하여 수직방향의 운동량 방정식을 포함할 수 있다.
ELCOM에서 수평방향 유속의 순간 변동성분에 의한 난류 영향의 폐합식(turbulent closure)은 수평와점성계수(horizontal eddy-viscosity, )를 사용한다. 수직방향에 대해서는 확산 모델의 수직와점성계수(vertical eddy-viscosity, ) 또는 혼 합층 모델 (mixed-layer model)을 사용할 수 있다.
x-방향 운동량 방정식
′
(1)
y-방향 운동량 방정식
′
(2)
z-방향 운동량 방정식(Hydrostatic pressure assumption)
(3)
연속 방정식
(4)
자유수면 방정식
(5)
물질 이류-확산 방정식
(6)
여기서, u = 종방향 유속(m/s), v = 횡방향 유속(m/s), w = 연직방향 유속(m/s), p = 압력(N/m2), = 수위(m), = 밀도 (kg/m3), g = 중력(m/sec2), = 와점성계수(m2/sec), h = 수심 (m), = 와확산계수(m2/sec), c = 물질의 농도(mg/L)이다.
ELCOM의 수치해석 기법은 Casulli and Cheng(1992)이 개발한 TRIM (Tidal, Residual, Inter-tidal Mudflat)모델의 방법을 기본적으로 채택하고 있으나, 수치해석의 정확도를 보완하였으며, 스칼라 질량 보존, 수치확산 제거, 그리고 수직난류 혼합 모델 추가 등의 개선이 이루어졌다. 운동량 방정식은 준음해법(semi-implicit)을 사용하며 2차 방정식 Euler-Lagrange 보간법을 사용하여 해를 구한다. Euler- Lagrange 보간법을 위한 2차 방정식 적용은 내부파의 인위 적인 완화(damping)를 줄여주므로 성층화된 저수지에서 파 의 운동을 더 잘 모의할 수 있는 장점이 있다. 자유수면 방정식은 준음해법으로 차분화하며 TRIM 모델에서 사용한 공액경사법으로 해를 푼다(Casulli and Cheng, 1992). 수치 해석을 위한 격자(grid) 구성은 직교좌표체계를 채택한다.
격자망은 Arakawa C-grid를 사용하며, 속도장은 셀의 표면 에 그리고 자유수면 높이와 스칼라 농도값은 셀의 중앙에 서 정의된다. 각 수주(water column) 격자 셀의 자유수면 높이는 자유수면 방정식에 의해 계산된 결과에 따라 수직 으로 이동한다.
이류-확산방정식의 수평방향의 이류항은 ULTIMATE 기 법(Leonard, 1991)과 함께 3차 정확도를 갖는 QUICKEST 수치해석 방법을 사용함으로써 농도의 급격한 변화가 있는 영역에서 해를 안정적으로 구할 수 있다. 수평 확산항은 2 차 정도의 양해법을 사용하여 차분화 한다. 수직방향의 난 류혼합은 각각의 수주에 대하여 1차원 혼합층 모델(mixed- layer model)을 적용하여 해석한다(Hodges and Dallimore, 2006). 그 외에 수위에 의한 압력(barotropic)과 밀도 차에 의한 압력(baroclinic) 항, 지구자전에 의한 전향력 항, 바람 에 의한 응력, 대기-수면 온도차에 의한 대류 항, 그리고 유입수와 유출수 영향 등에 대한 계산이 포함된다.
CAEDYM은 다양한 수리모델과 쉽게 연결될 수 있도록 설계된 수질 및 수생태계 해석모델이다. CAEDYM과 ELCOM 수리모델의 연동해석(coupling)은 수체의 수온 구조 또는 밀도가 물의 탁도와 수질농도(부유물질, 염분도 등)에 의존 하는 경우 수질-수리 상호작용을 반영할 수 있다는 장점이 있다. 이 모델은 C, N, P, Si의 다양한 순환과정, DO(dis- solved oxygen) 순환, 입자크기별 6개까지의 부유물질(SS), 7개 종의 식물플랑크톤, 동물플랑크톤, 수서식물, 어류 등 을 모의 변수에 포함할 수 있다. CAEDYM은 조류의 일주 기 광합성과 야간 호흡 등의 과정을 해석하기 위해 어떤 시간단위로도 사용될 수 있으며, 일반적으로 수리모델과 같 은 계산간격(∆)으로 모델계산이 수행된다.
CAEDYM에서 DO 동력학은 대기와의 교환, 퇴적물 산 소요구량(SOD), 호기성 미생물에 의한 유기물 분해와 질산 화, 광합성에 의한 DO 생산과 호흡에 의한 소모, 그리고 추가 생물 모의 항목에 의한 호흡을 포함한다. 대기 교환 은 Wanninkhof (1992) 모델과 Riley and Skirrow (1974)의 플럭스 방정식에 기반하고 있다. 현재 CAEDYM은 간단한 정적 0차 SOD 모델을 사용하며, 수체의 수온과 DO 농도 에 따라 반응속도가 달라진다. 미생물의 활동은 유기 탄소 (DOC)를 CO2로 분해하며, 이 과정에서 화학양론비 만큼의 DO가 소모된다.
모델은 무기태와 유기태, 그리고 용존상과 입자상 형태의 C, N, P를 고려하며, POM(particulate organic matter)에서 DOM(dissolved organic matter)으로 그리고 다시 DIM (dissolved inorganic matter)의 형태로 분해되는 과정이 모의 된다. 질소의 순환은 탈질화, 질산화, 그리고 대기중 N2의 고착 과정이 추가로 고려된다. 규조류에 의한 용존 Si (SiO2) 의 생물학적 섭취는 생물 내부 Si pool (ISi)로 전환되며, ISi 는 침강과 재부상의 기작을 갖게 된다. 용존무기탄소(DIC) 수지는 추가적으로 대기와 수체의 CO2 분압 (pCO2) 차에 의 해 결정되는 CO2 플럭스를 포함한다. 대기-수체 경계면에서 CO2 전달은 Weiss (1974) 식에 의해 계산된 CO2 용해도를 가지고 Wanninkhof (1992)에 따라 계산된다. DIC(dissolved inorganic carbon) pool에서 CO2의 분율을 계산하기 위해서 Butler (1982)를 따라 탄소 완충 시스템, pH 그리고 알칼리 도가 모의된다. pCO2 계산을 위해 필요한 기체상과 액체상 의 CO2 값들은 Henry's Law에 의해 결정된다.
