J. of Korean Orthopaedic Research Society Volume 5, Number 2, October, 2002
인체의 nebulin SH3 domain과 결합하는 단백질에 관한 연구
인제대학교 서울백병원 정형외과학교실 최수호・고한석・김덕원
= Abstract =
A Study on the Proteins that Interact with Human Nebulin SH3 Domain
Soo Ho Choi, M.D., Han Suk Ko, M.D., Deok Weon Kim, M.D.
Department of Orthopedic Surgery Seoul Paik Hospital, Inje University, Korea
Objective: By identifying the unknown substance responsible for binding with nebulin SH3 domain within the sarcomeric Z-line, we tried to find out Z-line structure which plays an important role on muscle contraction and maintenance of muscle funtion.
Method: First, the bait plasmid was made by binding the DNA binding domain of Gal4 protein of yeast and the SH3 domain. Second, library plasmid was made by binding activation domain and human skeletal cDNA library. Then, the base sequence of the clone, produced by combining the two proteins expressed by transgeni- cally converted plasmid in yeast, was analyzed.
Result: We screened out six true positive clones and analyzed the base sequence of the two of six clones.
We identified them to be alpha-actinin2.
Conclusion: We can theorize that Neublin SH3 domain and alpha-actinin2 plays a vital role for the integra- tion of Z-line. Thus, this is an important data in further studying muscle functions, mechanisms, and muscular disease as well.
Key Words: Nebulin, SH3 domain, Yeast two-hybrid, α-actinin 2
※ 통신저자: 김 덕 원
서울시 중구 저동 2가 8 5번지 서울백병원 정형외과
Tel: 02) 2270-0028, Fax: 02) 2270-0028 E-mail: [email protected]
서 론
최근 세포내의 신호전달체계에 있어서 단백질들 간의 상호작용에 대해 많은 연구가 이루어져왔다.
Src 암유전자에서 처음으로 발견된 S H 2와 S H 3 d o m a i n은 단백질들간의 상호작용을 조절하는데 있어서 중요한 역할을 수행한다1 4 ). 일반적으로 SH3 domain은 신호전달과정에서 d o w n s t r e a m 의 단백질과 특이적으로 상호작용하고, SH2 d o m a i n은 u p s t r e a m의 p h o s p h o t y r o s i n e에 나 타나는 특정 서열 인식 부위를 통해 t y r o s i n e - phosphorylated protein과 상호작용하는 것으 로 알려졌다. SH3 domain을 통한 신호전달은 단백질 k i n a s e와 같은 효소의 연쇄적 반응을 통 해 일어나거나 세포골격 결합구조의 변화로 일어 난다.
N e b u l i n의 C말단은 근절의 Z - l i n e안에 고정되 어 있고, SH3 domain을 포함하며 근원섬유 단 백질에서 처음으로 이 d o m a i n이 인식되었다
4 , 1 2 , 1 7 )
. Z-line의 결합과 thin filament의 형성에 서 n e b u l i n의 역할을 설명하기 위한 노력들이 이
루어졌고4 , 1 2 , 1 7 ) a c t i n1 7 ), titin4 ), 그리고 C a p - Z
p r o t e i n1 2 )이 SH3 domain의 결합단백질로 추정 되었다.
Wang 등1 8 )은 SH3 domain이 a c t i n과 결합하 여 Z - l i n e에 nebulin filament를 고정시키는데 관여할 것으로 추측하였고 Politon 등1 1 )은 p r o- line-rich 유사서열을 가지는 t i t i n을 지목하였 다. 그러나 nebulin SH3 binding partner로 확신을 줄 수 있는 결정적인 증거는 없다.
본 연구는 단백질간의 직접적 결합에 의해 nebulin SH3 domain과 결합하는 단백질을 찾 아 Z - l i n e의 구조를 밝힘으로써 근육수축의 기전 을 알아내어 근육의 기능유지 및 향상에 도움이 될 수 있게하고, 근육질환 연구에 기초자료를 제 공 하려고 하였다.
