Ecology of Vesicular-Arbuscular Mycorrhizal (VAM) Fungi in Coastal Areas of India
Thangaswamy Selvaraj1,2 and Hoon Kim*
Department of Agricultural Chemistry, Sunchon National University, Suncheon 540-742, Korea
1Research Institute of Agricultural Sciences, Sunchon National University, Suncheon 540-742, Korea
2Centre for Advanced Studies in Botany, University of Madras, Guindy Campus, Chennai - 600 025, India Received February 3, 2004; Accepted April 21, 2004
A survey was undertaken in three coastal saline soil sites of the southern penninsular tip of India for the occurrence and quantification of vesicular-arbuscular mycorrhizal (VAM) fungi. Sixteen VAM fungal species were isolated from rhizosphere soils of 15 plant species. To confirm infection efficiency, woody legume Prosopis juliflora present commonly in the three sites was inoculated in field with selected VAM fungal spores. Glomus macrocarpum was found as the most efficient VAM species. VAM fungi enabled all vegetations, Prosopis juliflora in particulr, to survive in the disturbed saline-stressed ecosystem.
Key words: Coastal area, colonization, vegetation, rhizosphere soil, VAM fungi.
Mycorrhizae are ubiquitous symbioses between fungi and plants in natural and man-made ecosystems, such as tropical forests, agrosystems, savanas, semideserts, and sand dunes.1) The term vesicular-arbuscular mycorrhiza (VAM) denotes the formation of special structures, namely vesicles and arbuscules, by the colonizing fungi in the host plant tissue, especially in the inner cortex of the root. These fungi have profound effects on host plants, resulting in greater water usage, enhanced resistance to drought and diseases, elevated rates of photosynthesis, and improved rates of growth under both normal and stress conditions.2, 3)
In India, large land areas have unfavorable acid and saline soil conditions.4) In the case of saline and sodic soils, drainage is poor and salt accumulates on the surface of the soil, thus adversely affecting the plant growth. The application of VAM fungi is the best simple way to improve the survival of the vegetations, because these organisms are important components to the long-term health and stability of maritime sand dunes.5) VAM in dune systems contribute to binding of sand grains into large aggregates and to improving soil structure, factors that can influence plant succession.6,7)
Because mycorrhizae (VAM) are important for the survival and growth of plants and for stabilizing sand grains,8,9) the present work was undertaken to study the occurrence of VAM fungal species in the coastal southern peninsular tip of India and to study the effect of VAM fungal species on the survival of saline-stressed vegetations using an appropriate experimental host plant present commonly in the sites examined.
Materials and Methods
Selection of study sites. Three different coastal saline soil sites with vegetations were selected. Site 1 is located in Kanyakumari, a coastal town present in the south most end of the southern peninsular tip of India. Site 2 is located in Colachel, a harbor west of site 1. Site 3 is located in Uvari, a coastal village northeast of site 1. All three sites were present in the southern peninsular tip of India. Common vegetations in all three sites were observed, and soil samples from the root- zone area and vegetation root samples were collected.
Collection of samples. All three sites had no previous occurrences of VAM fungi. Rhizosphere soils of the vegetations were analyzed for the occurrence of VAM spores.
The root samples were fixed in formalin acetic acid (FAA)10) [6 ml of formalin (40% formaldehyde), 1 ml of glacial acetic acid, 20 ml of ethanol (96%), and 40 ml of distilled water]
immediately after collection and analyzed for the incidence of VAM structures in the root tissues. In addition, physicochemical characteristics of the rhizosphere soils were analyzed.
Isolation and identification of VAM spores and inoculation studies of VAM spores in Prosopis juliflora.
VAM spores were isolated, and the dominant spores found in each site were recorded. For the isolation of VAM spores, wet- sieving and decanting technique11) was followed. The isolated VAM spores were identified by the synoptic keys of Gerdemann and Trappe,12) Raman and Mohankumar,13) Schenck and Perez,14) and Walker and Trappe.15) The dominant spores were prepared for VAM inoculum production using the pot-culture method. For inoculation, 1 g of soils with 250-500 VAM spores were used.
For inoculation studies, Prosopis juliflora, a common woody legume found in the coastal soil, common with
*Corresponding author
Phone: +82-61-750-3294; Fax: +82-61-752-8011 E-mail: [email protected]
mycorrhizal association was selected. Experimental plants were inoculated with different types of VAM fungi and harvested 75 days after inoculation for determination of the percent root colonization of VAM fungi, VAM spore number, and dry matter content.
