CONTENTS
Ⅰ. INTRODUCTION
Ⅱ. MATERIALS AND METHODS
Ⅲ. RESULTS
Ⅳ. DISCUSSION
Ⅴ. CONCLUSIONS REFERENCES KOREAN ABSTRACT EXPLANATION OF FIGURES
Ⅰ. INTRODUCTION
The intercellular connections that characterise advanced forms of life would not be possible without a mechanism to remove individual cells that are no longer needed, or that function abnormally. Such a physiological cell death, in the abscense of inflammation, is achieved by apoptosis, a structurally distinct programmed cell death pathway.
1)The decision of a cell to undergo apoptosis can be influenced by a wide variety of regulatory stimuli. Recent evidence suggests that alterations in cell survival contribute to the
pathogenesis of a number of human diseases, including cancer, viral infections, autoimmune disease, neurodegenerative disorders, and AIDS.
2)Stress has long been believed to be related to a variety of diseases.
3,4)But the belief was based on somewhat metaphysical philosophy and was lacking firm scientific evidence. In those recent years, numerous studies have focused on the mechanism how stress is related to the physiological and pathological changes of various organs. And accumulating evidence supports the relationship between stress and the body at a concrete science level.
5)Recently, quite a few studies have suggested that stress is strongly associated with orofacial diseases.
In clinic, it has been particulary interesting to observe that the patients under stressful conditions commonly show signs and symptoms of the decreased function of the salivary glands. In the previous studies, Hong and Park demonstrated that stress can actually cause the cell death of the submandibular glands in rats.
6)According to Hong and Jung, Caspase-3 (a key player in the DNA fragmentation process and other morphological
Immunohistochemical Expression of Caspase-9 on Rat Submandibular Gland under Cold and Restraint Stress
Eun-Young Ahn, D.M.D., Kyung-Hwan Muhn, D.M.D., Jae-O Cho, D.M.D.,M.S.D.,Ph.D.
Yang-Hyun Chun, D.M.D.,M.S.D.,Ph.D., Jung-Pyo Hong, D.M.D.,M.S.D.,Ph.D.
Department of Oral Diagnosis & Oral Medicine, School of Dentistry, Kyung Hee University
* This study was supported by a grant of the Korea Health 21 R&D Project, Ministry of Health & Welfare, Republic of Korea, HMP-00-CH-10-0009.
changes associated with apoptosis) was expressed in the rat submandibular glands under restraint stress.
7)In order to have a better understanding of the pathologic mechanism in submandibular gland disease, emphasizing the significant role of stress in the disease, the present study was performed to investigate the expression of caspase-9, "initiator"
caspase of mitochondria-induced apoptosis, with respect to apoptosis in cells of the submandibular gland under cold and restraint stress by using immunohistochemistry.
Ⅱ. MATERIALS AND METHODS
1. Experimental animals and tissue preparation
Thirty-six Sprague-Dawley rats (8-week-old) were purchased from Dae-Han Experimental Animal Research Center, Seoul, Korea. They were maintained at 20-23℃ and fed ad libitum on a normal laboratory diet. The rats were divided into 3 groups: cold stress (15 rats), restraint stress (15 rats) and control groups (3 rats). The rats of cold stress group were immersed in cold water (at 4℃) for 6 min. once a day throughout the experiment.
The rats of restraint stress group were placed in the plastic cages during the experiment. The stress groups were then sacrificed at 0, 1, 3, 5 and 7 day of the experiment and the submandibular glands were collected immediately. The submandibular glands were frozen in dry ice, embedded in O.C.T.
compound (Polyfreeze
TM, cat, #19636, Polysciences, Inc.) in cryomolds, and stored at -70℃ until use.
2. Preparation of Frozen sections
Serial frozen sections were cut (4-8 ㎛), placed on positive-charged and RNA-free microscope slides, and stored at -70℃ until use. Frozen sections were allowed to come to room temperature (for 30 minutes) and fixed in cold acetone+alcohol(1:1) for 10 minutes, kept refrigerated (at 4℃). The
specimens were rinsed in three changes of 0.01 M Tris buffered saline (TBS ; pH is 7.4±0.05. Tris : 0.05 M, NaCl : 0.15 m, Lab Vision Corp., U.S.A) solution.