CAEDYM은 7개 그룹의 식물성플랑크톤을 모의할 수 있 다. 조류 생체량은 Chl-a (μg Chl-a L-1) 또는 탄소 (mg C L-1) 로 계산할 수 있다. 각각의 식물성플랑크톤 그룹의 성장속 도는 20°C에서 최대 성장률에 수온 함수와 빛, 인, 질소, 그리고 규소(규조류인 경우)에 의한 제한요소들 중 최소값 을 곱하여 결정된다. 식물플랑크톤의 생체량은 동물플랑크 톤, 어류, 그리고 조개류의 먹이로 줄어들 수 있다. 식물플 랑크톤 성장에 미치는 빛 제한 효과는 광저해(photoinhibi- tion) 또는 비광저해(non-photoinibited)로 모의될 수 있다.
유의할 만한 광저해가 없는 경우, Webb 등(1974) 방법이 탄소 고착의 최대 잠재율의 제한 분율을 산정하기 위해 사 용된다. 광저해가 있는 경우, 최대 생산을 위한 광 포화 값
은 수심 적분을 사용한다.
조류 그룹내에서 C, N, P 동력학을 모델링하는 방법은 2 가지가 있다. 하나는 일정한 C, N, P : Chl-a 비를 사용하 는 것이며, 다른 하나는 동적인 세포내 저장을 고려하는 것이다. 첫 번째 방법은 간단한 Michaelis-Menten 방정식이 조류 성장에 영향을 미치는 영양염류의 제한 정도를 산정 하는데 사용된다. 사멸과 분비에 의한 영양염류의 손실은 고정된 내부 영양염류(IN, IP)와 Chl-a 비와 조류 감소량의 곱으로 계산되며 다시 입자상 유기물(POM) pool 로 전환된 다. 반면에 두 번째 방법은 동적으로 세포내에 저장한 영 양염류를 사용할 수 있으며 Droop (1974) 모델을 사용하여 성장을 조절할 수 있다. 이 방법은 식물플랑크톤이 지정된 최소와 최대 값 범위내에서 동적으로 영양염류를 섭취하여 가변적인 내부 영양염류 농도를 가질 수 있도록 허용한다.
사멸과 분비에 의한 영양염류 손실은 첫 번째 방법과 같이 계산되지만, 내부 영양염류 농도는 동적으로 계산된 값이 사용된다. CAEDYM에는 식물플랑크톤의 이동과 침강을 위한 4가지 모델이 포함되어 있다. 이것은 고정 침강속도, 계산된 세포 밀도에 따라 계산되는 Stokes 침강, 광저해가 없는 유영, 그리고 광저해가 있는 유영이다. 바닥에 가라앉 은 식물플랑크톤은 재부상이 허용된다면, 사용자가 지정한 기간(통상 24시간) 동안 지속적으로 존재하는 것으로 가정 한다. 만약, 식물플랑크톤이 수체로 다시 들어오지 않는 경 우, 조류는 해당하는 퇴적층의 영양염류 pools로 들어간다.
2.3. 입력자료 및 매개변수의 구성
모델의 격자 구성은 금강 본류와 직접 유입하천인 소옥 천, 회인천, 주원천, 삼산천, 품곡천을 포함하여 흐름방향(x) 과 횡방향(y)에 대한 격자의 크기는 150 m × 150 m, 수심 방향은 1 m의 간격으로 나누었으며, 총 128,402개의 cell로 구성하였다(Fig. 2(a)). 구축된 지형자료의 신뢰도를 평가하 기 위하여 퇴사량 측정 보고서(한국수자원공사, 2006)에 제 시된 저수위-저수용량 관계와 모의결과를 비교하였다.
AME(Absolute Mean Error)와 RMSE(Root Mean Square Error)는 각각 0.279×109 m3, 0.531×109 m3로써 상시만수위 저수용량에 대해 각각 3.5%와 6.72% 오차에 해당하며, 결 정계수(Coefficient of determination)는 0.997로 나타났다 (Fig. 2(b)). 오차는 격자 구성 단계에서 지형의 단순화 과 정과 경계의 불확실성으로 발생하였다.
경계조건(boundary condition)중 본류 유입량과 댐 방류량 자료는 한국수자원공사에서 제공하는 실시간 수문자료를 사용하였으며, 각 지류 하천의 유량은 실측값이 없기 때문 에 유역별 면적비를 적용하여 산정하였다. 또한 유입수 수 온자료에 대해서는 선행연구(Chung and Oh, 2006)에서 제 시한 회귀식을 사용하였다. 기상자료의 풍향과 풍속은 대청 댐 인근에 설치된 자동기상관측소(대청 AWS)에서 측정된 자료를 이용하였으며, 대기 기온, 이슬점 온도, 태양 복사 에너지, 운도 등의 자료는 대전지방기상청의 자료를 사용하 였다.
모의 기간은 조류의 성장과 소멸 과정을 포함할 수 있도
(a) (b)
Fig. 2. Discretized Daecheong Reservoir grid (150×150 m) used for ELCOM simulation. (Color bar scale is in units of EL. m.) (a) and Comparison of observed and simulated reservoir capacity curves (b).
Table 1. Model parameters used for algae calibrations
Parameter Description Unit CYANO DIATOM GREEN
μmax Maximum specific growth rate /day 0.7 1.1 0.7
Ycc Average ratio of C to chlorophyll a mg-C/mg-Chl.a 40.0 40.0 40.0
ISt Light saturation for max. production uE/m^2/s 1300 70 200
Kpa Half saturation constant for P mg/L 0.005 0.005 0.005
Kna Half saturation constant for N mg/L 0.045 0.050 0.045
KCa Half saturation constant for C mg/L 2 2 2
INmin/max Min/max internal N concentration mg-N/mg-Chl.a 2.0/4.0 2.0/4.0 2.0/4.0 ICmin/max Min/max internal P concentration mg-P/mg-Chl.a 0.1/0.6 0.1/0.6 0.1/0.6 ICmin/max Min/max internal C concentration mg-C/mg-Chl.a 15/80 15/80 15/80
vT Temperature multiplier for growth - 1.08 1.06 1.06
Tsta Standard temperature ℃ 20.0 20.0 20.0
Topt Optimum temperature ℃ 29.0 25.0 27.0
Tmax Maximum temperature ℃ 34.0 32.0 33.0
kr Respiration rate coefficient /day 0.12 0.12 0.12
vR Temperature multiplier for respiration - 1.06 1.07 1.05
phvel Type of vertical migration algorithm - 0* 1** 1**
*Buoyancy control by Stokes Equation , **Constant settling (Diatom 0.1 m/d, Green 0.02 m/d) 록 2001년 4월부터 10월까지(7개월)로 하였다. 수온의 초기
조건은 대청호 내 주요 측정망 지점의 실측 수온자료를 바 탕으로 2001년 4월 초(Julian Day 91.5) 자료를 사용하였다.