연구재료 및 방법
1. Bait plasmid의 제작과 t w o - h y b r i d s c r e e n i n g을 위한 준비
Nebulin SH3 domain을 증폭하기 위한 primer 쌍은 이 d o m a i n의 3'말단으로부터 4개 의 염기를 제외시키고 d o m a i n의 3배 정도의 P o s t - R (반복구간 이후 지역)를 포함하도록 제작 하였다.
사람 골격근(대퇴직근 0.5 g)으로부터 t o t a l R N A를 추출하고 nebulin SH3 domain에 특이 적인 H N eⅡ p r i m e r를 이용하여 c D N A를 합성 하였다. 제작된 c D N A를 주형으로 하여 S H 3 domain 전후의 H N eⅠ, HNeⅡ p r i m e r로 P C R을 수행하여 613 bp의 PCR 생성물을 얻은 후, 이것을 효모 two-hybrid screening을 위한 bait plasmid의 제작에 사용하였다.
효모 two-hybrid system을 위한 bait plas- m i d의 제작을 위해 Gal4 DNA-binding d o m a i n만을 가지는 p G B T 9이 이용되었다.
PCR 생성물을 먼저 pBluescript II SK(+)에 삽 입하여 E. coli에서 증폭시킨 후 제한효소 B a m HⅠ과 S a lⅠ으로 처리하고 절단된 i n s e r t를 p G B T 9에 다시 삽입하였다. SH3 domain이 삽 입된 vector, pGBT9-SH3-2, -9, -45를 선별하 였고 H N eⅠ, HNeⅡ로 다시 P C R하여 r e c o m- binant clone을 확인하였다. 이러한 과정을 거친 c l o n e은 염기서열 분석으로 open reading f r a m e을 확인하여 bait plasmid로 사용하였다.
효모의 two-hybrid screening에서 G a l 4의 activation domain 없이 Gal4 DNA-binding d o m a i n이 단독으로, 혹은 bait 단백질이 융합된 Gal4 DNA-binding domain이 단독으로 β- galactosidase 활성을 보인다면 그 bait plas- m i d는 이 s y s t e m에 사용할 수가 없다. 이에 따 라 Gal4 activation domain이 없는 조건에서도 β-galactosidase 활성이 나타나는지의 여부를 확 인하기 위하여 p G B T 9과 p G B T 9 - S H 3는 각각 홀로 H F 7 c에 형질 전환되었고 선발 배지인 SD/-Trp 에서 배양하였다. 이 때 p G B T 9은 선 발 m a r k e r로서 T r p 1을 가지고 m u t a t i o n된 HF7c 내에서 아미노산인 t r y p t o p h a n을 합성하 므로 상기조건에서 생장할 수 있다. 실험 결과 SD/-Trp 배지에서 생장한 형질전환체인 p G B T 9 과 p G B T 9 - S H 3는 colony-lift filter 분석을 통
하여 β-galactosidase 활성을 보이지 않았으므로 negative control로 사용되었다.
p C L 1은 G a l 4의 완전한 염기서열을 가지므로 β-galactosidase 활성의 positive control로 사 용하였다. 이것을 확인하기 위하여 p C L 1을 H F 7 c에 형질전환시킨 후, 선발 배지인 l e u c i n e 이 제외된 S D / - L e u에 배양한 후 c o l o n y - l i f t filter 분석한 결과 blue colony가 나타남을 확인 하였다. 또 다른 positive control로서 이미 결합 성이 알려진 두 단백질의 염기서열이 각각 삽입된 p V A 3와 p T D 1으로 H F 7 c를 동시에 형질전환시 키므로써 β-galactosidase 활성의 positive con- t r o l로 사용하였다. pVA3는 G a l 4의 D N A - binding domain이 융합된 쥐과의 p53 단백질을 암호화하며 p T D 1은 G a l 4의 a c t i v a t i o n d o m a i n이 융합된 SV40 large T-antigen을 암 호화한다. 두 단백질은 효모 내에서 동시에 발현 되어 His3 유전자와 L a c Z의 유전자를 발현시키 므로 β-galactosidase 분석 시 blue colony로 나타난다. 이러한 control 실험의 예비적 결과들 을 기초로 하여 two-hybrid screening의 본 실 험을 개시하였다.