3. Immunohistochemistry
Antibody Caspase-9 p10 (H-83 ; Santa Cruz Biotechnology, U.S.A.) is a rabbit polyclonal antibody raised against a recombinant protein corresponding to amino acids 315-397 mapping within the carboxy terminus of caspase-9 of human origin and reacts with the p10 subunit and precursor of caspase-9 (also designated ICE-LAP6) of mouse, rat and human origin by Western blotting, immunoprecipitation and immunohistochemistry.
Immunostaining analysis was performed on the frozen section by the method as described previously.
Specimens were processed for immunostaining using ABC methods. After the fixation, specimens were washed three times in TBS and incubated with 3% H
2O
2for 10 minutes at room temperature. And then specimens washed three times with D.W.(distilled water). For the protein blocking, 150 μl normal serum in 10 ml TBS for 20 minutes at room temperature. And then specimens were incubated with the rabbit anti-caspase-9 in humidity at room temperature that was diluted 1:50 with concentrated-reagent diluents. After 2hrs., the specimen were rinsed with TBS solution and then were incubated with biotinylated goat anti-rabbit immunoglobulin for 40 minutes at room temperature.
specimens were then washed in TBS and incubated
with avidin-biotin-peroxidase complex (ABC kit ;
VECTASTAIN
®, Vector Lab., Burlingame, CA,
USA) for 40 minutes. Thereafter, the tissues were
rinsed with TBS solution and imunohistochemical
reactions were detected by color development using
3-amino-9-ethylcarbazole solution (AEC kit ;
Vector Lab., Burlingame, CA, USA). After washing
in D.W., specimens were counterstained with
hematoxylin and mounted in VECTASHIELD
®Mounting Medium for retaining tissues. Thereafter,
immunohistochemistrial reactions were examined under the microscope and photographed.
Ⅲ. RESULTS
To localize the expression of caspase-9 in the submandibular glands, the glands of each group were fixed and processed for immuno- histochemistry using the specific antibody against caspase-9.
In control group, caspase-9 immunoreactivity was detected slightly.
In the cold stress group, caspase-9 immuno- reactivity was detected at any time during the experiment and appeared to be strong 3 days after the application of the stress, and then gradually decreased.
In the cold stress group, destruction of submandibular gland cells was not detected.
In the restraint stress group, caspase-9 immuno- reactivity appeared to be strong 3 days after the application of the stress, and then gradually decreased. And immunoreactivity of caspase-9 was hardly observed at day 7.
In the restraint stress group, destruction of submandibular gland cells was detected 5 day after the application of the stress. The arragement of cell was irregular and a lot of acinar cells were replaced by ductal cells at day 5. At day 7, the number of submandibular gland cell was markedly decreased.
Ⅳ. DISCUSSION
There are many functions in the oral cavity. It is the site for ingestion food, for starting the process of digestion, for tasting and for articulating speech.
It also has protective properties against various noxious agents such as microorganisms and physical trauma. The structure of the oral mucosa and its interaction with saliva contribute to all these tasks.
8)The saliva is an extremely complex fluid containing mucin, enzymes, immunoglobulins, growth factors, etc. and has several functions. In
addition to moistening the oral tissues as an aid for speech, and functioning as an aqueous solvent necessary for taste, it acts as a masticatory wetting agent which assists swallowing. Certain glands initiate the digestive process by secreting an enzyme. Also, saliva has an antibacterial action which inhibits or prevents the onset of dental caries and other infections
It is well documented that saliva plays an important role in the development and maintenance of a healthy oral cavity.
9)The salivary glands, the major salivary glands including the parotid, the submandibular and the sublingual glands, and the several minor glands, produce and secrete saliva. Among the salivary glands, we chose the submandibular gland because there are many researches that submandibular glands have more steady function and highly sensitivity to cold stress than other salivary glands.
7)The salivary glands are controlled by the autonomic nervous system. Parasympathetic stimulation increases salivary flow, while anticholinergic drugs inhibit secretions. However, the sympathetic influence on the salivary gland function is more complex. Generally, sympathetic activity inhibits salivation, but the opposite case is also observed. Emotional stress induces a variety of responses in the body. In most instances alarm-like reactions and depression and anxiety inhibit, while pleasurable, relaxed sensations promote increase salivation.
9,10)The hypofunction of salivary gland is influenced by many factors ; xerogenic drugs, aging, radiation therapy, systemic or local diseases such as Sjögren syndrome and diabetes, dehydration, and psychogenic factors such as emotional stress.