ELCOM 모형의 수온 해석은 열역학적 이론에 근거한 물 리적 모형이므로 수온에 대한 별도의 보정을 하지 않았으 며, 매개변수 역시 초기값을 그대로 사용하였다. 물에 의한 태양의 단파복사에너지(파장범위 280~2,800 nm)의 광소멸 계수(Kw)는 근적외선인 NIR(Near Infrared Radiation, 720~
2,000 nm), 광합성에 유용한 PAR(Photosynthetically Active Radiation, 400~700 nm), 자외선 A(UVA, 320~400 nm)와 자외선 B(UVB, 300~320 nm)로 구분하여 입력되었다. 이 중 깊은 수심으로 침투 가능한 PAR는 Beer-Lambert Law
에 의해 수체내로 지수함수적으로 감쇠 침투하는 것으로 가정하였다.
수질항목별 경계조건 농도 자료의 경우 환경부 수질 측 정망의 월평균 농도 자료를 이용하였다. 조류농도의 모의를 위한 입력자료 구성은 조류의 계절적 천이과정을 고려하여 남조류(cyano), 규조류(diatom), 녹조류(green)의 총 3종으로 구분하였으며, 국립환경과학원에서 발행하는 조류예보제 시 행결과의 보고서를 참고하여 전체 Cell에 대한 각각의 종 별 Cell의 분율을 구하여 입력자료로 사용하였다. 각 조류 종의 모의를 위한 매개변수는 Table 1과 같다(Hipsey, 2010).
수질관련 매개변수의 보정은 각 조류 종의 최대성장속도(μ max)와 최적성장온도(Topt)에 대하여 실측값과 최대한 일
Table 2. Statistical indices used to evaluate the model accuracy
Statistical index Equation Desired value
Absolute mean error
0Root mean square error
0
Coefficient of determination
1
Oi= observed, Pi= simulated, Oave= average of observations, N = number of observations
치가 되도록 실시하였으며, 그 외 매개변수는 대부분 모델 의 초기값을 사용하였다. SOD는 실측 DO 농도의 수직 분 포 값을 이용하여 보정하였으며, 2.0 g/m2/d를 사용하였다.
2.4. 모형의 적합성 평가
모의결과와 실측자료의 비교 및 오차에 대한 평가는 AME, RMSE 및 R2를 이용하였다(Table 2). AME(절대평균오차) 는 실측값과 모의값 잔차의 절대값을 산술평균한 값으로 그 값이 0에 가까울수록 모의결과의 정확도가 높고, 모델 의 성능이 우수하다는 것을 의미한다. RMSE(평균제곱오차 의 평방근)는 실측값과 모의값의 제곱오차로부터 평균제곱 오차를 구하여 제곱근을 구한 것으로 모의결과의 평균오차 를 나타내는 지표이다. RMSE는 단위가 실제 변량의 단위 와 같고 값이 0에 가까울수록 모델의 성능이 보다 우수하 다는 것을 나타낸다. 또한, R2(결정계수)는 모의값과 실측값 의 선형 회귀관계의 적합도를 결정하는 계수로써 1에 근접 할수록 모델의 적합도 및 정확도가 높게 평가된다.
3. 결과 및 고찰
3.1. 모델의 보정
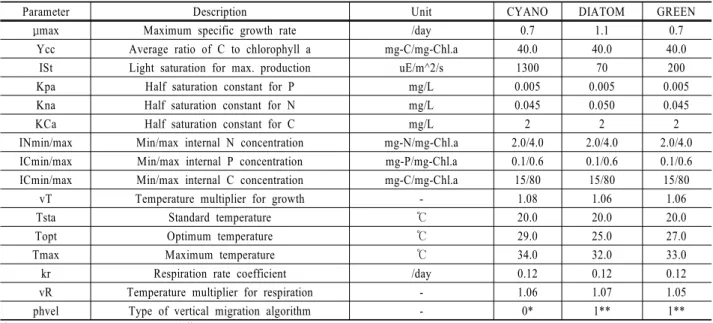
저수지의 수온성층 구조는 수체의 혼합, 영양염류의 공간 적 분포, 입자상 물질의 침강속도 및 조류의 성장속도 등 에 영향을 미치는 중요한 인자이므로 수질모델링에서 우선 보정되어야 한다. ELCOM 모형의 수온예측 성능을 평가하 고자 2001년에 R3과 R4 지점에서 수심별 수온의 실측값 및 모의값의 변동을 Fig. 3에 비교하였다. 홍수기 큰 강우 가 없었기 때문에 모델은 전 기간에 걸쳐 저수지 수온 성 층현상을 적절히 재현하였다. 모의값과 실측값의 오차는 회 남수역인 R3에서 AME 평균 1.575°C (범위 0.595~2.931°C), RMSE 평균 2.152°C (범위 0.782~3.973°C), R2 평균 0.945 (범위 0.786~0.993)이었다. 댐 앞지점인 R4에서는 AME 평 균 1.381°C (범위 0.320~2.126°C), RMSE 평균 1.808°C (범 위 0.416~2.873°C), R2 평균 0.961 (범위 0.763~0.995)을 보 였다. 하천수의 영향이 있는 회남수역보다 수심이 깊은 댐 앞에서 오차가 줄어드는 경향을 보인다. 수온 전이층에서 발생한 오차는 유입수 수온을 회귀식으로 산정하는 과정에 서 발생한 것으로 판단되며, 오차의 크기는 실측 수온 값
을 사용하면 개선될 것으로 예상된다.
