J. Fish Pathol., 22(3)
:239 ~ 251 (2009)
239
배양 조건이 Photobacterium damselae subsp. damselae의 미생물학적 성상 및 병원성에 미치는 영향
권문경�∙조병열�∙박명애
국립수산과학원 병리연구과, �국립수산물품질검사원 인천지원
Effects of the microbiological properties and pathogenicity of Photobacterium damselae subsp. damselae
under different culture conditions
Mun-Gyeong Kwon
�, Byoung Youl Cho
�and Myeong-ae Park Pathology Division, National Fisheries Research Institute
�
Incheon Branch Office, National Fisheries Products Inspection Service
The effects of microbiological properties and pathogenicity of Photobacterium damselae subsp. damselae were investigated under different culture conditions, temperature, pH, NaCl and iron concentration on cul- ture media. Favorable conditions for bacterial growth were between 15~30℃, pH 5~9, 0~4% NaCl concen- tration and iron contents of over 10 mM, whereas the bacterial growth was inhibited under iron chelator existence. When P. damselae was cultured in iron-limited tryptic soy broth, total protein concentration of extracellular products, cytotoxic ability of ECPs on cell line, bacterial viability in flounder serum, phospho- lipase and siderophore activities of ECPs were significantly increased. On the other hand, the activities of P.
damselae cultured under iron-added conditions were decreased.
In this study, the iron-limited conditions were similar to the host in which iron concentration is low. Dur- ing infection caused by P. damselae, the conditions could be related to the pathogenesis of the pathogen.
Key words: Photobacterium damselae subsp. damselae, Culture conditions Pathogenicity, Flounder,
Immunogenicity
세균은 환경으로부터 영양분을 흡수하고 대사 부산물을 배출하는 상호 작용을 통하여 성장하 고 증식한다. 또한 세포 표면은 일정하지 않아 감염될 때 숙주 체내의 환경과 배양 조건의 변 화에 따라 달라지며(Brown et al., 1988), 이러한 변화는 포유류(Davies et al., 1994)나 어류(Gar- duño et al., 1993a, b)에 대한 병원성 요인과 항원 성에도 영향을 미치는 것으로 알려져 있다.
Photobacterium damselae subsp. damselae (P.
damselae)는 Truper and De'Clari (1997)에 의해
Vibrio damsela에서 재분류된 균으로, damselfish, Chromis punctipinnis에서 처음으로 보고되었으
며(Love et al., 1981), 청백돌고래, Tursiops trun-catus(Fujioka et al., 1988),
방어, Seriola quin-queradiata(Sakata et al., 1989),
감성돔, Sparusaurata(Vera et al., 1991)
및 터봇, Scophthalmusmaximus(Fouz et al., 1992)에서 질병을 일으켰다
는 보고가 있으며, 우리나라에서는 동해안 지역 의 넙치 양식장에서 많은 피해를 일으킨다고 보 고되었다 (권 등, 2005). 아직까지 P. damselae의�Corresponding Author : Mun-Gyeong Kwon, Tel : 051-720-2476 Fax : 051-720-2498, E-mail : [email protected]
6...
배양 조건 변화에 따른 병원 미생물학적 성상에 대하여 연구된 바가 없으나, 어류의 감염과 연 관시켜 생각해보면, 배양 조건에 따른 phenotype 의 변화가 어체에 감염 시 일어나는 균체의 성 상 변화와 관련이 있을 것으로 추정되며, 이러 한 변화가 어체의 방어 기전에도 영향을 미칠 수 있을 것으로 추정된다.
따라서 본 연구에서는 배양 조건에 따른 P.
damselae의 미생물학적 특성과 숙주 면역반응
에 대한 균체의 저항성에 대하여 조사하였다.재료 및 방법
실험 균주
P. damselae는 동해안 넙치 양식장에서 분리되
어 상법에 따라 생화학적 성상과 16s rRNA ho- mology를 동정 후 -70℃에 보존된 것을 사용하 였다.배양 조건에 따른 크기와 형태 변화
배양 조건에 따른 P. damselae 특성은 tryptic soy broth 배지를 기본 배지로 하여 24시간 진탕 배양하면서, 온도(10~30℃), pH(3~11), 염분 (0~10%), iron-chelator제인 2,2'-dipyridyl(DIP) (0~400 μM) 및 iron-added인 FeCl3(10~200 mM) 등의 조건을 달리하여 배양 성상의 차이를 조사 하였으며, 대조구는 염분농도 1%, pH 7의 TSB 배지에 접종 후 25℃에서 배양하였다.
전자현미경적 관찰
배양 조건에 따른 P. damselae의 형태 및 크기 에 미치는 영향을 조사하기 위하여 pH 9, NaCl 4%, DIP 100 μM, FeCl3100 mM 조건에서 배양 후 전자현미경을 이용하여 관찰하였다. 즉, 각 조건의 TSB 배지에서 24시간 배양된 균액을 8,000 rpm, 10분간 원심 분리 후 멸균 PBS (pH 7.2)로 3회 세척하여 준비하였다. 준비된 균액은 4℃, 2.5% glutaraldehyde 고정액으로 고정 후 상
법에 따라 uranyl acetate and lead citrate 중염색을 적용하여 투과전자현미경(JEM 2010, JEOL LTD) 으로 관찰하였다.