11,12)Recently a few studies have suggested that stress is strongly associated with the function of salivary gland.
Stress can have an effect on human body which
is apt to maintain homeostasis and modulate three
main systems such as autonomic nervous system,
the hormonal system and immune response
system.
5)The events occurring in the brain in
response to stress, specifically at the hypothalamic-
pituitary-adrenal(HPA) axis, are similar to those occurring in response to an activated immune system. In both systems, the HPA axis is stimulated by corticotrophin-releasing factor(CRF), resulting in a down-regulation of immune function.
13)Therefore, stress-induced alterations in endocrine function and the potential pathophysiologic effects of such endocrine changes lead to a depletion of the subject's ability to resist stress, which ultimately leads to the disease process.
14)The relationship of stress to the orofacial area can be considered in four aspects
5): normal physiological and psychological functions of the mouth, stress-relieving orofacial activities, stress and dental treatment, and stress-related orofacial disorders and diseases.
Recently a few studies have suggested that stress is strongly associated with orofacial diseases. Chun and Hong
15)indicated that stress causes various forms of diseases in the region including orofacial psychosomatic diseases in which emotional stress appears to play a major role (lichen planus, aphthous stomatitis), orofacial diseases in which psychologic factors appear to play a role (erythema multiforme, benign mucous membrane pemphigoid, geographic tongue), orofacial infections where emotional stress is a significant predisposing factor (recurrent herpes labialis, acute necrotizing ulcerative gingivitis), orofacial lesions induced by neurotic habits inflicting trauma (biting of oral tissues, physical trauma with foreign objects, leukoplakia due to smoking, bruxism and clenching), neurotic orofacial symptoms (xerostomia, halitosis, burning mouth syndrome, altered or loss of taste perception, pain or discomfort with no tissue change), and orofacial pain induced by emotional stress (temporomandibular disorders, muscle tension headache, atypical odontalgia).
Depending on the type and degree of stress, the tissue response to it could be physiologic or pathologic. If stress exceeds adaptability of the tissue, it may affect various organs starting with alterations at cellular and molecular levels by cellular responses to injury: cell adaptive response, cell stress response (production of stress protein),
reversible cell injury, and ultimately irreversible cell injury such as apoptosis (programmed cell death).
16,17)Particularly, several reports have suggested that stress triggers apoptosis,
18,19,20)a process of genetically programmed alteration of cell structure that leads to failure of proliferation and eventual cell death.
16)In multicellular organisms, homeostasis is maintained through a balance between cell proliferation and cell death.
2)Such physiological cell death, in the abscense of inflammation, is achieved by apoptosis, a structurally distinct programmed cell death pathway.
1)Apoptosis is important in the regulation of normal cell population density, and may be one mechanism of deleting abnormal cells or cells that have been damaged by toxins, radiation injury, or other stimuli, and is thought to be responsible for numerous physiolosic and pathologic event including the following; the programmed destruction of cells during embryogenesis and metamorphosis, cell deletion in proliferating cell population, cell death in tumors, death of immune cells, pathologic atrophy of hormone-dependent tissues, cell death induced by cytotoxic T-cells, cell death produced by a variety of injurious stimuli, ect.
16)Morphologically, cells undergoing apoptosis demonstrate nuclear/cytoplasmic condensation and membrane protrusion. These initial changes are followed by fragmentation of the nuclear content and subsequent encapsulation of these fragment into
"apoptotic bodies" that are quickly and uno- btrusively consumed by adjacent cells.
21)Biochemically, apoptotic cells are characterized by reduction in the mitochondrial transmembrane potential, intracellular acidification, production of reactive oxygen species, externalization of phosphatidylserine residues in membrane bilayers, selective proteolysis of a subset of cellular proteins, and degradation of DNA into internucleosomal fragments.
22-27)Apoptosis can be influenced by a wide variety of
extrinsic and intrinsic stimuli, can be inhibited by
factors such as growth factor, estrogen and cysteine
protease inhibitors etc, and triggered by inducer of apoptosis such as TNF, Fas ligand, glucocorticoids, bacterial toxins, oncogenes etc. In the previous studies, Hong and Park demonstrated that stress can actually cause the cell death of the submandibular glands in rats.
