한국어병학회지 제
22
권 제3
호(2009) J. Fish Pathol., 22(3) : 375 ~ 382 (2009)
375
�
Corresponding Author :
Various methods have been developed to deliver the target genes into cells. These methods include physical approaches such as direct DNA injection, encapsulation of DNA into liposomes, and gene gun technology (Cooper, 1996; Yang et al., 1996;
Prince, 1998) and gene delivery by retrovirus parti- cles having recombinant genomes. Among them, retroviral-mediated gene transfer system has some distinct advantages such as efficiently introducing genes into any dividing cell types, stable integration into chromosomal DNA, faithful transmission of encoded gene and simultaneous introducing gene into large numbers of cells (Morgan et al., 1993).
Thus it has been a powerful tool for gene therapy and identification of gene function in mammals
(Kolodka et al., 1993; Olsen et al., 1993).
Retroviruses having RNA genome replicate through an integrated DNA intermediate (Linial et al., 2005). The retroviral genome contains gag gene encoding group-specific antigen, pol gene encoding viral enzymes including reverse transcriptase, inte- grase and protease, env gene for viral envelope pro- teins and long terminal repeats (LTRs) at each end of genome. LTRs are essential for the initiation of viral DNA synthesis, integration of proviral DNA and the regulation of viral gene transcription.
Therefore, for integration of a target gene into chro- mosomal DNA of cells, the target gene flanked with LTRs was constructed and transfected into a stable cell line wherein pol gene was expressed. In this
Integration of a target gene into chromosomal genome of BF-2 cells using UV-inactivated snakehead retrovirus (SnRV)
Se Ryun Kwon
�, Toyohiko Nishizawa and Mamoru Yoshimizu Faculty of Fisheries Sciences, Hokkaido University, Hakodate, 041-8611, Japan
Integration and expression of a target gene into chromosomal genomes of host cell by retrovirus mediated gene transfer system usually require complicate and laborious procedures. In the present study, we investi- gate a simple method to integrate a target gene into genome of BF-2 cells using ultraviolet (UV)-inactivated snakehead retrovirus (SnRV), a fish retrovirus. First of all, an optimization of transfection condition was determined with BF-2 cells using Lipofectamine 2000 and Transome. Using 0.5 ㎕ Lipofectamine 2000 resulted in 33.8, 40.6 and 40.2% of transfection efficacy with high survival rate (minimum 80%) in 0.5, 1 and 2 ㎍ DNA, respectively, and those of Transome were all less than 5%. It was confirmed that UV-treat- ment for 5 min was enough to inactivate infectivity of SnRV. Next, a cassette composed of GFP (green flu- orescent protein) gene flanked by LTR (long terminal repeats) sequences derived from SnRV was con- structed and transfected into BF-2 cells followed by treatment with UV-inactivated SnRV for optimization of integration and expression of the cassette gene. As the results, the fluorescence was expressed in BF-2 cells treated with UV-inactivated SnRV 3 and 5 times, while there was no expression in BF-2 cells with once and non treatment. Accordingly, it was confirmed that GFP gene was integrated into chromosomal genome of BF-2 cells with UV-inactivated SnRV.
Key words: Integration, Transfection, Snakehead retrovirus, SnRV, Ultraviolet-inactivation
�
Corresponding Author : Se Ryun Kwon, Tel : +81-138-40-8810 Fax : +81-138-40-8810, E-mail : [email protected]
19...
376
procedure, multiple plasmids for expression of the target gene and pol gene or single plasmid harbor- ing multiple expression cassettes of those genes have been used. However, using the multiple plas- mids system may result in low transfection efficien- cy, while using the single plasmid system may lead to suppression of gene expression due to promoter interference (Hu et al., 2009).
In the present study, we attempt a simple method to integrate a target gene into host cell genome using an integrase of retrovirus which was inacti- vated by ultraviolet (UV) irradiation. As a model in the present study, snakehead retrovirus (SnRV) and BF-2 cell line were adopted. SnRV, a fish retro- virus, was persistently infected in SSN-1 cell line derived from the Southeast Asian striped snakehead fish (Ophicephalus striatus), and complete genome of SnRV was sequenced (Frerichs et al., 1991;
Hartet et al., 1996). BF-2 cells derived from bluegill fry (Lepomis machrochirus) are susceptible to SnRV (Frerichs et al., 1991). Here, green fluores- cent protein (GFP) gene with LTR sequences origi- nated in proviral genome of SnRV was tried to trans- fect to BF-2 cells for integration of the target gene into host cell genome by UV-inactivated SnRV.