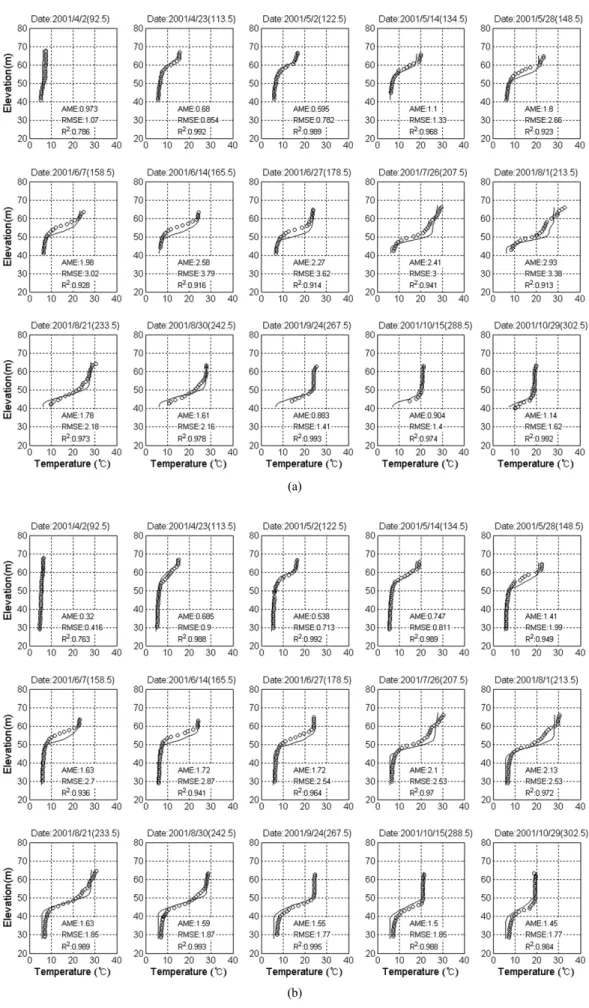
대청호 내 R3과 R4 지점에서 DO 농도의 실측 수직분포 와 ELCOM-CAEDYM에서 모의한 수직분포를 Fig. 4에 비 교하였다. 비교 결과, 모의값은 대체로 봄철의 완전 혼합상 태에서 성층화가 진행되면서 심층의 DO가 고갈되는 현상 을 잘 재현하는 것으로 나타났다. 그러나, 수온약층 인근에 서 모의값은 실측값과 유의할 만한 편차를 보였으며, 이는 저수지 상류부의 수심이 얕고 온도가 높은 퇴적층에서 유 기물 분해가 활발히 일어나면서 DO가 고갈된 후 댐 방향 으로 확산 진행하는 현상이 정밀하게 모의되지 못한 것이 원인으로 판단된다. 성층 저수지의 DO 공간분포는 대기로 부터의 재폭기, 유기물과 영양염류의 산화작용, 조류의 광 합성과 호흡, 퇴적물 산소요구량, 그리고 하천수 유입과 수 온 변동 등 다양한 물리적, 화학적, 생물학적 상호작용의 결과로 결정되어 정확한 예측이 매우 어렵다. 특히, 조류가 많이 성장하는 시기에는 표층 또는 유광층에서 DO 농도가 과포화되는 현상이 나타난다. DO 농도의 공간분포 모의를 정확히 하기 위해서는 정밀한 입력자료와 수리, 수질 생태 모델링이 요구된다. 본 연구에서는 유입수 DO 농도에 대 한 실시간 계측자료가 없어 월간 측정 자료를 사용함으로 써 기온과 수문상황에 따라 변화되는 하천수의 DO 농도를 정확히 모형에 반영하지 못하였다. 그리고 퇴적물의 산소요 구량도 공간적인 실측자료가 부족하여 일률적으로 2.0 g/m2/d 의 값을 사용한 것도 오차의 원인으로 판단된다. 제한된 입 력자료와 매개변수를 사용한 것에 반해, ELCOM-CAEDYM 은 성층화가 강하게 일어나는 여름철에 수온약층 이하 심 수층 구간에서의 DO 고갈에 따른 혐기성 상태로의 환경변 화를 잘 모의하였다. 이러한 현상은 혐기화된 퇴적층으로부 터의 인과 질소 그리고 철과 망간과 같은 광물의 용출 정 도에 직접적인 영향을 미치므로 저수지 수질관리를 위한 모델링에서 매우 중요한 기작에 해당한다. 그 이후 모의결 과는 가을 수직혼합 시기에 대기의 기온 하강과 함께 DO 가 재폭기되는 기작도 적합하게 재현하는 것으로 판단된다.
조류의 성장과 밀접한 관련이 있는 질소와 인의 시공간 적인 농도변화 모의성능을 확인하고자 T-N(total nitrogen), DIN(dissolved inorganic nitrogen), T-P(total phosphorus) 및 PO43--P 농도에 대한 ELCOM-CAEDYM의 모의값과 실측 값의 시계열 비교를 Figs. 5~8에 나타내었다.
(a)
(b)
Fig. 3. Comparison of simulated (line) and observed (circle) profiles of water temperature at the R3 (a) and R4 (b) in 2001.
(a)
(b)
Fig. 4. Comparison of simulated (line) and observed (circle) profiles of dissolved oxygen concentration at the R3 (a) and R4 (b) in 2001.
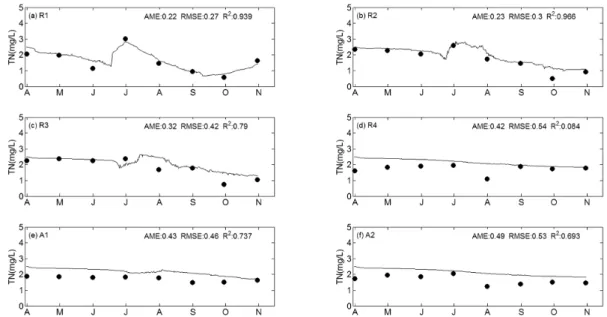
Fig. 5. Comparison of simulated (line) and observed (circle) time series of T-N concentrations in 2001.
(a) R1, (b) R2, (c) R3, (d) R4, (e) A1, and (f) A2
Fig. 6. Comparison of simulated(line) and observed(circle) time series of DIN concentrations in 2001.
(a) R1, (b) R2, (c) R3, (d) R4, and (e) A2 (No data for A2 site)
T-N의 경우, R1, R2, R3, R4, A1, A2 지점에서 모의값 과 실측값의 R2 값은 각각 0.939, 0.966, 0.790, 0.084, 0.737, 0.693으로써 R4 지점을 제외한 모든 지점에서 모의 값은 실측값의 시계열 변동경향을 잘 재현하고 있다. R4지 점에서 R2 값이 낮은 이유는 실측값이 시간에 따라 변화가 없이 일정한 값을 보이므로 실측값의 평균값을 사용한 예 측값과 모델을 사용한 예측값에 큰 차이가 없기 때문이다.