균성장 및 Extracellular products (ECPs) 생성능 각 배양 조건에서 P. damselae의 균수변화는 배양 후 24시간째에 흡광도 (600 nm)를 측정하 였다. 배양된 균액은 4℃, 8,000rpm, 10분간 원심 분리하여 분리된 상징액은 0.45 ㎛ membrane에 filteration하여 ECPs로 사용하였으며, total pro- tein 농도는 Biuret 법으로 측정(Asan pharm, Co., LTD)하였다.
ECPs의 효소 활성 및 용혈능
배양 조건이 분리 균주의 ECPs의 효소 활성 및 용혈능에 미치는 영향은 온도 10, 20, 30℃, pH 5, 7, 9, 염분 농도 0, 2, 6%, 2,2‘-dipyridyl 농 도 0, 100, 200 μM과 FeCl30, 10, 200 mM의 조건 에서 배양 후 활성을 조사하였다. 준비된 ECPs 를 0.1 M PBS (pH 7.2)로 희석하여 total protein 농도를 20 ㎍/㎖로 조정한 후 API zym kit (Bio- Merieux, Spain)를 이용하여 상법에 따라 분석하 였다.
Caseinase activity는 Austin et al.(1998)에 따라 azocasein(Sigma Co. USA)을 기질로 하여 측정 하였으며, gelatinase activity는 Zhang and Austin(2000)에 따라 caseinase activity 법을 변형 하여 측정하였으며, phospholipase와 lipase activi- ty는 Liu et al.(1996)에 따라 1%(w/v) agarose에 egg yolk 또는 1%(v/v) Tween 80을 첨가하여 clear zone을 측정하였다. 적혈구 용혈능은 Austin et al.(1998)에 따라 넙치 적혈구와 양적혈 구에 대하여 최대 희석 배수를 용혈소가로 평가 하였다.
Siderophore
활성ECPs 중 siderophore는 Schwyn and Nei- lands(1987)의 CAS liquid 법에 따라 Strepto-
myces pilosus의 siderophore인 desferal(Deferox-
amine mesylate salt, Sigma Co. Ltd.)을 이용하여 반응을 검정하였다. 10 mM HDTMA(hexade- cyltrimethyl ammonium)가 첨가된 iron(Ⅲ) solu- tion 1.5 ㎖(FeCl3∙6H2O 1 mM, HCl 10 mM)에 2 mM CAS 용액과 piperazine을 첨가하여 CAS as- say solution을 제작하여 ECPs, desferal 용액과 반응시켰다. TSB를 blank로 사용하여 630 nm에 서 흡광도를 측정하였으며, 결과는 siderophore units로 나타내었다.〔(Ar-As)/Ar〕× 100= % siderophore units Ar= reference abs 630; As= sample abs 630
Cytotoxicity
분리된 균주에서 생성된 ECPs의 세포 독성은 어류 유래 주화세포인 EPC(Epithelioma papillo-
sum cyprini)
를 이용하여 측정하였다.10% fetal bovine serum(FBS, GibcoBRL)과 1%
antibiotic-antimycotic agent(GibcoBRL)가 첨가된 Eagle's minimum essential medium(EMEM, Sig- ma)에 EPC가 1×106 cells/㎖의 농도가 되도록 조정하여 24-well plate(Corning, Inc.)에 200 ㎕씩 접종한 다음 20℃에서 24시간 단층 배양하였다.
배양 후 ECPs를 10% FBS 첨가 EMEM으로 단 백질 농도가 200 ㎍/㎖가 되도록 제작하여 각 well에 200 ㎕ 씩 주입한 다음 20℃에서 24시간 배양하였다. 배양된 plate는 Ho and Chou(2001) 의 방법에 따라 cell line을 2% formalin-PBS로 10분간 고정한 후 0.4% crystal violet-PBS로 10 분간 염색하였다. Well은 PBS로 세척 후 50%
ethanol(Merck)과 1% sodium-dodecyl sulfate-PBS 에 현탁하여 ELISA reader(630 nm)로 흡광도 값 을 측정하였으며, 대조구의 흡광도 값에 대한 ECPs 접종 well과 흡광도 값의 차이로 cytotoxi- city를 계산하였다.
혈청의 살균 작용에 대한 생존율
혈청의 살균 작용에 대한 P. damselae의 배양 조건별 생존율을 조사하였다. 즉, 각 조건에서 48시간 배양 된 균체는 4℃, 8,000rpm에서 10분 간 원심 분리한 다음 세척하여 멸균식염수로 106 cfu/㎖로 준비하였다. 준비된 균체는 넙치 혈 청을 동량 섞은 후 25℃ 진탕 배양기에서 반응 시키면서 0, 1, 3, 6 및 12 시간 경과할 때마다 단 계 희석하여 Miles and Misra(1938)에 따라 생균 수를 계수하였다.
통계 처리
결과의 통계 처리는 SPSS (SPSS Inc., 1997) program으로 ANOVA-test를 실시하여 Duncan′s multiple range test (Duncan, 1955)로 평균간의 유 의성을 검정하였다.