Results and Discussion
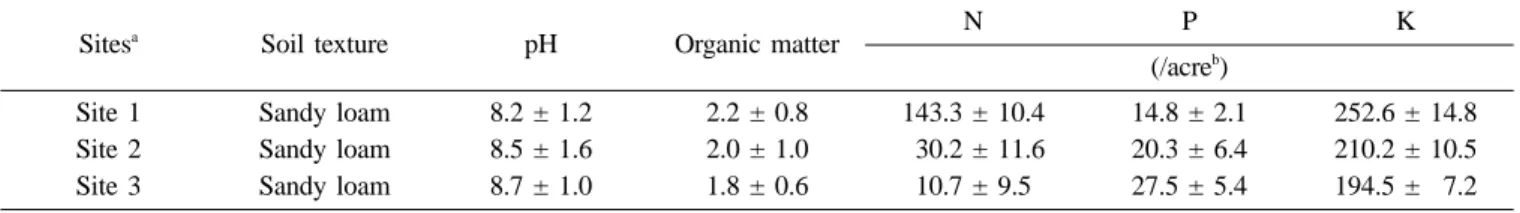
Soils of all three sites were alkaline (pH 8.2-8.7), sandy loam type. The alkaline nature of coastal soils in India has already been reported.16) Total nitrogen, available phosphorus, and available potassium of the soil ranged from 110.7 to 143.3, 14.8 to 27.5, and 194.5 to 252.6 kg/acre, respectively (Table 1). Sediments were analyzed for organic matter, N, P, and K contents. Previous report16) on fertility of Indian soil revealed that N, P, and K levels in the soil were low, thus the soil could only act as a low-fertility soil.
A total of 16 different VAM fungal species were detected in the rhizosphere soils, among which two belong to the genus Acaulospora, three to Gigaspora, two to Sclerocystis, two to Scutellospora, and the remaining seven to Glomus (Table 2).
Fifteen commonly occurring angiosperms belonging to 10 different families were selected from the selected sites and screened for VAM in the roots as well as for VAM structures in root-zone soils. All 15 plant species were positive for VAM colonization in the roots, although the type of VAM fungi colonizing the roots varied. VAM structures such as sporocarps and spores belonged to different VAM fungal species. The number of VAM fungal spores present in the root-zone soils ranged from 120 to 470, and the percent root colonization varied from 25 to 95 (Table 3).
Among the 15 plants selected for the current study, 7 were common for all three sites, and the remaining 8 were present only at specific sites. In particular, Justicia simplex, Borassus flabellifer, and Physalis minima were present only at a single site. Furthermore, 3 plants belonged to non-mycorrhizal families showing no mycorrhizal incidence, and the other 12 plants belonged to mycorrhizal families showing VAM incidence in their roots (Table 3).
All 15 plant species recorded in the three coastal sites showed mycorrhizal association. This result not only agreed with the widespread occurrences of mycorrhizae reported in natural ecosystem,17) but also revealed that mycorrhizal (VAM) fungi, due to their importance in plant growth and structure,5) may play a key role in plant protection through
their involvement in the development of methods for stabilizing the sand dunes. Trappe18) also mentioned that VAM fungi provide a green cover on the coast and improve the growth of coastal crops. Although reported as non- mycorrhizal families,19) Amarantaceae and Euphorbiaceae were revealed to be mycorrhizal in the present study, in agreement with the report of Vijayalakshmi and Rao.20) Mycorrhizal and non-mycorrhizal plant species occurring together in different sites examined was reported previously.21) At site 1, two, three, five, two, and two species of Acaulospora, Gigaspora, Glomus, Sclerocystis, and Scutellospora, respectively, were recorded. At site 2, two, three, six, one, and two species of Acaulospora, Gigaspora, Glomus, Sclerocystis, and Scutellospora, respectively, were recorded. At site 3, two, three, seven, one, and two species of Acaulospora, Gigaspora, Glomus, Sclerocystis, and Scutellospora, respectively, were recorded. Among the 16 VAM species isolated, 13 species including Acaulospora elegans, A. levis, Gigaspora albida, Gig. gigantea, Gig.
margarita, Glomus citricola, Gl. fasciculatum, Gl.