2. 효모 two-hybrid screening test를 통한 N e b u- lin SH3 domain과 결합하는 단백질의 발견
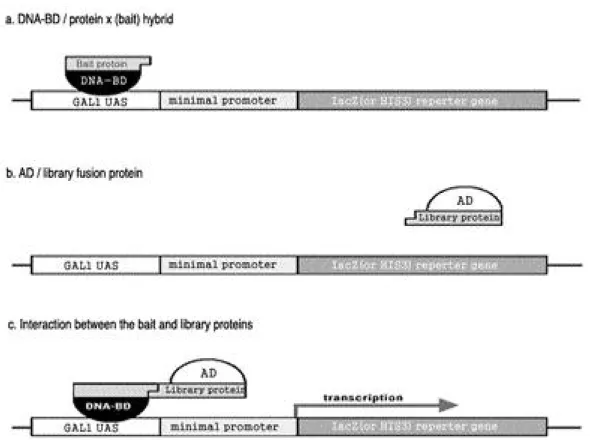
효모 two-hybrid system은 세포 내에 존재하 는 두 단백질간의 직접적 결합 여부를 조사하는 방법이다. 효모 세포에서 전사의 활성자인 G a l 4 단백질은 각각 독립적으로 기능하는 D N A - b i n d- ing domain과 activation domain으로 구성되 어 있다. 이 두 d o m a i n이 분리되어있는 경우에 는 G a l 4의 활성이 나타나지 않으나, 두 d o m a i n 의 위치가 근접한 경우에는 G a l 4의 활성이 나타 난다. Gal4 단백질의 DNA-binding domain에 융합된 bait 단백질과 activation domain에 융 합된 library 단백질이 결합된 경우에 한하여 Gal4 단백질은 활성화되고 이에 따라 r e p o r t e r 유전자인 H i s 3와 LacZ gene의 발현이 이루어진 다(Fig. 1).
효모의 two-hybrid screening을 위해서는 bait 단백질을 Gal4 DNA-binding domain에
융합시킨 bait plasmid와 cDNA library를 Gal4 activation domain에 융합시킨 l i b r a r y p l a s m i d를 준비하고 이들을 동시에 효모에 형질 전환 시킨다. 만약 형질전환된 효모내의 두 p l a s- m i d로부터 발현된 bait 단백질과 library 단백질 이 결합한다면 이 효모들은 reporter 유전자인 His3 유전자를 발현시키므로 histidine 결핍 선 발배지에서 자랄 수 있다. 그러므로 선발배지에서 생장한 c o l o n y들은 1차 선발된 것들이며 2차 선 발을 위하여 다시 β-galactosidase 분석하였다.
이 분석에 의하여 LacZ 유전자를 발현시키는 blue colony를 선발한다. 두 차례에 걸친 선발과 정을 통하여 많은 위양성 c o l o n y들이 제거된다.
결 과
이상의 control 실험에 준거하여 효모의 H F 7 c 균주 내에서 단독으로 β-galactosidase 활성을 나타내지 않는 p G B T 9 - S H 3를 그 b a i t로 이용하 여 사람 골격근 cDNA library에 대한 검색을 시도하였다.
효모의 Gal4 activation domain을 발현시키 는 vector 즉, pGAD10에 삽입된 사람 골격근 cDNA library 및 bait plasmid인 p G B T 9 - S H 3로 H F 7 c를 동시에 형질전환 시킨 후 효모세 포 내에서 각각의 융합 단백질을 동시에 발현시키 고 이들 단백질의 결합여부를 확인하고자 하였다.