결 과
배양 조건에 따른 크기와 형태 변화
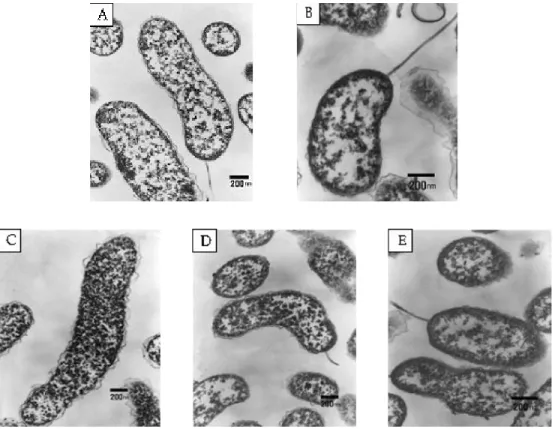
DIP 100 μM 첨가구와 NaCl 4% 첨가구의 균체 내부에는 대조구에 비하여 많은 과립이 관찰되 었으나, FeCl3100 mM 첨가구에서 배양한 균체 의 내부에서는 과립이 거의 관찰되지 않았으며, 대조구에 비하여 세포벽이 두꺼워졌다.
배양 조건이 균체의 크기에 미치는 영향은 배 양 조건별로 배양된 균체 30개의 크기를 전자현 미경으로 측정하였다. 균체의 크기는 대조구에 서 1.41±0.161×0.38±0.020 ㎛, FeCl3100 mM 첨가구에서 0.885±0.0.40×0.39±0.060 ㎛으로 장축의 길이가 대조구에 비하여 유의적으로 짧 게 나타났다 (Table 1) (p<0.05). DIP 100 μM 첨가 구에서는 1.56±0.505×0.34±0.040 ㎛, pH 9 시 험구에서는 1.59±0.205×0.39±0.016 ㎛로 대 조구와 유의적인 차이를 나타내지 않았다. 그러 나 배지 중의 NaCl 4% 시험구에서는 2.44±
0.382×0.56±0.066 ㎛로 장축의 길이가 대조구 에 비하여 유의적으로 길어졌다 (p<0.05).
균성장 및 Extracellular products (ECPs) 생성능 배양 조건이 P. damselae의 성장 및 ECPs 생 성에 미치는 영향을 조사한 결과 Fig. 2와 같이
나타났으며, Fig. 2의 결과를 이용하여 P. damselae 가 흡광도 600 nm에서 0.1 성장할 때 분비되는 ECPs의 total protein 농도를 계산하여 비교하였다.
Table 1. Size changes of Photobacterium damselae subsp. damselae under different incubation conditions. Variations of
P. damselae subsp. damselae on different conditions in sizeIncubation condition
Size of bacteria
Major axis(㎛) Minor axis(㎛)
Control 1.41 ±0.161
a0.38 ±0.020
pH 9 1.59±0.205
a0.39±0.016
NaCl 4% 2.44±0.382
b0.42±0.066
DIP 100μ M 1.56±0.505
a0.34±0.040
FeCl
3100mM 0.885±0.040
b0.39±0.060
Alphabetic superscripts indicate statistic significance among treatments(P<0.05).
Fig. 1. Transmission electron micrographs of Photobacterium damselae subsp. damselae cultured on different conditions.
A, Control; B, pH 9; C, NaCl 4%; D, Iron-limited(2,2 ‘-dipyridyl 100 μ M); E, Iron-added (FeCl
3100 mM) at 24hrs .
배양 온도가 P. damselae의 성장 및 ECPs 생 성에 미치는 영향을 조사한 결과 성장은 10~30
℃까지 가능하였으나, 10℃에서는 흡광도 600 nm 에서 0.16±0.01로 매우 낮게 나타났으며, 15~30
℃까지는 비슷한 흡광도 값을 나타내었다. ECPs 의 total protein 농도는 모든 조건에서 비슷하게 나타났으나, 10℃에서는 세균의 흡광도 값 0.16 에 ECPs의 protein 농도가 3.02로 세균이 흡광 도 값으로 0.1 성장할 때 ECPs의 protein은 1.89 g/㎗ 이었으나, 30℃에서는 세균의 흡광도 값 0.43에 ECPs의 proteins 농도가 3.10 g/㎗로 세균 이 흡광도 값으로 0.1 성장할 때 ECPs의 protein 은 0.76 g/㎗가 분비되어 배양 온도가 낮을수록 분비되는 ECPs protein 농도는 높게 나타났다 (Fig. 2A).
pH가 P. damselae의 성장 및 ECPs 생성에 미
치는 영향을 조사한 결과 pH 5~9까지는 0.331~0.397로 성장이 비슷하게 나타났으나, pH 3과 11에서는 거의 성장이 나타나지 않았다.
ECPs의 total protein 농도는 pH 9에서 가장 높게 나타났으나, 세균이 흡광도 값으로 0.1 성장할 때 ECPs의 protein은 pH 11에서 10 g/㎗ 로 가장 높게 나타났으며, pH 7에서 0.76 g/㎗로 가장 낮 게 나타났다 (Fig. 2B).
NaCl 농도가 P. damselae의 성장 및 ECPs 생 성에 미치는 영향을 조사한 결과 0~6%까지는 성장이 가능하였으나, 8% 이상에서는 성장을 하지 않았다. ECPs의 total protein 농도는 염분농 도 2%에서 가장 높게 나타났으나, 세균이 흡광 도 값으로 0.1 성장할 때 ECPs의 protein은 염분 농도 10%에서 13 g/㎗ 로 가장 높게 나타났으 며, 염분농도 2%에서 0.76 g/㎗로 가장 낮게 나
Fig. 2. Effects of culture conditions(A, temperature; B, pH; C, NaCl; D, 2,2'-dipyridy; E, FeCl
3)on the growth and ECPs pro-
duction of Photobacterium damselae subsp. damselae in TSB. Total protein concentrations of ECPs and cell density of bac-
teria at 24 hrs after incubation.