Materials and Methods
Cells and Virus
BF-2 and SSN-1 cells were maintained using minimum essential medium (MEM) and Leibovitz (L-15) medium, respectively, and which were sup- plemented with 10% (v/v) fetal bovine serum (FBS), 100 IU of penicillin G and 100 ㎍/㎖ of streptomycin.
SnRV was obtained from culture fluid of SSN-1 cells spontaneously producing SnRV after centrifu- gation (3,000 x g, 10 min) to remove cell debris.
Infectivity titer of SnRV was determined using 96- well microplates seeded with BF-2 cells, and cyto- pathic effect (CPE) was evaluated to determine 50% of tissue culture infectious dose (TCID
50).
Transfection of BF-2 cell line
The pCMS - EGFP with 5.5 kb in size (Clontech, Palo Alto, CA) containing green fluorescent protein (GFP) gene under the CMV promoter was used.
Large amount of plasmid was prepared from Eschrichia coli DH5αby using the QIAGEN plas- mid mini kit (Qiagen, Maryland, USA) according to the manufacture’ s instruction. The obtained plas- mid was adjusted to 0.5-1 ㎎ DNA/㎖ by absorbance at 260 nm.
BF-2 cells were seeded at a density of 5×10
5cell/well in 24-well plates 1 day before transfection.
Monolayers with 80% confluence were transfected with different concentration of pCMS-EGFP using Lipofectamine 2000 (Invitrogene, Carlsbad, CA) or Transome (Wako, Osaka, Japan) according to the manufacturer’s instructions. Both transfection reagents, Transome and Lipofectamine 2000, were based on liposome origin. After 3 days of transfec- tion, green fluorescence was observed under an Olympus inverted microscope fitted with a FITC filter set. Transfection efficiency was calculated by percentage of cell numbers expressing GFP fluores- cence to total cell numbers in each well. And cell survival rate was calculated by percentage of cell numbers remaining in the well to number of initial cells plated. All procedures were conducted tripli- cates.
Inactivation of SnRV by ultraviolet irradiation SnRV was irradiated with 250 ㎼/ cm
2of UV lamp for 1, 3, 5, 10 and 20 min to break SnRV genome, and inactivation of the UV-treated SnRV was con- firmed by titration with BF-2 cells.
Se Ryun Kwon, Toyohiko Nishizawa and Mamoru Yoshimizu
377 Genome integration using non-replicable SnRV
Construction of LTR-GFP-LTR cassette for inte- gration
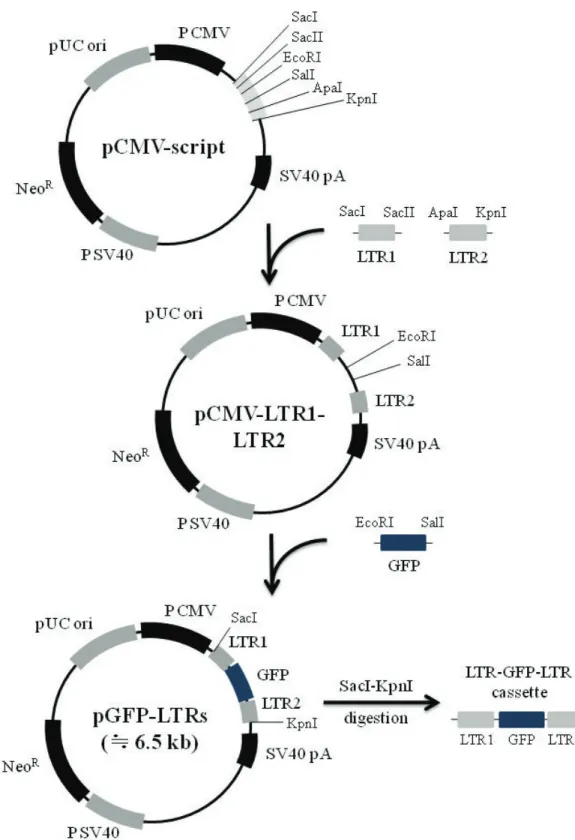
LTR1 and LTR2 genes of SnRV provirus (Hart et al., 1996) were amplified using following PCR primer sets the primer set for LTR 1 gene was LTR1 SacI (5′ -ccg agc tcc ttg tga cac cac tat act tc-3′ ) and LTR1 SacII (5′ -aac cgc ggt cac cca agt aga acg ctg ag-3′ ) including each restriction enzyme site (under bar), and the primer set for LTR2 gene was LTR2 ApaⅠ (5′ -aag ggc ccc tga aaa tgt gtc att tct gg-3′ ) and LTR2 KpnI (5′ -gag gta ccc tgt aaa cgg tga aag tac g-3′ ) including each restriction enzyme site (under bar). After digested with each restriction