6)According to Hong and Jung, Caspase-3 (a key player in the DNA fragmentation process and other morphological changes associated with apoptosis) was expressed in the rat submandibular glands under restraint stress. Recent evidence suggests that the failure of cells to undergo apoptotic cell death might be involved in the pathogenesis of a variety of human diseases, including cancer, autoimmune diseases, and viral infections. In addition, a wide number of diseases characterized by cell loss, such as neurodegenerative disorders, AIDS (acquired immunodeficiency syndrome), and osteoporosis, may result from accelerated rates of physiologic cell death.
2)Apoptosis is a morphologically and biochemically distinct form of cell death carried out by a genetically determined cell suicide program.
21,28,29)Several apoptosis-regulating genes identified in Caenorhabditis elegans have counterparts in humans, demonstrating that the basic mechanism of apoptosis is evolutionarily conserved.
30)Genetic studies of apoptosis in C. elegans have identified ced-3 and ced-4 as proapoptotic genes and ced-9 as an antiapoptotic gene F4 . Several mammalian homologs of CED-3, CED-4, and CED-9 have been identified. Apaf1 is homologous to CED-4 ; members of the Bcl2 family are homologous to CED-9 ; and at least 11 mammalian cysteine proteases (''caspases'') are homologous to CED-3.
31,32,33)Caspases are synthesized as zymogens or proenzymes (30-50 kDa) containing an N-terminal prodomain together with one large (20 kDa) and one small (10 kDa) subunit, and present in cells as inactive proenzymes,
34)but crystal structures suggest the active enzymes are heterotetramers composed of two small and two large subunits.
Importantly, specific aspartic acid cleavage sites exist between the prodomain of a caspase and each of its subunits, allowing for the possibility of a
caspase cascade, wherein one caspase could process and active another.
33,35)Caspases can be subdivided based on a number of criteria including phylogenetic analysis, substrate specificity, and the length of their prodomains. "Initiator" caspases, including caspase- 8 and -9, contain long prodomains which allow them to interact with specific adapter proteins. Such interactions bring initiator caspases in close proximity to one another and promote the activation of one zymogen by another.
36,37)Initiator caspases are responsible for either directly or indirectly activating various "effector" caspases, including caspase-3, -6, -7, which contain short prodomain.
Effector caspases cleave a number of structural and regulatory proteins and are directly responsible for many of the apoptotic features.
35)Among "effector"
caspases, Caspase-3 is activated by two sequential proteolytic events that cleave the 32 kDa precursor at aspartic acid residues to generate an active heterodimer of 20 kDa and 12 kDa subunits.
38)The activation might be autocatalytic, or it might occur via a caspase cascade, similar to the serine protease cascade in the blood clotting process. Such a cascade might amplify apoptotic signals leading to fast and irreversible apoptosis.
39)The activated caspases cleave a variety of target proteins, thereby disabling important cellular processes and breaking down structural components of the cell.
40)The targets of such cleavage events include poly(ADP- ribose) polymerase, sterol regulatory element binding proteins, retinoblastoma (RB) protein, nuclear lamins, DNA-dependent protein kinase, U1 70-K protein, and the large subunit of the DNA replication complex C.
39)In addition, activated caspases lead to cleavage of the 45 kDa subunit of DNA fragmentation factor (DFF-45). This cleavage activates a pathway leading to fragmentation of genomic DNA into nucleosomal fragments, a hallmark of apoptosis.
41)In terms of the hierarchical activation of caspase, caspase-8 and -9 appear to be the apical caspases activated in death receptor- and mitochondrial stress-induced apoptosis.
42)Caspase-8, a member of a mammalian caspase
family, has been demonstrated to play a key role in mediating Fas-induced apoptosis.
43)Cross-linking of the Fas receptor by engagement of the Fas ligand or agonistic antibodies results in the formation of so-called death-inducing signal complex (DISC), which includes adaptor protein FADD/MORT-1 and Caspase-8.
44)The formation of the DISC leads to the activation of Caspase-8, an initiator of the downstream apoptotic process that includes the activation of Caspase-3, -6, and -7 and mitochondrial damage.
34)Caspase 9, a key player in mitochondria-induced apoptosis, Apaf3, a 45 kDa protein ''also known as ICE-LAP-6 or Mch6'',
45,46)forms a multiprotein complex containing Apaf1 and cytochrome c.