타났다 (Fig. 2C).
Iron-limited 조건이 P. damselae의 성장 및 ECPs 생성에 미치는 영향을 조사한 결과 iron- limited인 DIP 100 μM에서부터 성장 억제가 나타 났으며, 300~400 μM에서는 성장이 거의 나타나 지 않았다. ECPs의 total protein 농도는 DIP 0 μM
에서 가장 높게 나타났으나, 세균이 흡광도 값으 로 0.1 성장할 때 ECPs의 protein은 DIP 400 μM 에서 11 g/㎗ 로 가장 높게 나타났으며, DIP 0 μM 에서 0.76 g/㎗로 가장 낮게 나타났다 (Fig. 2D).
FeCl3 첨가 조건이 P. damselae의 성장 및 ECPs 생성에 미치는 영향을 조사한 결과 FeCl3
Table 2. Enzymatic and hemolytic activities of ECPs of Photobacterium damselae subsp. damselae grown under different
culture conditions
Culture Temp. pH NaCl (%) 2,2、 -dipyridyl FeCl
3condition (℃) (μ M) (mM)
Enzyme 10 20 30 5 7 9 0 2 6 0 100 200 0 10 200
Negative control - - - - - - - - - - - - - - -
Alkaline phosphatase + + + + + + + + + + + + ++ ++ ++
Esterase(C 4) + + + + + + + + + + + + + + +
Esterase Lipase(C 8) + + + + + + + + + + + + + + +
Lipase(C 14) - - - - - - - - - - - - - - -
Leucine arylamidase + + + + + + + + + + + + + + +
Valine arylamidase - - - + + - + + - + + + + + +
Cystine arylamidase - - - - - - - - - - - - - - -
Trypsin - + - + + - + + - + + + + + +
α -chymotrypsin - + - + + - + + - + + + + + +
Acid phosphatase - + + + + + + + - + + + + + +
Naphthol-AS-BI
-phosphohydrolase + + + + + + + + + + + + + + +
β -glucosidase - - - - - - - - - - - - - - -
N-acetyl-β-glucosaminidase + + + + + + + + - + + + + + +
α -mannosidase - - - - - - - - - - - - - - -
α -fucosidase - - - - - - - - - - - - - - -
Gelatinase + + + + + + + + + + + + + + +
Caseinase + ++ + ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++
Phospholipase +++ ++ ++ +++ ++ ++ ++ ++ ++ ++ +++ +++ ++ ++ +
Lipase + + + + + + + + + + + + + + +
Hemolytic titeration
FRBCs
1)16 32 32 16 32 32 32 32 16 32 64 256 32 16 4
SRBCs
2)8 16 16 8 16 16 16 16 8 16 32 128 16 16 4
* White area : -, negative; +, positive
Gray area : -, no zone of clearing; +, diameter of clearing or opalescence zone below 3mm; ++, diameter of clearing or opalescence zone between 3.5~6mm; +++, diameter of clearing or opalescence zone over 6.5 mm.
1)
FRBCs : Flounder red blood cells,
2)SRBCs: Sheep red blood cells.
의 농도가 높을수록 성장량이 높게 나타났으나, ECPs의 protein 농도는 낮게 나타났다. 세균이 흡광도 값으로 0.1 성장할 때 ECPs의 protein은 FeCl3 0 mM에서 0.76 g/㎗ 로 가장 높게 나타났 으나, 200 mM에서 0.21 g/㎗로 가장 낮게 나타 났다(Fig. 2E).
ECPs의 효소 활성 및 용혈능
배양 조건이 P. damselae의 ECPs의 효소 활성 및 용혈능에 미치는 영향을 조사한 결과는 Table 2와 같다. 배양 온도에 따른 ECPs의 효소 활성은 trypsin, α-chymotrypsin 및 acid phos- phatase에서 차이가 관찰되었다. 용혈능 조사 결 과, 넙치 적혈구에 대한 ECPs의 용혈소가는 10
℃ 배양 조건에서 16이었으며 20℃ 이상일 때 32로 나타났다. 배양 배지의 pH에 따른 ECPs의 효소 활성은 valine arylamidase에서 차이가 관찰 되었다. 용혈능 조사 결과, 넙치 적혈구에 대한 ECPs의 용혈능은 pH 5일 때 16, pH 7과 9에서 는 32이었으며, 양의 적혈구에 대해서는 각각 8 과 16으로 나타났다. 배양의 염분 농도에 따른 ECPs의 효소 활성은 acid phosphatase와 valine arylamidase에서 차이가 관찰되었다. 용혈능 조 사 결과, 넙치 적혈구에 대한 용혈능이 양 적혈
구에 비하여 높게 나타났으며, 염분 농도 6%에 서는 0%와 2% 첨가구에 비하여 낮은 활성을 나타내었다. Iron-chelator 첨가에 따른 ECPs의 효소 활성은 DIP 농도가 높을수록 phospholi- pase 활성이 높았으며 용혈능도 높게 나타나, 200 μM 첨가구에서의 용혈소가가 넙치 적혈구 와 양 적혈구에 대하여 각각 256과 128로 나타 났다. Iron-added에서 ECPs의 효소 활성은 FeCl3 첨가에 따른 차이를 보이지 않았다. 용혈능을 조사한 결과, 넙치 적혈구에 대한 용혈능이 양 적혈구에 비하여 높게 나타났으며, FeCl3의 첨가 농도가 증가할수록 용혈능은 낮게 나타났다.