많은 형질전환체의 c o l o n y들이 S D / - T r p / - Leu/-His/+3-AT 배지 위에서 생장하였다. 선발 배지에서 생장한 이 c o l o n y들은 p G B T 9에 의해 t r y p t o p h a n을, 그리고 p G A D 1 0에 의해 l e u c i n 을 합성할 뿐 아니라 각각의 p l a s m i d로부터 발현 되는 두 융합단백질의 결합에 의해 reporter 유 전자인 H i s 3를 발현시킴으로서 h i s t i d i n e을 합성 한다.
SD/-Trp/-Leu/-His/+3-AT 배지 위의 형질전 환체 중 대부분은 colony-lift filter 분석 시 β- galactosidase 활성을 보이는 6개의 양성 c l o n e 를 선택하였는데 그 중 두 c l o n e은 염기서열 분석 으로 α-actinin 2임을 확인하였다.
이들의 위양성 여부를 검정하기 위해 G a l 4 activation domain과 α-actinin 2의 융합단백
질을 암호하는 p l a s m i d와 Gal4 DNA-binding d o m a i n과 nebulin SH3 domain의 융합단백질 을 암호하는 p l a s m i d를 H F 7 c로 함께 형질전환 하였을 때 그 형질전환체는 S D / - T r p / - L e u / - His/+3-AT 배지에서 강한 성장을 보였으며 β- galactosidase 분석에서 청색을 보이는 양성반응 을 확인하였다. Gal4 activation domain과 α- actinin 2의 융합단백질과 Gal4 DNA-binding d o m a i n만이 함께 존재하는 형질전환체는 h i s t i- d i n e이 포함된 SD/-Leu/-Trp 배지에서 생장시 킨 후 β-galactosidase 활성을 분석하여 양성반 응을 나타내지 않음을 확인하였다.
구입한 library plasmid는 p G A D 1 0의 E c o R
Ⅰ 지역에 i n s e r t가 삽입된 것이었으므로 염기서 열 분석 후 α-actinin 2를 암호하는 서열을 포함 하는 두 p l a s m i d의 insert 크기를 알고자 E c o R
Ⅰ처리하였다. 제한효소로 절단한 결과 6.6 kb인 p G A D 1 0의 b a n d를 포함하여 약 2 kb의 b a n d 가 agarose 전기영동 상에서 관찰되었다.
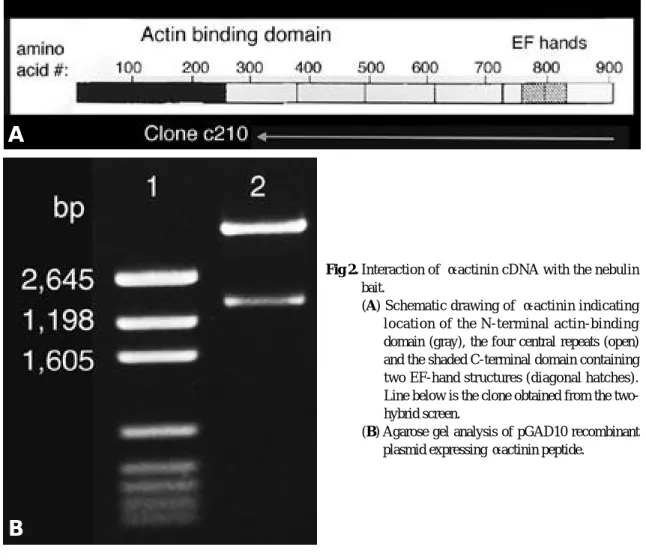
pGAD10 vector에 삽입된 DNA 단편의 총 길 이는 약 2 kb로 예상된다. 약 700 bp의 염기서 열을 분석하였고 그 시작점으로부터 C -말단으로 2 kb의 예상되는 삽입 DNA 단편을 그림으로 나타내었다(Fig. 2). 독립된 두 c l o n e으로부터 얻은 두 p l a s m i d에 삽입된 DNA 절편의 총 길이 와 α-actinin 2에서 시작되는 염기서열도 같았다.