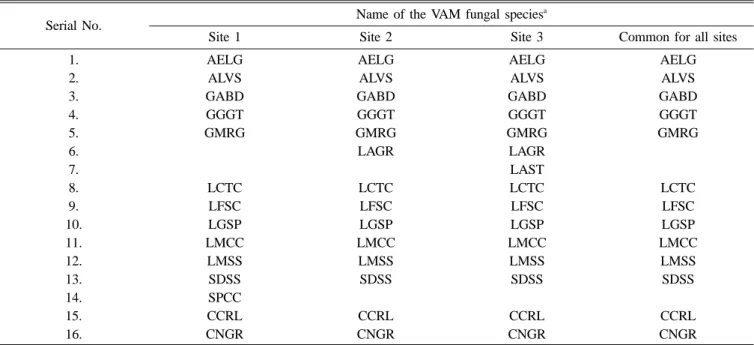
geosporum, Gl. macrocarpum, Gl. mosseae, Sclerocystis dusii, Scutellospora coralloides, and Scu. nigra were encountered in all three sites (Table 4).
Table 1. Physicochemical characteristics of soils at three different coastal sites in the southern peninsular tip of India
Sitesa Soil texture pH Organic matter N P K
(/acreb)
Site 1 Sandy loam 8.2 ± 1.2 2.2 ± 0.8 143.3 ± 10.4 14.8 ± 2.1 252.6 ± 14.8 Site 2 Sandy loam 8.5 ± 1.6 2.0 ± 1.0 030.2 ± 11.6 20.3 ± 6.4 210.2 ± 10.5 Site 3 Sandy loam 8.7 ± 1.0 1.8 ± 0.6 10.7 ± 9.5 27.5 ± 5.4 194.5 ± 07.2
aSites are described in the text.
bValues are means of 10 replicates with standard error 2%.
Table 2. VAM fungal species identified in the root-zone soils at three coastal study sites in the southern tip of the penin- sular India
Serial No. Identified VAM fungal species Abbreviation
1. Acaulospora elegans AELG
2. Acaulospora levis ALVS
3. Gigaspora albida GABD
4. Gigaspora gigantea GGGT
5. Gigaspora margarita GMRG
6. Glomus aggregatum LAGR
7. Glomus australe LAST
8. Glomus citricola LCTC
9. Glomus fasciculatum LFSC
10. Glomus geosporum LGSP
11. Glomus macrocarpum LMCC
12. Glomus mosseae LMSS
13. Sclerocystis dusii SDSS
14. Sclerocystis pachycaulis SPCC 15. Scutellospora coralloides CCRL
16. Scutellospora nigra CNGR
Glomus australe and Sclerocystis pachycaulis were found only in one site (Table 5). Achyranthus aspera and Prosopis juliflora in all sites showed highest number (6) of VAM species. Among the investigated plant species, variations in the percent root colonization and species composition of associated VAM fungi were observed. The percentage of VAM colonization was highest in Prosopis juliflora (95%) and lowest in Trianthema decandra (25%) (Table 3).
The number of spores in the rhizosphere soils at the three sites varied significantly, in agreement with the earlier findings of Rama Pulla Reddy and Manoharachary.22) This variation in spore number was due to the increased variations in the concentration of NaCl in the soil solution. Ragupathy and Mahadevan23) revealed that the VAM spore density was influenced by the salinity in the coastal line. Juniper and Abbott24) also reported that the increased NaCl concentration in the soil solution reduced the growth of VAM spores.
Occurrence of both host-specific and non-host-specific
association of VAM fungi was also revealed. Among the 15 host plant species investigated, Phyllanthus maderaspatensis and Justicia simplex showed host-specific associations of the fungi Glomus aggregatum and Gl. australe, as reported previously.19,25) Although VAM fungi are cosmopolitan in distribution, their frequency is markedly influenced by various environmental and edaphic factors.26) Both abundance and cosmopolitanism of VAM fungi and vascular plants are related to nutrients and availability of moisture in soil.27) In addition, latitudes and temperature gradient influence the occurrence of VAM fungal community.28)
Although VAM species have not been known to be strictly site-specific,29) in the present study, Glomus australe in site 3 and Sclerocystis pachycaulis in site 1 showed site-specificity.