Siderophore
활성배양 조건에 따른 ECPs 중의 siderophore의 활성을 측정한 결과는 Fig. 3과 같다. 모든 조건 에서 시간이 경과함에 따라 siderophore 생성량 이 증가하였으며, 배양 48시간 째 iron-limited 조건인 DIP 100 μM 첨가구에서 배양하여 분리 한 ECPs의 siderophore 활성은 98.7±5.7%
siderophore units로 대조구의 70± 6.8%
siderophore units보다 유의적으로 높은 값을 나 타내었으며(p<0.05), iron-added 조건인 FeCl3100 mM 첨가구에서 배양된 균의 ECPs에서는 45.8
Fig. 3. Effects of different incubation conditions on the siderophore production of Photobacterium damselae subsp. damselae. The activity of siderophore was deter- mined by CAS liquid assay. Alphabetic superscripts indi- cate statistic significance among treatments(P<0.05).
Fig. 4. Effects of the incubation conditions of Photobacteri-
um damselae subsp. damselae varying with temperature,pH, NaCl, 2,2'-dipyridyl, and FeCl
3concentrations on the
cytotoxicity in EPC. Alphabetic superscripts indicate statis-
tic significance among treatments(P<0.05).
±5.4units로 대조구에 비하여 유의적으로 낮은 활성을 나타내었다.
Cytotoxicity
조건별로 배양된 세균의 ECPs가 어류 주화세 포인 EPC에 미치는 영향을 조사한 결과는 Fig. 4 와 같다. 온도, 염분농도와 pH에 의해서는 대조 구와 유의적 차이가 없이(p>0.05) 각각 37.5±
4.5~45.5±6.5%, 48.9±3.6~54.3±7.4%와 40.5
±5.9~45.5±7.4%로 나타났다. DIP 첨가구에서 는 DIP 농도가 증가할수록 EPC에 대한 cytotox- icity가 높아져 DIP 200 μM이상에서는 대조구에 비하여 유의적으로 높아(p<0.05), 100 μM에서는 51.8±4.9%, 400 μM에서는 78.9±10.5%로 나타
났다. FeCl3 첨가 배지에서는 FeCl3 농도가 높을 수록 cytotoxicity가 낮게 나타나 100 mM이상의 구에서는 대조구에 비하여 유의적으로 낮아 (p<0.05), 10 mM에서는 39.6±6.8%, 200 mM에 서는 15.5±5.5%로 나타났다.
혈청의 살균 작용에 대한 생존율
배양 조건이 넙치 혈청의 살균 작용에 대한 P.
damselae
의 생존능에 미치는 영향을 조사한 결과는 Fig. 5와 같다.
배양 배지의 pH를 달리하여 배양한 결과 넙 치 혈청에 대하여 pH5와 pH9에서 배양한 균체 가 pH7에서 배양한 균체에 비하여 다소 생존율 이 높게 나타났으나(Fig. 5A), 염분 농도에 따른
Fig. 5. Survival rates of Photobacterium damselae subsp. damselae(10
6cfu/㎖) incubated on different conditions in the serum of olive flounder. P. damselae was grown in the medium supplemented with varying concentrations of NaCl(%), (A), 2,2'- dipyridyl(μ M), (B), FeCl
3(mM), (C) and pH (D).
(A)
(C) (D)
(B)
차이는 보이지 않았다. 배지에 iron-chelator인 DIP를 첨가하여 배양된 P. damselae를 혈청과 반응시켰을 때 대조구에 비하여 생존율이 매우 높게 나타났으나 (Fig. 5B), 첨가된 DIP의 농도 에 따른 차이는 보이지 않았다(Fig. 5C). 배지에 iron-added 조건으로 FeCl3를 첨가하여 배양한 P.
damselae를 혈청과 반응 시 대조구에 비하여 생
존율이 낮았으며(Fig. 5D), 첨가된 농도가 높을 수록 생존율이 낮게 나타났다.고 찰
P. damselae의 병원성 기작에 관한 연구로는
cytolytic toxin과 haemolytic toxin의 분비와 활성 (Kreger 1984; Kreger et al., 1987; Fouz et al., 1993) 및 damselysin이라는 phospholipase 효소 의 존재 (Kreger et al., 1987)와 이 효소에 의한 조혈 조직 파괴와 면역 반응 저하(Fouz et al., 1995) 등이 알려져 있다. 그러나 이 병원체는 감 염 대상이 되는 숙주의 범위가 넓어 어종에 따 른 병원성의 발현 기작 및 정도가 다를 것으로 예상되어, 본 연구에서는 배양 조건에 따른 실 험균의 형태 및 크기의 변화 관찰, 성장 시 분비 되는 ECPs의 protein 농도, 효소활성, 용혈능, siderophore 활성 및 cytotoxin 활성과 넙치의 살 균작용에 대한 균체의 생존능 변화에 대하여 조 사하였다.배양 조건에 따른 균체 형태를 관찰한 결과, DIP 첨가구에서는 대조구에 비하여 세포 내 많 은 과립이 관찰되었으며, FeCl3 첨가구에서는 대 조구에 비하여 세포벽이 두꺼워졌다. Brown and Williams (1985)는 모든 세균이 환경의 변화에 대한 적응 방법으로 세포 표면의 구성 요소를 변화시키며, 특히 iron과 같은 metal ion은 poly- saccharide 합성 시 cofactor로서 작용하여 그 영 향을 더욱 크게 한다고 하였다. Jacques et
al.(1994)은 Pasteurella multocida를 iron-restric-
tion과 iron-sufficient 조건에서 배양한 결과, DIP 의 농도가 증가할수록 capsule의 두께가 얇아졌으며, FeCl3의 농도가 증가할수록 두터워져 본 연구와 같은 결과를 나타내었다. 또한 Abeck et
al.(1990)은 Haemopbilus ducreyi를 iron-limited
조건에서 배양 시 periplasm에 큰 space가 형성 된다고 하였으며, Ferrala et al.(1986)은 Azotobac-ter abroococcum을 iron starvation 조건에서 배양
하면 diffuse와 extensive가 많아 환경에서 철 흡 수 기능이 증가된다고 하였다.배양 조건에 따른 성장 및 ECPs의 protein 농 도 조사 결과, 성장 속도와 분비되는 ECPs의 protein 농도는 일치하지 않았으며, 균체가 흡광 도 (600 nm) 값으로 0.1 성장할 때 온도 조건구 에서는 30℃, pH 조건구에서는 pH 9, NaCl 첨가 구에서는 NaCl 10%, DIP 첨가구에서는 DIP 400 μM에서 ECPs의 protein 농도가 가장 높게 나타 났으며, FeCl3 첨가구는 모두 대조구에 비하여 낮게 나타났다.