고 찰
N e b u l i n은 척추동물의 골격근에만 존재하는 것으로 알려져 있으며 액틴과 결합하는 거대 단백 질로서 전체 근원섬유의 2 ~ 3 %를 차지한다1 9 ). 단
Fig. 1. Schematic diagram of the MATCHMAKER Gal4-based two-hybrid systems(Clontech). The DNA-BD is a DNA-binding domain that corresponds to amino acid 1-147 of the yeast Gal4 protein, which binds to the Gal1 UAS upstream of the reporter gene. The lacZ and His3 reporter genes are separate constructs integrated in the yeast genome. The AD is an activation domain that corresponds to amino acid 768-881 of the Gal4 protein and has transcriptional activation function.
일 nebulin 분자는 thin filament의 길이 전체 에 걸쳐있고 근육이 수축, 이완될 때 함께 움직인 다. Nebulin은 조직과 분화단계에 따라 600~900 kDa의 다양한 크기로 존재하며, 분화 가 이루어지는 동안 thin filament의 길이를 조 절하는 기준자로 작용한다1 ).
N e b u l i n은 super repeat, simple repeat, SH3 domain 3부분으로 구성되는데 3 5 - r e s i d u e m o d u l e이 1 8 5번 반복되는 super repeat 부분이 사람 nebulin 염기서열의 9 7 %를 차지한다6 , 1 8 ). SH3 domain은 근절의 Z - l i n e에 있는 n e b u l i n 의 C -말단에 나타난다.
N e b u l i n의 SH3 domain은 5 0여 개의 아미노 산으로 이루어져 있으며 효모에서 포유류에 이르 기까지 다양한 종에서 보존되어 있다2 0 ). SH3
d o m a i n을 가지고 있는 단백질은 S r c와 p h o s- pholipase C-γ, phosphatidylinositol 3- k i n a s e의 s u b u n i t인 p 8 0과 같이 효소와 연관된 것과 세포골격의 작용에 관여하는 m y o s i n , tensin, contactin, spectrin, vinculin, 효모 의 액틴 결합 단백질인 ABP1, chick src sub- s t r a t e인 p80/85 등이 알려져 있다2 ). SH3 d o m a i n과 결합하는 단백질로는 세포골격인 p a x i l l i n과 3BP1, 3BP2, guanine nucleotide exchange protein (SOS), Ras GTPase-acti- vating protein (GAP), marcrophage에서 c - c b 1이 알려졌으며 이들 단백질들은 p r o l i n e - r i c h m o t i f를 가지고 있어 이 m o t i f가 SH3 domain 의 결합 부위로 작용한다고 밝혀졌다3 , 5 , 1 0 ). 특히 이들 단백질 가운데 상당수는 focal adhesion에
Fig 2. Interaction of α-actinin cDNA with the nebulin bait.
(A) Schematic drawing of α-actinin indicating location of the N-terminal actin-binding domain (gray), the four central repeats (open) and the shaded C-terminal domain containing two EF-hand structures (diagonal hatches).
Line below is the clone obtained from the two- hybrid screen.
(B) Agarose gel analysis of pGAD10 recombinant plasmid expressing α-actinin peptide.
A
B
관여하는 단백질들이다.