또한 모의값과 실측값의 오차크기를 나타내는 AME와 RMSE도 대부분 실측 T-N 농도값의 20% 내에 들어오는 것을 확인할 수 있다. 모델은 R1, R2 및 R3 지점에서 강 우 후 T-N 농도가 상승한 후 조류성장이 활발했던 7월부 터(Fig. 9 참조) 다시 감소하는 실측값의 경향을 잘 반영하 였다. DIN도 T-N의 경우와 유사한 변동 양상을 보였고, 실
측값을 잘 모의하는 것으로 나타났으며, 저수지내에서 T-N 의 구성성분은 대부분 무기질소가 차지하는 것을 실측값과 모의결과에서 모두 확인할 수 있다. DIN 농도는 녹조발생 이 가능한 최저 농도인 0.3 mg-N/L를 크게 상회하는 것으 로 나타났다(Wetzel, 1983).
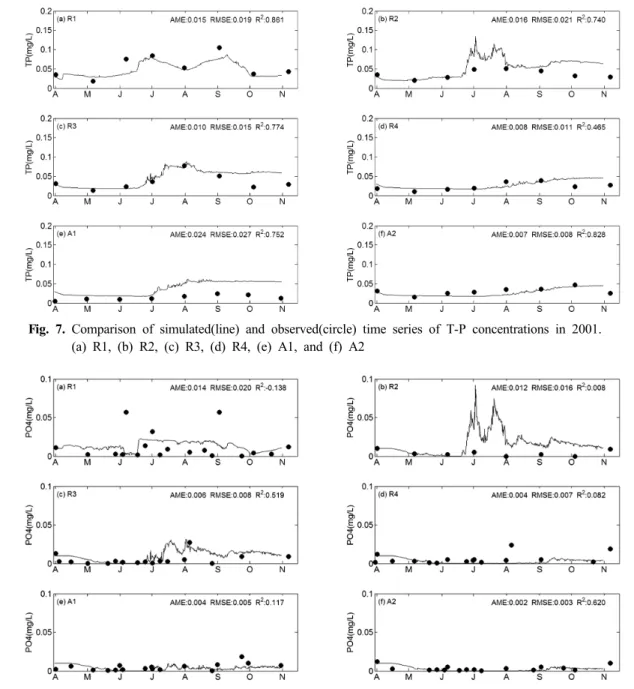
저수지 내 T-P 농도는 6월말과 7월 이후 발생한 강우-유 출 때문에 상승하는 경향을 보였으며, R1, R2, R3, R4, A1, A2 지점에서 모의값과 실측값의 R2 값은 각각 0.861, 0.740, 0.774, 0.465, 0.752, 0.828로써 모델은 실측값의 시 계열 변동 특성을 잘 재현하였다(Fig. 7). 특히 모델은 조류 대발생이 있었던 R3 지점에서 T-P 농도가 6월 말부터 증 가하여 8월 초에 최고값을 보이다가 점차 감소하는 실측값 의 경향을 잘 반영하는 것을 확인할 수 있다. 그러나, R1
Fig. 7. Comparison of simulated(line) and observed(circle) time series of T-P concentrations in 2001.
(a) R1, (b) R2, (c) R3, (d) R4, (e) A1, and (f) A2
Fig. 8. Comparison of simulated(line) and observed(circle) time series of PO43--P concentrations in 2001.
(a) R1, (b) R2, (c) R3, (d) R4, (e) A1, and (f) A2
과 A2 지점을 제외한 모든 지점에서 9월 이후 모델은 실 측 T-P 농도보다 대체로 높게 예측하는 경향을 보였다.
PO43--P 농도는 R1 지점의 일부 실측값을 제외하면, 대부 분 저수지 내에서 0.02 mg/L이하의 낮은 농도값을 보였으 며, R2를 제외한 대부분의 지점에서 모의값은 실측값의 범 위를 비교적 잘 재현하고 있다. 그러나 실측 PO43-
-P 농도값 이 매우 작고 시계열 변동성이 뚜렷하지 않아 모의값과 실 측값의 R2 값은 0.008~0.620의 낮은 범위를 보였다. R2 지 점에서 PO43--P 모의 농도값이 높게 나타난 원인은 강우시 소옥천에서 유입하는 높은 부하 때문이지만, 저수지내 월간 측정망 자료에는 이러한 현상이 나타나지 않았다. 강우시를 제외한 시기에 저수지내 표층에서 PO43-
-P의 실측 농도는 대부분 0.02 mg/L이하로 매우 낮아 조류 성장의 제한요인 으로 작용하는 것으로 판단된다. 유광층에서 낮은 PO43--P
농도 조건은 다른 조류 종에 비해 Microcystis 에 유리하게 작용한다(Reynolds, 2006). 특히, 강우시 수온약층 상부로 유입한 탁수에 포함된 높은 인 농도(한국수자원공사, 2009) 는 부력조절이 가능한 Microcystis 가 우점할 수 있는 유리한 조건을 제공한 것으로 판단된다(Reynolds et al., 1987).
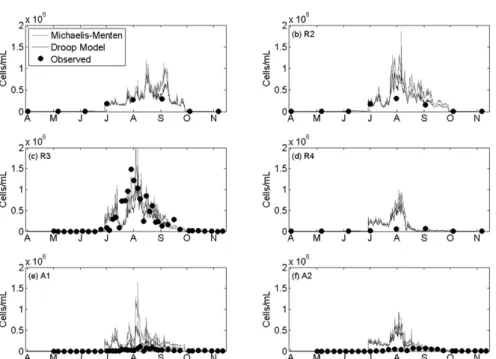
조류의 영양염류 동력학 모의를 위한 방법으로 Michaelis- Menten 방정식을 이용하는 방법과 Droop 모델을 사용하는 두 가지 방법을 모두 사용하여 모의를 하였으며, 그 결과 는 Fig. 9와 같다. 모의 결과는 2001년에 실측한 Chl-a 농 도와 Microcystis 셀 수의 관계를 이용하여 모의결과를 셀 수로 변환하여 비교하였다. 대청호에서 실측한 Chl-a 농도 와 Microcystis 셀 수의 관계 1.07×10-6 μg Chl-a/cell은 Reynolds (2006)가 제시한 0.36×10-6 μg/cell과 유사한 값을 보였다. 모의결과는 R3에서 7월말부터 8월초에 발생한 국
Fig. 9. Comparison of simulated(line) and observed(circle) time series of Microcystis concentrations in 2001.