ECPs 중의 효소활성 및 용혈능 조사 결과, 다 른 배양 조건에 비하여 배양 배지 중의 철의 농 도에 영향을 많이 받는 것으로 조사되었으며, siderophore, cytotoxin 활성 및 넙치 혈청의 살균 작용에 대한 생존율에서도 유사한 결과를 나타 내었다. 즉, iron-limited 조건에서 phospholipase 활성과 용혈능이 다른 배양 조건에 비하여 높아 졌으며, iron-added 조건에서는 낮게 나타났다.
Kreger(1984)는 V. damsela 19 균주에 대하여 용 혈능과 병원성과의 관계를 조사한 결과 병원성 이 높은 균주에서 hemolytic units가 높게 나타났 으나, 본 연구에서는 iron-limited 조건에서 배양 시 용혈능이 높아져 iron-limited 환경은 병원성 을 높일 수 있을 것으로 생각된다. 또한, Serratia
marcescens에 대하여 온도를 달리하여 배양 시
30℃의 용혈능이 37℃보다 10배 높게 나타났으 며(Poole and Braun, 1988), exocellular nuclease 활 성은 저하되었으나(Jepsen et al., 1987), 본 연구 에서는 온도 조건에 따라 용혈능에 큰 차이를 보이지 않아 온도의 영향은 균주의 종류에 따라 차이가 있는 것으로 생각된다.병원성 세균이 숙주의 철을 획득하는 능력은
감염 시 필수적인 것으로 (Weinberg, 1984), Litwin and Calderwood(1993)는 세균이 숙주에서 free iron을 취하거나 다양한 mechanism을 통하 여 숙주의 철결합물을 바로 이용한다고 하였다.
또한, Ho et al.(2004)은 세균이 숙주의 철을 이용 시 siderophore나 hemolysin을 분비하여 heme 복합체에서 철을 유리시켜 이용하거나, siderophore 없이 transferrin, hemoglobin 및 heme-albumin과 같은 숙주 철 결합물을 곧바로 이용하는 등 다 양한 경로가 있다고 보고하였다. 세균은 철을 이용하기 위하여 환경변화에 따라 siderophore 활성에 변화를 나타내는데(Brown and Willams, 1985), 본 연구에서도 iron-limited 조건에서 siderophore 활성이 높아지고 넙치 혈청의 살균 작용에 대한 생존률도 높게 나타났다. Fouz et
al.(1994)은 혈청의 살균 작용에 대한 P. damse- lae의 생존능 검토 결과, iron 이용능이 높을수록
병원성이 높아진다고 보고하였으며, 본 연구에 서도 iron 이용능이 높은 iron-limited 조건 (Fer- rala et al., 1986)에서 siderophore 활성이 높아져 iron에 대한 이용능이 증가한 것으로 추정된다.또한 P. damselae는 in vivo와 in vitro에서 높은 용혈능을 나타내며(Kreger et al., 1987; Fouz et al., 1992; 1993), 용혈 후 유리되는 iron을 생존과 증 식에 이용하여 숙주에 병원성을 나타내므로 (Fouz et al., 1994), 본 연구에서 iron-limited 조건 에서 배양한 균체의 높은 용혈능과 siderophore 및 cytotoxin 활성이 넙치 혈청에 대한 생존률을 높인 것으로 추정된다.
세균의 iron-limited 배양 조건이 cytotoxcicity 에 미치는 영향에 대하여 일부 세균에서 보고되 었다. Ho et al.(2004)은 Salmonella choleraesuis SC-5를 iron-limited 조건에서 배양 시 cytotoxin 생성량이 증가한다고 하였으나, Wong and Lee(1995)는 V. parahaemolyticus에서는 strain에 따라 다르다고 보고하였다.