본 연구에서는 사람 nebulin SH3 domain과 결합하는 단백질을 찾기 위해 yeast two-hybrid s y s t e m을 이용하였으며, 이를 위해 Gal4 DNA- binding domain에 SH3 domain을 융합시킨 p l a s m i d와 Gal4 activation domain에 사람 골 격근 cDNA library가 융합된 p l a s m i d를 각각 b a i t와 f i s h로 사용하였다. 염기서열 분석을 통해 bait plasmid의 open reading frame을 확인하 였다. Nebulin SH3 domain이 융합된 G a l 4 - binding domain이 단독으로 존재할 때는 reporter 유전자에 대한 전사활성 능력을 보이지 않음을 확인한 후 이 실험의 bait plasmid로 사 용하였다. Nebulin의 SH3 domain과 결합 반응 을 보인 c D N A를 염기서열 분석하고 상동성을 검 색한 결과, 이 c D N A는 α-actinin 2와 100% 일 치함을 알 수 있었다.
α-actinin2 는 골격근에 존재하는 α- a c t i n i n으 로 c a p Z와 함께 Z - l i n e을 구성하는 대표적인 단
백질이다1 6 ). 서로 등을 맞대는 actin filament의
중첩으로 생기는 Z - l i n e의 폭은 Z filament의 반복층에 의해 결정되며 어류의 white muscle은 1층7 ), rat soleus muscle은 3 ~ 4층1 3 ), nema- line myopathy 환자에서는 1 0여 층으로 나타난
다1 6 ). Z-line의 폭은 fast twitch fiber에서는
좁고 slow twitch fiber에서는 넓게 나타나서 근 육의 수축 속도와 반비례하므로, Z-line의 폭을 조절하는 것은 생리학적으로 대단히 중요하며 nemaline myopathy의 경우에서 보듯이 병리학 적으로도 대단히 중요하다. 평행한 actin fila- m e n t를 엮어 격자층을 만드는 Z filament는 α- a c t i n i n으로 알려져 있다1 6 ). 그러나 격자층을 만 드는 Z filament 간의 간격이 18 nm의 이론치
와1 3 ) 36~38 nm의 측정치8 ) 간에 차이를 보이므
로써 Z - l i n e의 구조가 단순히 a c t i n과 α- a c t i n i n 으로 이루어지는 것이 아니고, 다른 단백질의 결 합이 Z - l i n e의 두께를 결정할 것으로 보인다.
Sorimachi 등은1 5 ) Z - l i n e까지 뻗어있는 t i t i n의 N 말단이 α- a c t i n i n과 직접 결합하여 Z fila- ment 층의 수를 결정할 것이라는 제안을 하였 다. 한편으로 Nave 등9 )은 n e b u l i n의 C 말단이 α- a c t i n i n과 결합하여, α- a c t i n i n의 a c t i n과의
결합을 조절할 수 있을 것이라는 추정을 하였다.
결 론
Nebulin SH3 domain이 α-actinin 과 결합 함을 처음으로 밝혔다. 이는 α- a c t i n i n은 근육외 에도 여러 조직에서 세포와 세포간의 결합과 세포 와 기질간의 결합을 조절하고, actin구조의 조절 에 관여하므로 이들 조절 기능을 연구하는데 중요 한 정보가 될것이고, 근섬유의 결합과 분리를 조 절하는 것으로 추정되어 근육의 기능유지 및 향상 과 근질환의 연구에 도움이 될 것이다.
R E F E R E N C E S
01 ) Chen MJ, Shih CL, Wang K: Nebulin as an actin zipper: A two-module nebulin fragment promotes actin nucleation and stabilizes actin filaments. J Biol C h e m, 268: 20327-20334, 1993.
02 ) Drubin DG, Muholand, J, Zhu Z, Botsein D:
Homology of a yeast actin-binding protein to signal transduction proteins and myosin-I. N a t u r e, 343:
288-290, 1990.
0 3 ) Egan SE, Giddings BW, Brooks MW, Buday L, Sizeland AM, Weinberg RA: Association of SOS ras exchange protein with Grb2 is implicated in tyrosin kinase signal transduction and transforma- tion. Nature, 363: 45-51, 1993.