(a) R1, (b) R2, (c) R3, (d) R4, (e) A1, and (f) A2
부적인 남조류 녹조현상과 9월에 들어 소멸하는 과정을 적 절히 반영하는 것으로 나타났다. 공간적으로 녹조현상은 R3와 그 상류지점인 R2에서 강하게 나타났으며, R4와 A1, A2 지점에서는 상대적으로 작았다. Droop 모델은 조류 생 체내 영양염류의 동적 모의를 고려할 수 있어, 설정된 최 소값과 최대값 범위 내에서 영양염류를 체내에 축적하여 사용할 수 있는 방법이다. 따라서 Droop 모델은 Michaelis- Menten 모델을 사용하는 것에 비해 조류가 주변 수체의 영양염류 농도변화에 덜 민감하게 반응하여 농도의 증가 폭이 상대적으로 작게 모의되었다. Fig. 9에 제시된 결과는 이러한 기작의 차이를 잘 반영하고 있으며, Droop 모델을 사용할 경우 좀 더 완만하고 작은 농도변화를 보이는 것을 확인할 수 있다. R3를 제외한 대부분의 지점에서 모델은 실측값을 과대 평가하는 것으로 나타났다. R1, R2, R4 지 점의 실측값은 모두 월간 측정망 자료이므로 모의결과와 비교하는데 한계가 있는 것으로 판단되며, A1과 A2 지점 은 취수탑 인근으로써 모델에서 반영하지 않은 폭기장치가 운영되고 있어 모의결과의 재현성이 떨어지는 것으로 판단 된다.
3.2. Microsystis 녹조발생 원인분석
보정을 거친 ELCOM-CAEDYM 모형을 이용하여 국부적 인 남조류 대발생의 인과관계를 파악하였다. 소옥천 유입수 는 대청호의 여름철 녹조발생에 큰 영향을 미치는 것으로 보고된 바 있다(천세억 등, 2006). 2001년 R3 지점에서는 6월 중순까지 조류농도에 큰 변동이 없었으나, 7월 중순부 터 급격히 증가하여 8월 초순에 Chl-a 농도가 최고 140 mg/m3까지 상승하였으며 그 이후 곧바로 감소하여 9월에 는 20 mg/m3 이하를 유지하였다. 그러나 소옥천이 합류하 는 추소리와 R3 지점에서 조류가 대발생한 것과 달리, R4
지점과 취수탑이 위치한 A1, A2 수역의 Chl-a 농도는 연중 20 mg/m3 이하를 유지하였다(국립환경연구원, 2001). 이러 한 결과는 소옥천 유입과 저수지 내에서 수리학적 혼합현 상과 밀접한 관계가 있을 것으로 판단되었다. 특히, 소옥천 은 하수처리장 방류지점이 저수지 인근에 위치하여 조류 성장에 직접 이용되는 무기인 농도가 금강본류 유입수보다 훨씬 높게 유입된다. 2001년의 경우, 소옥천에서 유입하는 인의 농도가 금강 본류에 비해 10배 정도 높았다(국립환경 연구원, 2001). 이는 가뭄이 심해 유역으로부터 소옥천 유 출량이 줄어들어 옥천 하수처리장 방류수의 기여도가 증가 한 것이 원인으로 판단된다. 이러한 높은 농도의 인이 강 우시 본류의 표층으로 유입되어 조류의 성장에 필요한 영 양분을 충분히 공급하고 있으며, 금강 본류와 소옥천이 합 류하는 지점인 추소리 지역은 정체구간을 형성하여 조류가 발생하기에 충분한 조건을 갖추고 있다.
Fig. 10에 2001년 7월 14일 조류경보 발생단계부터, 8월 4일 대발생을 거쳐 8월 10일의 대발생 소멸단계까지의 저 수지 내 표층에서 Microcystis 의 공간분포와 대발생 당시의 PO43--P 농도분포의 모의결과를 나타내었다. R3 지점의 표 층에 공급된 인은 조류의 섭취에 의해 소모되어 조류 대발 생 시기인 8월 4일에는 농도가 다시 낮아지는 경향을 보였 으며, 이는 다시 조류 성장의 제한요소로 작용하여 8월 10일 에는 조류 대발생은 다시 소멸단계로 접어들었다(Fig. 10(d)).
모의결과는 2001년에 R3 수역에 발령된 조류 대발생 경보 는 소옥천 유입과 저수지의 수리학적 혼합 특성에 큰 영향 을 받은 것을 확인해 준다. 하천 유입수의 밀도류 유동과 R3 지점에서의 국부적인 수직혼합은 R3 수역과 R4 수역에 서 서로 다른 영양염류의 환경이 조성되는 결과를 가져왔 으며, 인 농도의 측면에서 R3 수역은 부영양상태를 그리고 R4 수역은 빈영양상태를 보였다. 이러한 결과는 두 지점에
(a) Microcystis concentration at alarm stage (b) Microcystis concentration at maximum counts stage
(c) Microcystis concentration at grand algal bloom stage (d) Microcystis concentration at decline stage
(e) PO43-
-P concentration at grand bloom stage: plan view (f) PO43-
-P concentration at grand bloom stage: curtain view Fig. 10. Spatial distributions of Microcystis (mg Chl-a/m3) and PO43--P(mg/L) concentrations in Daecheong Reservoir during
the cyanobaecteria bloom in 2001.