enzyme, the LTR genes were ligated into the multi- ple cloning site (MCS) of pCMV-script vector (Stratagene, La Jolla, CA) to transform E. coli DH5a (pCMV-LTR1-LTR2). Next, GFP gene was amplified from pCMS-EGFP using PCR primers, EGFP-F (5′ -ccg aat tcg ata tcg cag cac cat ggc ctg aaa taa cc-3 ′ ) containing EcoRI-site (under bar) and EGFP-R (5′ -gtg tcg act cga gct gcc cca gct ggt tct ttc c-3′) containing SalI-site (under bar). After digestion with EcoRI and SalI, the GFP gene was inserted into the corresponding MCS, which located between LTR1 and LTR2 genes of pCMV-LTR1- LTR2 (pGFP-LTRs, Fig. 1). The GFP gene flanked by LTRs was cut out from pGFP-LTRs by digestion with SacI and KpnI, and gel purified by GeneClean kit (MP Biomedicals, Solon, Ohio) according man- ufacture’ s instruction. This gene cassette was sub- jected to the following integration experiments.
Integration of LTR-GFP-LTR cassette into BF-2 cell genome
The purified gene was transfected into BF-2 cells seeded in four wells using 0.5 ㎕ Lipofectamine 2000 under the condition described above. After incubation at 25 � C for 16 h, the transfected cells were washed out with MEM 3 times, and inoculated
with 100 ㎕ of the UV-inactivated SnRV. The inocu- lated cells were incubated at 25� C for 10 days. Dur- ing the incubation, 1, 3 or 5 times of inoculation with UV-inactivated SnRV were conducted for the integration induction in each well at 3 days intervals.
The cells expressing fluorescence by GFP in each well were counted at 10 days post transfection.
Results and Discussion
Optimization of transfection condition
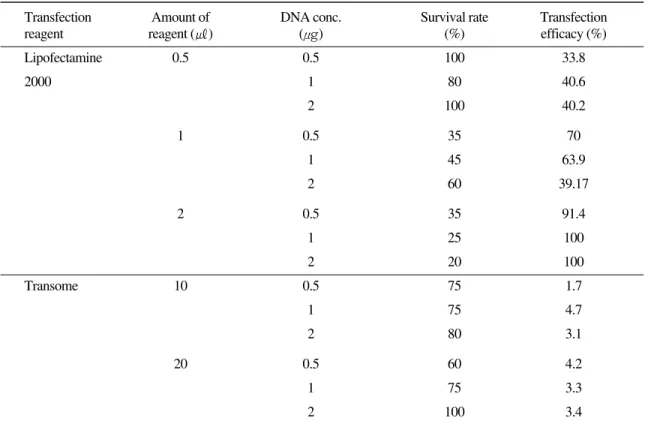
Optimum transfection efficiencies with low cytotoxicity were determined with BF-2 cells by Lipofectamine 2000 and Transome (Table 1). In the case of Lipofectamine 2000, cell viability was ranged from 20% to 100%, and it decreased with increasing the amount of reagent, but not influenced by concentration of the target DNA. Transfection efficacy of surviving cells was 33.8% to maximum 100%, and it increased with increasing concentra- tion of the target DNA. The maximum transfection efficacy (100%) were observed in use of 2 ㎕ Lipo- fectamine 2000 and 1 ㎍ or 2 ㎍ of DNA but sur- vival rates of those cells were 25% or less due to cell toxicity (Table 1). It was thus considered that these concentrations were impractical because sur- viving cell number was not enough for subculture.
In using of 0.5 ㎕ Lipofectamine 2000, transfection efficacy were relatively low (33.8, 40.6 and 40.2%
in 0.5, 1 and 2 ㎍ DNA, respectively), but the sur- vival rates of them were from 80% to 100%. From these results, it was considered that use of small amount of Lipofectamine 2000 was rather than large amount of one in consideration of cell survival rate, although use of 2 ㎕ of Lipofectamine 2000 and 1 ㎍ of DNA was best in the transfection effi- ciency of BF-2 cells.