04 ) Gautal M, Goulding D, Bullard B, Weber K, Furst DO: The central Z-line region of titin is assembled from a novel repeat in variable copy numbers. J Cell Sci, 109: 2747-2754, 1996.
05 ) Knudsen BS, Zheng J, Feller SM: Affinity and specificity requirements the first Src homology 3 domain of the Crk proteins. EMBO J, 10: 2191- 2199, 1995.
06 ) Labeit S, Kolmerer B: The complete primary structure of human nebulin and its correlation to muscle structure. J Mol Biol, 248: 308-315, 1995.
07 ) Luther PK: Three-dimensional reconstruction of a simple Z-band in fish muscle. J Cell Biol, 113:
1043-1055, 1991.
08 ) Meyer RK, Aebi U: Bundling of actin filaments by α-actinin depends on it molecular length. J Cell Biol, 110: 2013-2024, 1990.
09 ) Nave R, Furst DO, Weber K: Interaction of α- actinin and nebulin in vitro: Support for the exis- tence of a fourth filament system in skeletal muscle.
FEBS Lett, 269: 163-166, 1990.
1 0 ) Odai H, Sasaki K, Iwamatsu A, Hanazono T, Mitani T, Mitani K, et al: The proto-oncogene product c-Cb1 becomes tyrosine phosphorylated by stimulation with GM-CSF or Epo and constitutively binds to the SH3 domain of Grb2/Ash in human hematopoietic cells. J Bio Chem, 270: 10800-10805, 1 9 9 5 .
1 1 ) Politou AS, Millevoi S, Gautel M, Kolmerer B, Pastore A: SH3 in muscles: Solution structure of the SH3 domain from nebulin. J Mol Biol, 276:
189-202, 1998.
1 2 ) Schafer DA, Hug C, Cooper JA: Inhibition of CapZ during myofibrillogenesis alters assembly of actin filaments. J Cell Biol, 128: 61-70, 1995.
1 3 ) Schroeter JP, Bretaudiere J-P, Sass RL, JP, Sass RL, Goldstein MA: Three-dimensional structure of the Z band in normal mammalian skeletal muscle. J Cell Biol, 133: 571-583, 1996.
1 4 ) Sicheri F, Kuriyan J: Structures of src-family tyro- sine kinase. Curr opin strucr Biol, 7(6): 777-785, 1 9 9 7 .
1 5 ) Sorimachi H, Freiburg A, Kolmerer B, Ishiura S, Stier G, Gregorio CC, et al: Tissue-specific expression and α-actinin binding properties of the Z- disc titin: Implications for the nature of vertebrate Z- discs. J Mol Biol, 270: 688-695, 1997.
1 6 ) Vigoreaux JO: The muscle Z band: Lessons in stress management. J Muscle Res Cell Motil, 1 5 : 237-255, 1994.
1 7 ) Wang K, Knipfer M, Huang QQ, van Heerden A, hus LC, Futierrez et al: Human skeletal muscle nebulin sequence encodes a blueprint for thin fila- ment architecture. Sequence motifs and affinity pro- files of tandem repeats and terminal SH3. J Biol C h e m , 271: 4304-4314, 1996.
1 8 ) Wang K, Knipfer M, Huang QQ, van Heerden A, Hsu LC, Gutierrez G, et al: Human skeletal muscle nebulin sequence encodes a blueprint for thin filament architecture. Sequence motifs and affinity profiles of tandem repeats and terminal SH3. J Biol Chem, 271: 4304-4314, 1996.
1 9 ) Wang K, Williamson C: Identification of a N2 line protein of striated muscle. Proc Natl Acad Sci USA, 77: 3254-3258, 1980 .
2 0 ) Weng Z, Thomas SM, Rickles RJ, Taylor JA, Brauer AW, Seidel-Dugan C, et al: Identification of Src, Fyn, and Lyn SH3-binding proteins: Implica- tions for a function of SH3 domains. Mol Cell Biol, 14: 4509-4521, 1994.