배양 조건이 넙치 혈청 살균능에 대한 P.
damselae의 생존능에 미치는 영향을 조사한 결
과, iron-limited 조건에서 높아졌다. Ho etal.(2004)은 Salmonella choleraesuis SC-5를 iron-
limited 조건에서 배양 시 cytotoxin 생성량이 증 가되고, iron-rich에서 배양한 균에 비하여 mouse serum에 대한 생존율이 높아 철 결핍 조건에서 병원성이 높게 나타나 본 연구와 같은 결과를 나타내었다.Iron-limited 배양 조건은 어체 내와 유사한 환 경으로 배양균의 성장을 느리게 하지만(Fernan- des-Beros et al., 1989; Chart and Rowe, 1993), 병 원체에 의해서 생성되는 다양한 병원성 factor는 증가시키는 것으로 판단된다. 본 연구에서도 iron-limited 조건에서 배양 시 효소 활성, 용혈 능, siderophore 분비 및 cytotoxin 생성이 증가되 어, 넙치 혈청의 살균 작용에 대한 생존능이 높 아지므로, 어체에 감염 시 어체 내에서도 유사 한 현상이 나타나 넙치에 높은 병원성을 나타내 는 것으로 생각된다.
따라서 백신의 효과는 숙주의 면역계가 감염 원의 특이 항원을 인식하는 정도에 따라 좌우되 므로(Colquhoun et al., 2002) 효과적인 백신의 개 발을 위해서는 숙주에 감염 시 발현되는 특이적 인 상태의 균주를 이용하는 것이 필요하다고 사 료되어 숙주 내와 유사한 iron-limited 조건에서 배양된 P. damselae의 면역반응에 대한 검토가 필요할 것으로 사료된다.
요 약
배양 온도, 배양 배지 중의 pH, NaCl 농도, iron 농도와 같은 배양 조건이 P. damselae의 미 생물학적 성상 및 병원성에 미치는 영향을 조사 하였다. P. damselae는 배양 온도 15~30℃에서 배지의 pH 5~9, NaCl 농도 0~6%와 FeCl3 첨가 시 성장이 촉진되었지만, iron-chelator 첨가 시 성장이 억제되었다. Iron-limited 조건에서 배양 시 ECPs protein 농도, phospholipase, 용혈능, siderophore 및 cytotoxicity 활성 이 높게 나타났 고, 넙치 혈청의 살균작용 대한 생존율도 증가 되었으나, iron-added 조건에서는 감소되어, 배양
조건 중 iron 농도에 영향을 가장 많이 받는 것 으로 나타났다. 따라서, 본 연구에서 적용한 iron-limited 조건은 철농도가 낮은 숙주 내와 유 사한 환경이므로 iron-limited에서 P. damselae 배 양 시 나타나는 병원성의 증가는 어체에 감염 시 나타나는 병원성 발현 기전과 유사할 것으로 사료된다.
감사의 글
본 연구는 국립수산과학원 (양식생물 백신 연구, RP-2009-AQ-044)의 지원에 의해 운영되 었습니다.
참 고 문 헌
Abeck, D., Johnson, A.P., Korting, H.C., Stolz, W., Struck-Suer, E. and Braun-Falco, O.: Effect of iron limitation on protein composition and ultrastructure of Haemophilus ducreyi.
FEMS Microbiol. Lett., 68:41-44, 1990.
Austin, B., Austin, D.A., Dalsgaar, I., Gudmundsd бttir, B.K., Thornton, S.H., Larsen, J.L., O'Hici, B. and Powell, R.: Characterization of atypical Aeromonas salmonicida by dif- ferent methods. Syst. Appl. Microbiol., 21:
50-64, 1998.
Brown, M.R., Anwar, H. and Costerton, J.W.: Sur- face antigen in vivo; a mirror for vaccine de- velopment. Can. J. Microbiol., 39:491-498, 1988.
Brown, M.R. and Williams, P.: The influence of en- vironment on envelope properties affecting survival of bacteria in infections. Ann. Rev.
Microbiol., 39:527-556, 1985.
Chart, H. and Rowe, B.: Iron restriction and growth of Salmonella enteritidis. Epidemiol. Infect.
110:41-47, 1993.
Colquhoun, D.J., Alvheim, K., Dommarsnes, K.,
Syvertsen, C. and S�rum, H.: Relevance of incubation temperature for Vibrio salmonici-
da vaccine production. J. Appl. Microbiol.,
92:1087-1096, 2002.Davies, R.L., Gibbs, H.A., McCluskey, J., Coote, J.G., Freer, J.H. and Parton, R.: Develop- ment of an intraperitoneal implant chamber for the study of in vivo-grown Pasteurella
haemolytica in cattle. Microbial Pathogene-
sis, 16:423-433, 1994.Duncan, D.B.: Multiple-range and multiple F tests.
Biometrics, 11:1-42, 1955.
Fernandez-Beros, M.E., Gonzalez, C., McIntosh, M.A. and Cabello, F.C.: Immune response to the iron-deprivation-induced protein of Sal-
monella typhi in thyphoid fever. Infect. Im-
mun., 57:1271-1275, 1989.Ferrala, N.F., Champlin, A.K. and Fekete, F.A.:
Morphological differences in the capsular polysaccharide of nitrogen-fixing Azotobac-
ter chroococcum B-8 as a function of iron
and molybdenum. FEMS Microbiol., 33:137-142, 1986.Fouz, B., Toranzo, A.E., Biosca, E.G., Mazoy, R.
and Amro, C.: Role of iron in the patho- genicity of Vibrio damsela for fish and mammals. FEMS Microbiol. Lett., 121:181- 188, 1994.