A significant correlation between the number of spores and percent root colonization was observed in Trianthema decandra and Prosopis juliflora, as previously reported by in other plants.28,30)
Table 3. Percent root colonization, VAM spore count, and VAM species associated in the root-zone soils of three coastal sites of the southern peninsular tip of India
Serial
No. Family and plant species Sitesa Percent root colonization
VAM spore number/
100 g of soil
VAM fungal species associatedb
I NON-MYCORRHIZAL FAMILIES
AMARANTACEAE
1. Achyranthus aspera 1, 2, 3 70 240 AELG, GABD, LCTC, LMCC,
SDSS, CNGR EUPHORBIACEAE
2. Croton sparciflorus 1, 2, 3 80 320 ALVS, GGGT, LFSC, LMCC
3. Phyllanthus maderaspatensis 2, 3 60 210 GMRG, LAGR, LGSP, LMCC
II MYCORRHIZAL FAMILIES
ACANTHACEAE
4. Justicia simplex 3 45 180 GGGT, LAST, LFSC, LMCC
5. Tridax procumbens 1, 2, 3 90 410 AELG, LCTC, LMSS, CCRL
MIMOSAE
6. Acacia planifrons 1, 2 65 230 ALVS, LFSC, LMCC, CCRL
7. Prosopis juliflora 1, 2, 3 95 470 GABD, GMRG, LFSC, LMCC,
SDSS, CNGR PALMACEAE
8. Borassus flabellifer 1 70 260 GGGT, LMCC, LMSS, SPCC
9. Cocos nucifera 1, 2, 3 75 300 AELG, GMRG, LMCC, CNGR
POACEAE
10. Chloris bournei 1, 2, 3 50 185 AELG, GGGT, LMCC, LMSS
11. Cynodon dactylon 2, 3 40 170 GABD, LGSP, LMCC
PORTULACACEAE
12. Portulaca oleracea 1, 3 40 150 LAST, LCTC, LMCC
13. Trianthema decandra 2, 3 25 120 LFSC, LMCC, CCRL
RANUNCULACEAE
14. Casuarina equisetifolia 1, 2, 3 60 215 GABD, LGSP, LMCC, CNGR
SOLANACEAE
15. Physalis minima 1 45 160 LCTC, CCRL
aSites are described in the text.
bAbbreviations as in Table 2
The plants of Prosopis juliflora in the fields inoculated with selected dominant VAM spores were analyzed for percent root colonization, VAM spores, and dry matter content. Results showed that the plants inoculated with Glomus macrocarpum gave higher levels of percent root colonization (95%), VAM spores (610), and dry matter content (630 mg/plant), followed by those inoculated with Glomus fasciculatum and Scutellospora corralloides (Table 6).
Analysis of the experimental host plant Prosopis juliflora inoculated with selected VAM spores revealed that Glomus macrocarpum was the most efficient saline-tolerant VAM fungi, with maximum VAM infection in the root, and more beneficial for the plant growth. In spite of saline toxicity, VAM
fungi were efficient in tolerating the saline-stressed ecosystem. Similar results have been reported by Mandal and Kaushik31) and Mehrotra32) on VAM-treated low-fertile soils.
In addition, woody legume tree members, Prosopis juliflora in particular, are recognized as useful for revegetation of water- deficient, low-nutrient environments due to their ability to form higher percentage of symbiotic association with Rhizobium bacteria and VAM (mycorrhizal) fungi, which improves nutrient acquisition and helps plants to become established and cope with stress situation.33) Moreover, woody legumes generally exhibit a considerable degree of dependence on mycorrhizae to thrive under stressed situations.16,34-39)
Acknowledgment. Support of the Director, Centre for Advanced Studies in Botany, University of Madras is gratefully acknowledged.
References
1. Hayman, D. S. (1982) Influence of soil fertility on activity and survival of vesicular-arbuscular mycorrhizal fungi. Phy- topathol. 72, 1119-1125.
Table 4. List of VAM fungal species isolated from the soils of three coastal study sites
Serial No. Name of the VAM fungal speciesa
Site 1 Site 2 Site 3 Common for all sites
1. AELG AELG AELG AELG
2. ALVS ALVS ALVS ALVS
3. GABD GABD GABD GABD
4. GGGT GGGT GGGT GGGT
5. GMRG GMRG GMRG GMRG
6. LAGR LAGR
7. LAST
8. LCTC LCTC LCTC LCTC
9. LFSC LFSC LFSC LFSC
10. LGSP LGSP LGSP LGSP
11. LMCC LMCC LMCC LMCC
12. LMSS LMSS LMSS LMSS
13. SDSS SDSS SDSS SDSS
14. SPCC
15. CCRL CCRL CCRL CCRL
16. CNGR CNGR CNGR CNGR
aAbbreviations used as in Table 2
Table 5. Specificity of VAM fungal species from three coastal study sites
Sitea Specificityb
Site 1 SPCC
Site 2 NIL
Site 3 LAST
aSites are described in the text
bAbbreviations used as in Table 2
Table 6. Percent root colonization, VAM spore count, and dry matter content of Prosopis juliflora plant species harvested 75 days after inoculation with different VAM species
Treatment Percent root colonization VAM spores/100 g of soil Dry matter content (mg/plant)
Control 0 0 250a ± 10.6
Glomus macrocarpum 95 ± 1.5 610 ± 2.2 630a ± 12.6
Glomus fasciculatum 84 ± 2.0 540 ± 1.8 580a ± 12.0
Scutellospora coralloides 75 ± 1.8 490 ± 1.4 510a ± 11.2
aDifferences in the values are statistically significant (p = 0.5) using F-test.