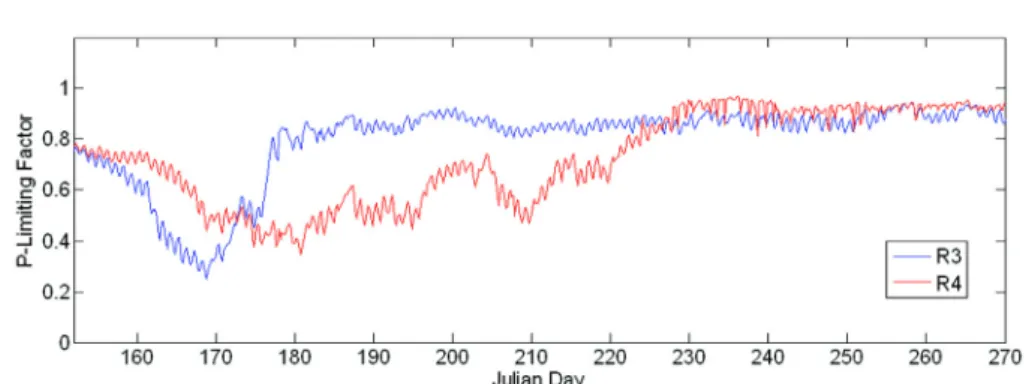
서의 조류성장에 필요한 제한 영양염인 인 농도의 제한정 도를 서로 비교해 보면 더욱 확실하게 알 수 있다(Fig. 11).
그림에서 보듯이 R3 수역에서는 홍수 유입이후 인 농도에 의한 조류성장 제한이 크지 않은 반면, R4 수역은 인 농도 가 조류성장의 중요한 제한요인으로 작용하고 있다.
Fig. 12는 2001년 조류 대발생 기간 동안에 대청호에서 발생한 남조류, 규조류, 그리고 녹조류의 공간적 분포 모의 결과를 비교하여 보여준다. 남조류는 회남수역에 밀집하여
대발생을 일으킨 반면, 규조류와 녹조류는 저수지 유입부에 서 낮은 농도로 나타나고 있다. 조사결과에서도 볼 수 있 었듯이 조류 대발생 시기에 대청호에서는 남조류인 Micro- cystis 가 절대적으로 우점 하였으며, ELCOM-CAEDYM 모 의결과는 이러한 조사결과와 매우 잘 일치하고 있다. 규조 류와 달리 남조류인 Microsystis 는 세포내 기낭이 있어 수 체내에서 광합성과 호흡과정에서 부력조절이 가능하다. 이 러한 조류의 생리적 특성은 성층화된 저수지에서 다른 조
(a) Microcystis plan view (b) Microcystis curtain view
(c) Diatom plan view (d) Diatom curtain view
(e) Green algae plan view (f) Green algae curtain view
Fig. 12. Spatial distributions of Microcystis, Diatom and Green algae during grand algal bloom at Daecheong Reservoir in 2001.
류 종에 비해 상대적으로 우위의 경쟁력을 가지게 한다.
즉, 광합성을 통해 세포내에 탄수화물이 축적되면 밀도가 증가하여 침강하고, 다시 호흡과정에서 탄수화물이 소모되 면 밀도가 감소하여 부상하게 되어 표층의 빛과 표층 아래 의 영양염류를 모두 활용할 수 있는 특혜를 누리게 된다 (오광철 등, 1995). 그러나 규조류와 같이 부력조절 기능이 없는 조류는 바람에 의한 수직혼합이나 대류 혼합이 발생 하여야 수동적(Passive)으로 부상할 수 있다. 그러나 2001 년과 같이 성층강도가 강하며 바람이 약한 상태에서는 침 강할 수 밖에 없어, R3지점에서는 Microsystis 가 우점하게 된 것으로 판단된다.
4. 결 론
본 연구에서는 2001년 대청호에서 발생한 Microcystis 녹 조현상의 원인해석을 위해 3차원 수리-수질 연동해석 모델 인 ELCOM-CAEDYM을 구축하였으며, 2001년에 발생한 저수지의 수문·수리현상을 재현하고 수리현상과 연계하여 남조류의 녹조현상에 영향을 미치는 환경인자와 영양염류 의 공간적 분포특성을 분석하였다. 본 연구를 통해 도출된 주요결론은 다음과 같다.
1) 모델은 2001년 조류 대발생 당시 저수지 내에서 수온과 DO 농도의 수직분포, 그리고 영양염류 농도의 시공간적 변화를 적절히 재현하였으며, 회남수역(R3)에서 발생한 국부적인 남조류 녹조현상을 적절히 모의하였다. 그러나 그 외 지역에서는 모의값이 실측값보다 높게 예측되는 경향을 보였다.
2) 조류의 영양염류 동력학 모의를 위한 방법으로 Michaelis- Menten 모델과 Droop 모델을 비교한 결과, Droop 모델 을 사용할 때 Michaelis-Menten 모델을 사용하는 것 보 다 조류농도가 주변 수체의 영양염류 농도변화에 덜 민 감하게 반응하였다.
3) 대청호 호수역(Lacustrine zone)은 여름철에 강우에 의해 외부로부터 인이 공급되지 않으면 대체로 PO43--P 농도 가 매우 낮은 빈영양 상태를 보인다. 그러나 강우시 유 입한 하천수에 포함된 높은 인이 표층의 유광층으로 공 급될 경우, 즉시 조류가 급성장하는 경향을 보이며 조류 의 성장과 함께 표층의 PO43--P 농도는 다시 급격히 감 소하는 것으로 나타났다.
4) 2001년의 경우 조류성장의 제한요소인 인은 소옥천으로 부터 강우시 부하되었으며, 유입하천수의 수리학적인 밀 도류 유동특성 때문에 회남수역에서 수직혼합되면서 유 광층에 인이 공급되었고, 그 결과로써 회남수역에서는 남조류인 Microcystis 농도가 급격히 증가하였다.
5) 반면, 강우-유출 후 회남수역에서 나타난 부영양상태와 대조적으로 댐앞 수역은 상대적으로 인 농도가 매우 낮 은 빈영양상태를 보였다. 이것은 저수지의 수리학적 현 상과 국부적인 수질환경이 매우 밀접하게 관계되어 있 음을 보여주는 결과이다.
6) 따라서, 대청호에서 남조류 녹조발생의 기작을 정확히
해석하고 효과적인 녹조제어 대책을 수립하기 위해서는 다양한 수문사상과 홍수규모에 대한 오염물질의 이류-확 산 해석과 수리-수질-생태계의 연동 모델링 연구가 필요 하다.
사 사
이 논문은 2009년도 충북대학교 학술연구지원사업의 연 구비지원에 의하여 연구되었음. 본 연구를 위해 ELCOM- CAEDYM 모델을 지원해 주신 서호주대학교 물연구센터 Imberger 교수님께 감사드립니다.
참고문헌
국가수자원관리종합정보시스템(2009). http://www.wamis.go.kr/.
국립환경과학원(2005). 대청호 수질 및 조류발생 실태조사.
국립환경연구원(2001). 금강수계 상수원 수질조사 2001.