Fouz, B., Novoa, B., Troanzo, A.E. and Figueras, A.: Histopathological lesions caused by Vib-
rio damsela in cultured turbot, Scophthal- mus maximus(L.): inoculations with live
cells and extracellular products. J. Fish Dis., 18:357-364, 1995.Fouz, B., Barja, J.L., Rivas, C., Amaro, C. and Toranzo, A.E.: Toxicity of the extracellular products of Vibrio damsela isolates from dis- eased fish. Curr. Microbiol., 27:341-347, 1993.
Fouz, B., Larsen, J.L., Nielsen, B., Barja, J.L. and Toranzo, A.E.: Characterization of Vibrio
damsela strains isolated from turbot Scoph- thalmus maximus in Spain. Dis. Aquat. Org.,
12:155-166, 1992.Fujioka, R.S., Greco, S.B., Cates, M.B. and Schroeder, J.P.: Vibrio damsela from wounds in bottlenose dolphins Tursiops
truncatus. Dis. Aquat. Org., 4:1-8, 1988.
Garduno, R.A., Thornton, J.C. and Kay, W.W.: Fate of the fish pathogen Aeromonas salmonicida in the peritoneal cavity of rainbow trout.
Can. J. Microbiol., 39:1051-1058, 1993a.
Garduno, R.A., Thornton, J.C. and Kay, W.W.:
Aeromonas salmonicida grown in vivo. In-
fect. Immun., 61:3854-3862, 1993b.Ho, W.-L. and Chou, C.-C.: Effects of carbon and nitrogen sources, sodium chloride and cul- ture conditions on cytotoxin production by
Salmonella choleraesus. Inter. J. Food Mi-
crobiol., 67:81-88, 2001.Ho, W.-L., Yu, R.-C. and Chou, C.-C.: Effect of iron limitation on the growth and cytotoxin production of Salmonella choleraesuis SC- 5. Inter. J. Food Microbiol., 90:295-302, 2004.
Jacques, M., Belanger, M., Diarra, M.S., Dargis, M.
and Malouin, F.: Modulation of Pasteurella
multocida capsular polysaccharide during
growth under iron-restricted condition and invivo. Microbiol., 140:263-270, 1994.
Jepsen, P.K., Riise, E., Biedermann, K., Kristensen, P.C. and Emberg, C.: Two-level factrial screening for influence of temperature, pH, and aeration on production of Serratia
marcescens nuclease. Appl. Environ. Micro-
biol., 53:2593-2596, 1987.Kreger, A.S.: Cytolytic activity and virulence of
Vibrio damsela. Infect. Immun., 44:326-331,
1984.
Kreger, A.S., Bernheimer, A.W., Etkin, L.A. and Daniel, L.W.: Phospholipase D activity of
Vibrio damsela cytolysin and its interaction
with sheep erythrocytes. Infect. Immun., 55:3209-3212, 1987.Litwin, C.M. and Calderwood, S.B.: Role of iron in regulation of virulence genes. Clin. Microbi- ol. Rev., 6:137-159, 1993.
Liu, P.C., Lee, K.K. and Chen, S.N.: Pathogenicity of different isolates of Vibrio harveyi from diseased Kuruma prawns Penaeus
japonicus. Curr. Microbiol., 33:129-132,
1996.Love, M., Fisher, D.T., Horse, J.E., Farmer, J.J., Hickman, F.W. and Fanning, G.R.: Vibrio
damsela, as a marine bacterium, causes skin
ulcers on the damselfish Chromis punctipin-nis. Science, 214:1140-1141, 1981.
Miles, A.A. and Misra, S.S.: The estimate of the bactericidal power of the blood. J. Hygiene, 38:873-885, 1938.
Poole, K. and Braun, V.: Influence of growth tem- perature and lipopolysaccharide on hemolyt- ic activity of Serratia marcescens. J. Bacteri- ol., 170:5146-5152, 1988.
Sakata, T., Matsuura, M. and Shimokawa, Y.: Char- acteristics of Vibrio damsela isolated from diseased yellowtail(Seriola quinqueradiata).
Nippon Suisan Gakkashi, 55:135-141, 1989.
Schwyn, B. and Neilands, J.B.: Universal chemical assay for the detection an determination of siderophores. Anal. Biochem. 160:46-56, 1987.
SPSS Inc.: SPSS Base 7.5 for Window, SPSS Inc., 444N. Michigan Avenue, Chicago, IL, 60611, 1997.
Truper, H.G. and De'Clari, L.: Taxonomic note:
necessary correction of specific epithets
formed as substantives (nouns) 'in apposi- tion', Int. J. System. Bacteriol., 41:529-534, 1997.
Vera, P., Navas, J.L. and Fouz, B.: First isolation of
Vibrio damsela from seabream(Sparus aura- ta). Bull. Eur. Assoc. Fish Pathol., 11: 112-
113, 1991.Weinberg, E.D.: Iron withholding: a defense against infection and neoplasia. Physiol. Rev., 64:65-102, 1984.
Wong, H.C. and Lee, Y.S.: Effect of iron on the re- covery of viable bacteria from the cardiac blood in an experimental mouse model and
on the serum resistance and macrophage phagocytosis of Vibrio parahaemolyticus. J.
Diarrhoeal Dis. Rev., 13:219-223, 1995.
Zhang, X.-H. and Austin, B.: Pathogenicity of Vib-
rio harveyi to salmonids. J. Fish Dis., 23:93-
102, 2000.권문경, 박상언, 방종득, 박수일: 넙치, Par-
alichthys olivaceus에서 병원성 Photobac- terium damselae subsp. damselae의 분리.
한국어병학회지, 18:205-214, 2005.