2. Rosendahl, C. N. and Rosendahl, S. (1991) Influence of vesicular-arbuscular mycorrhizal fungi (Glomus spp.) on the response of cucumber (Cucumis sativus) to salt stress. Envi- ron. Exp. Bot. 31, 313-318.
3. Bethlenfalvay, G.. J. (1992) Mycorrhizae in crop productiv- ity. In Mycorrhizae in sustainable agriculture, Bethlenfal- vay, G. J. and Linderman, R. G. (eds.), Special Publication No. 54, pp. 1-27, American Society of Agronomy, Madi- son, WI, USA.
4. Tata, S. N. and Wadhawani, A. M. (eds.). (1992) Hand- book of agriculture. ICAR, New Dehli, India.
5. Koske, R. E. and Polson, W. R. (1984) Are VA-mycor- rhizae required for sand dune stabilization Bioscience 34, 420-424.
6. Nicolson, T. H. (1960) Mycorrhiza in the Gramineae. II.
Development in different habitats, particularly sand dunes.
Trans. Br. Mycol. Soc. 43, 132-145.
7. Koske, R. E., Sutton, J. C. and Sheppard, B. R. (1975) Ecology of Endogone in Lake Huron sand dunes. Can. J.
Bot. 53, 87-93.
8. Puppi, G. and Riess, S. (1987) Role and ecology of VA- mycorrhizae in sand dunes. Angew. Bot. 61, 115-126.
9. Rose, S. (1988) Above and below ground community devel- opment in a marine sand dune ecosystem. Plant Soil. 109, 215-226.
10. Phillips, J. M. and Hayman, D. S. (1970) Improved proce- dures for clearing roots and staining parasitic and vesicular- arbuscular mycorrhizal fungi for rapid assessment of infec- tion. Trans. Br. Mycol. Soc. 55, 158-161.
11. Gerdemann, I. W. and Nicolson, T. N. (1963) Spores of mycorrhizal Endogone species extracted by wet-sieving and decanting. Trans. Br. Mycol. Soc. 46, 235-244.
12. Gerdemann, I. W. and Trappe, J. M. (1974) The endogo- naceae in the Pacific Northwest. Mycol. Memoir. 5, 1-76.
13. Raman, N. and Mohankumar, V. (1988) Techniques in Myc- orrhizal Research, p. 279, University of Madras, Sivakami Publications, Madras, India.
14. Schenck, N. C. and Perez, Y. (1987) Manual for the identi- fication of VA-mycorrhizal fungi, p. 245, University of Florida, Synergistic Publications, Gainesville, FL, USA.
15. Walker, C. and Trappe, J. M. (1993) Names and epithets in the Glomales and Endogonales. Mycol. Res. 93, 339-344.
16. Selvaraj, T. (1998) Studies on mycorrhizal and rhizobial symbioses on tolerance of tannery effluent treated Prosopis juliflora, Ph.D. Thesis, University of Madras, Chennai, India.
17. St. John, T. V. and Coleman, D. C. (1983) The role of myc- orrhizae in plant ecology. Can. J. Bot. 61, 1005-1014.
18. Trappe, J. M. (1989) In Mycorrhizae for Green Asia, Mahadevan, A., Raman, N. and Natarajan, K (eds.), Pro- ceedings for first Asian conference on mycorrhizae, Univer- sity of Madras, pp. 347-349, Alamu Printing works, Madras, India.
19. Selvaraj, T. (1989) Studies on vesicular-arbuscular mycor- rhizae of some crop and medicinal plants, Ph.D. Thesis, Bharathidasan University, Trichy, India.