김범철, 박주현, 허우명, 임병진, 황길순, 최광순, 최종수(2001).
국내 주요 호수의 육수학적 조사(4): 주암호. Korean J.
Limnol., 34(1), pp. 30-44.
김호섭, 황순진, 공동수(2007). 부영양 저수지에서 남조류의 발달과 천이 및 영향 요인. Korean J. Limnol., 40(1), pp.
121-129.
박재충, 박정원, 김종달, 신재기(2005). 안동호에서 환경요인과 식물플랑크톤의 시・공간적 변동. Algae, 20(4), pp. 333-343.
오광철, 오희목, 이진환, 맹주선(1995). 대청호 식물플랑크톤 의 일주성 수직운동. Korean J. Limnol., 28(4), pp. 437-446.
이정준, 이정호, 박종근(2003). 대청호의 남조류 수화 발달과 환경요인 변화와의 상관 관계. Korean J. Limnol., 36(3), pp. 269-276.
정세웅, 박재호, 김유경, 윤성완(2007). 대청호 부영양화 모
의를 위한 CE-QUAL-W2 모델의 적용. 수질보전 한국물
환경학회지, 23(1), pp. 52-63.
천세억, 이재안, 이재정, 유영복, 방규철, 이열재(2006). 대청
호 유입유량 변동과 수질 및 조류증식의 관계. 수질보전
한국물환경학회지, 22(2), pp. 342-348.
한국수자원공사(2006). 대청댐 퇴사량측정보고서.
한국수자원공사(2007). 댐운영 실무편람.
한국수자원공사(2009). 대청호 조류제어 차단막 설치효과
예측 및 연구.
허우명, 김범철, 조규송(1991). 소양호 부영양화에 따른 N/P 비의 변화와 남조류 Bloom. Korean J. Limnol., 24(4), pp.
283-288.
Ahn, C. Y., Kim, H. S., Yoon, B. D., and Oh, H. M. (2003).
Influence of Rainfall on Cyanobacterial Bloom in Daecheong Reservoir. Korean J. Limnol., 36(4), pp. 413-419.
Ambrose, R. B., Wool, T. A., and Martin, J. L. (1993). The Water Quality Analysis Simulation Program, WASP5, Part A: Model Documentation. U.S. EPA Center for Exposure Assessment Modeling, Athens, GA.
Botelho, D. A. and Imberger, J. (2007). Dissolved-oxygen response to wind-inflow interactions in a stratified reservoir.
Limnol. Oceanogr., 52, pp. 2027-2052.
Butler, J. N. (1982). Carbon Dioxide Equilibria and Their Appli- cations. Addison-Wesley, Massachusetts, USA.
pp. 537-544.
Chapra, S. C. (1997). Surface Water-Quality Modeling. McGraw- Hill Inc., New York.
Chung, S. W. and Oh, J. K. (2006). Calibration of CE-QUAL- W2 for a monomictic reservoir in monsoon climate area.
Water Science and Technology, 54(12), pp. 29-37.
Chung, S. W., Hipsey, M. R., and Imberger, J. (2009).
Modelling the propagation of turbid density inflows into a stratified lake: Daecheong Reservoir, Korea. Environmental Modeling and Software, 24, pp. 1462-1482.
Chung, S. W., Lee, H. S., and Jung, Y. R. (2008). The Effect of Hydrodynamic Flow Regimes on the Algal Bloom in a Monomictic Reservoir. Water Science and Technology, 58(6), pp. 1291-1298.
Cole, T. M. and Buchak, E. M. (1995). CE-QUAL-W2: A Two-dimensional, Laterally Averaged, Hydrodynamic and Water Quality Model, User's Manual. U.S. Army Engineers Water- ways Experiment Station, Vicksburg, MS.
Donaghay, P. L. and Osborn, T. R. (1997). Toward a theory of biological-physical control of harmful algal bloom dynamics and impacts. Limnol. Oceanogr., 42(5), pp. 1283-1296.
Droop, M. R. (1974). The nutrient status of algal cells in continuous culture. J. Mar. Biol. Assoc., UK 54, pp. 825-855.
Hamrick, J. M. (1992). A three dimensional environmental fluid dynamics computer code: Theoretical and computational aspects. Special report, The college of William and Mary, Virginia institute of marine science, Glouceslter point, VA.
Hipsey, M. R (2010). Computational Aquatic Ecosystem Dynamics Model: CAEDYM v3. v3.2 User Guide. Centre for Water Research, University of Western Australia.
Hipsey, M. R., Antenucci, J. P., Brooker, J. D., Burch, M. D.,
Reynolds, C. S., Oliver, R. L., and Walsby, A. E. (1987).
Cyanobacterial dominance: the role of buoyancy regulation in dynamic lake environments. New Zealand Journal of Marine and Freshwater Research, 21, pp. 379-390.
Riley, J. P. and Skirrow, G. (1974). Chemical Oceanography.
Academic Press, London.
Robarts, R. D. and Zohary, T. (1987). Temperature effects on photosynthetic capacity, respiration, and growth rates of bloom-forming cyanobacteria. New Zealand Journal of Marine and freshwater Research, 21, pp. 391-399.
Robson, B. J. and Hamilton, D. P. (2004). Three-dimensional modelling of a Microcystis bloom event in the Swan River estuary, Western Australian. Ecological Modeling, 174, pp.
203-222.
Spigel, R. H. and Imberger, J. (1987). Mixing processes relevant to phytoplankton dynamics in lakes. New Zealand Journal of Marine and Freshwater Research, 21, pp. 361-377.
Wallace, B. B. and Hamilton, D. P. (2000). Simulation of water-bloom formation in the cyanobacterium Microcystis aeruginosa. Journal of Plankton Research, 22(6), pp. 1127- 1138.
Wanninkhof, R. (1992). Relationship between gas exchange and wind speed over the ocean. J. Geophys. Res., 97, pp. 7373- 7381.
Webb, R. A., Kleinberg, R. L., and Wheatley, J. C. (1974).
Experiments on Dynamic Parallel Magnetism in Superfluid 3He. Phys., Rev., Lett., 33, pp. 145-148.
Weiss, R. F. (1974). Carbon dioxide in water and seawater: the solubility of a non-ideal gas. Marine Chem., 2, pp. 203-215.
Wetzel, R. G. (1983). Limnology, Saunders College Publishing, Philadelphia, PA.