Transfection efficiencies by Transome were all
less than 5% including the preliminary experiments
Se Ryun Kwon, Toyohiko Nishizawa and Mamoru Yoshimizu 378
Fig. 1. Construction strategies for LTR-GFP-LRT cassette harboring long terminal repeat (LTR) sequences at both 5、and 3、
ends of green fluorescent protein (GFP) gene.
Genome integration using non-replicable SnRV 379
Table 1. Cell viability on transfection reagent and DNA concentration
Transfection Amount of DNA conc. Survival rate Transfection
reagent reagent (㎕) (μ g) (%) efficacy (%)
Lipofectamine 0.5 0.5 100 33.8
2000 1 80 40.6
2 100 40.2
1 0.5 35 70
1 45 63.9
2 60 39.17
2 0.5 35 91.4
1 25 100
2 20 100
Transome 10 0.5 75 1.7
1 75 4.7
2 80 3.1
20 0.5 60 4.2
1 75 3.3
2 100 3.4
although toxicity of Transome to BF-2 cells was quite low level. It was also confirmed that Transfec- tion efficacy by Lipofectamine 2000 was obviously better than that by Transome under microscopic observation (Fig. 2).
Even if it is considered that there could be differ- ence in condition of commercial tranfection reagents based on liposomes between mammalian and fish cell lines, because fish cells have longer cell cycles than mammalian cells and lower optimal temperatures for growth (Rocha et al., 2005), Lipo- fectamine 2000 was better than Transome at least for transfection of BF-2 cells.
Inactivation of SnRV by UV irradiation
Integration of a target gene using retrovirus system requires that the target gene is transfected into the cells in which retroviral integrase gene is expressed, meaning that multiple plasmids for expression of the
target gene and integrase gene or single plasmid har- boring multiple expression cassettes of those genes are required. In the present study, we planed that inte- gration of a target gene was conducted using integrase derived from UV-inactivated retrovirus. Thereupon, dose of UV-irradiation for inactivation of SnRV was investigated prior to the integration experiment (Fig.
3). As a result, infectivity titer of SnRV was reduced within 3 min of irradiation by 250 ㎼/ cm
2of UV, and no infectivity titer was detected by more than 5 min of UV-irradiation. It was thus concluded that 5 min of UV- irradiation was enough to inactivate SnRV infectivity.
Energy of this UV irradiation was calculated as 7.5 × 10
4μ J/ cm
2, which is within the doses required to UV- inactivate fish pathogenic viruses (Yoshimizu, 2009).
Integration of LTR-GFP-LTR cassette using UV- inactivated SnRV
The LTR-GFP-LTR cassette was integrated using
Se Ryun Kwon, Toyohiko Nishizawa and Mamoru Yoshimizu 380
Fig. 3. Change of infectivity titer of SnRV according to duration of ultraviolet (UV) irradiation
Fig. 2. Microphotograph of BF-2 cells transfected with pCMS-EGFP using Transome (left) and Lipofectamine 2000 (right).
Microphotograph was taken under fluorescent microscopy 72 h after transfection.
UV-inactivated SnRV (Fig. 4). Fluorescence by GFP was expressed in BF-2 cells inoculated with UV-inactivated SnRV 3 and 5 times, but no fluores- cence was observed in the cells inoculated once and negative control cells. The present results suggested that integrase activity of SnRV could be left alive from UV-irradiation even if those virus lost infectiv- ity completely. It was also demonstrated that the tar- get gene was possible to be integrated into cell genome by using non-replicable retrovirus. No inte- gration reaction was observed in the cells transfect- ed with LTR-GFP-LTR cassette by once inoculation of UV-inactivated SnRV, and GFP expression rate
in BF-2 cells treated with UV-inactivated SnRV was as low as the transfection rate using Transome.
Those could be due to not enough efficiency by
Lipofectamine 2000 as described above and also
due to low virus titer of SnRV. Therefore, by
improving those points, higher integration efficacy
could be expected. We would improve pGFP -
LTRs by substituting the neomycin resistance gene
for GFP gene to select easily the cells inserted the
target gene. The demonstrated method for gene
integration by UV-inactivated retrovirus could be
useful and convenient tool for future analyses of
gene expression and function in fish cell lines.
Genome integration using non-replicable SnRV 381
요 약