20. Vijayalakshmi, M. and Rao, A. S. (1988) Vesicular-arbuscu- lar mycorrhizal association of some Asteraceae and Ama- rantaceae. Acta. Bot. Indica. 16, 168-174.
21. Khan, A. G. (1974) The occurrence of mycorrhizae in halo- phytes, hydrophytes and xerophytes and of Endogone spores in adjacent soils. J. Gen. Microbiol. 81, 7-14.
22. Rama Pulla Reddy, P. and Manoharachary, C. (1990) Occur- rence and distribution of vesicular-arbuscular mycorrhizal fungi in soils polluted with sewage and industrial effluents.
In Mycorrhizal symbiosis and plant growth, pp. 26-27, Pro- ceedings of 2nd National Conference on Mycorrhiza, Banga- lore, India.
23. Ragupathy, S. and Mahadevan, A. (1991) Ecology of vesic- ular-arbuscular mycorrhizal (VAM) fungi in a coastal tropi- cal forest. Indian J. Microbial Ecol. 2, 1-9.
24. Juniper, S. and Abbott, L. K. (1991) The effect of salinity on spore germination and hyphal extension of some VA- mycorrhizal fungi. Abstracts of the 3rd European Sympo- sium on Mycorrhizas, University of Sheffield, Sheffield, U.K.
25. Bloss, H. E. and Walker, C. (1987) Some endogonaceous fungi of the Santa Catalina mountains in Arizona. Mycolo- gia 79, 649-654.
26. Slankis, V. (1974) Soil factors influencing formations of mycorrhizae. Ann. Rev. Phytopathol. 12, 437-457.
27. Anderson, R. C., Liberta, A. E. and Dickman, I. A. (1984) Interaction of vascular plants and vesicular-arbuscular myc- orrhizal fungi across soil moisture-nutrient gradient. Oecolo- gia (Berl.). 64, 111-117.
28. Koske, R. E. (1987) Distribution of VA-mycorrhizal fungi along latitudinal temperature gradient. Mycologia 79, 55-68.
29. Mosse, B. (1973) Advances in the study of vesicular-arbus- cular mycorrhiza. Ann. Rev. Phytopathol. 11, 171-196.
30. Walker, C., Mize, W. and Mc Nabb, H. S. (1982) Popula- tion of endogonaceous fungi at two location in central Iowa.
Can. J. Bot. 60, 2518-2529.
31. Mandal, B. S. and Kaushik, J. C. (1995) Interaction between VA-mycorrhizal fungi and Rhizobium and their effect on the growth parameters of Acacia nilotica. Hary- ana Univ. J. Res. 25, 107-111.
32. Mehrotra, M. D. (1995) A practical approach to mycor- rhizal containerised seedlings in forest nurseries. Indian For- ester 121, 670-671.
33. Herrera, M. A., Salamanca, C. P. and Barea, J. M. (1993) Inoculation of woody legumes with selected arbuscular mycorrhizal fungi and rhizobia to recover desertified medi- terranean ecosystems. Appl. Environ. Microbiol. 59, 129- 133.
34. Carpenter, A. T. and Allen, M. F. (1988) Responses of Hedysarum borelae Nutt. to mycorrhizas and Rhizobium:
plant and soil nutrient changes in a disturbed shrub-steppe.
New Phytol. 109, 125-132.
35. Habte, M., Fox, R. L., Aziz, T. and El-Swaify, S. A. (1988) Interaction of vesicular-arbuscular mycorrhizal fungi with erosion in an oxisol. Appl. Environ. Microbiol. 54, 945-950.
36. Jasper, D. A., Abbott, L. K. and Robson, A. D. (1989) Aca-
cias respond to additions of phosphorus and to inoculation with VA mycorrhizal fungi in soils stockpiled during min- eral sand mining. Plant Soil. 115, 99-108.
37. Osonubi, O., Mulongoy, K., Awotoye, O. O., Atayese, M.
O. and Okali, D. U. U. (1991) Effects of ectomycorrhizal and vesicular-arbuscular mycorrhizal fungi on drought toler- ance of four leguminous woody seedlings. Plant Soil. 136, 131-143.
38. Roskoski, J. P., Pepper, I. and Pardo, E. (1986) Inoculation of leguminous trees with rhizobia and VA-mycorrhizal fungi. For. Ecol. Manage. 16, 57-68.
39. Sylvia, E. M. (1990) Inoculation of native woody plants with vesicular-arbuscular mycorrhizal fungi for phosphate mine land reclamation. Agric. Ecosyst. Environ. 31, 253- 261.