39)A critical role of mitochondria in mediating apoptotic signal transduction pathway has been demonstrated recently.
47)Biochemical and structural changes of mitochondria in apoptosis include mitochondrial swelling, disruption of mitochondrial outer membrane, mitochondrial depolarization, and the release of cytochrome c.
47,48)Kroemer's group reported that pro-caspases-9 is localized in mitochondira and active caspase-9 is released during apoptosis.
49)The intermembranous space of the mitochondira seems to be a well-buffered compartment where dangerous molecules, including caspases, cytochrome c, and apoptosis-inducing factor (AIF), can be safely stored.
50,51)There is a recent evidence that caspase function may be modulated by physical interaction with Apaf1, cytochrome c (Apaf2), and Bcl2 family members.
31,52,53)The release of cytochrome c may trigger the interaction of Apaf1, a mammalian CED-4 homolog, and Caspase-9, which in turn results in the activation of Caspase-9.
31,39)Physical association of Caspase-9 and Apaf1 is mediated by the interaction of their respective caspase recruitment domain (CARD).
39)CARD is also found in other caspases with large prodomains, such as Caspase-4 and Caspase-8, which can associate with Apaf1 in mammalian cells.
52)Activated Caspase-9 then cleaves and activates pro- Caspase-3, an event that leads to the cleavage of other death substrates,
cellular and nuclear morphological changes, and ultimately, cell death. An active site mutant of Caspase-9 is able to block activation of Caspase-3 by Caspase-9.
39)Bcl2 and BclxL, two members of the Bcl2 family, prevent apoptosis induced by a variety of death stimuli.
54)The antiapoptotic protein Bcl-xL has also been shown to interact with Caspase-9 and Apaf1, resulting in the inhibition of Caspase-9 activation.
52,53)
They are localized mainly to the outer mitochondrial, nuclear membranes and endoplasmic reticular membrane through their carboxy-terminal membrane anchorage domains. Both of them contain the Bcl2 homology domains designated BH1, BH2, BH3, and BH4, all of which are essential for the antiapoptotic activity of Bcl2/BclxL.
54,55)Overex- pression of Bcl2/BclxL has been shown to block all apoptosis-induced mitochondrial changes.
43)Apoptotic signals are generally believed to be mediated through a hierarchy of caspase activation controlled by one of two distinct pathways that are associated with either caspase-8(i.e. death receptors) or caspase-9(i.e. mitochondria). Some studies demonstrated that caspase-8 can be activated in caspase-9 dependent manner. However, neither study was able to determine whether caspase-9-mediated activation of caspase-8 is physiologically relevant of merely a bystander effect.
56)The overall results indicate that submandibular gland cells were destroyed under restraint stress, and expression of caspase-9 was observed after the application of the stress.
So, we suggest that change of mitochondria is involved in the pathway which submandibular gland cell apoptosis caused by stress is mediated.
However, it is still unclear what mechanisms are involved in the relationship between caspase-8 (a key player of membrane receptor-induced apoptosis) and submandibular gland cell apoptosis under stress, and has to be elucidated in further studies.
Therefore, we will have to especially investigate
mitochondrial changes during stress-induced
apoptosis using transmission electron microscopy.
Ⅴ. CONCLUSIONS
In order to better understand the pathologic mechanism in submandibular gland disease, emphasizing the significant role of stress in the disease, the present study was performed to investigate the expression of caspase-9, "initiator"
caspase of mitochondria-induced apoptosis, with respect to apoptosis in cells of the submandibular gland under cold and restraint stress.
Eighteen Sprague-Dawley rats (8-week-old) were divided into 3 groups: cold stress (15 rats), restraint stress (15 rats) and control groups (3 rats).
The rats of cold stress group were immersed in cold water (4℃) for 6 min. once a day throughout the experiment. The rats of restraint stress group were placed in the plastic cages (150 x 50mm) during the experiment. The stress groups were then sacrificed by neck-dislocation at 0, 1, 3, 5 and 7 day of the experiment and the submandibular glands were collected immediately. The excised tissues were frozen and stored at -70℃ until use. The expression of caspase-9 in the submandibular gland tissues was observed by ABC immunohistochemistry method. The results were as follows:
1. In the control group, caspase-9 immunoreactivity was detected slightly.
2. In the cold stress group, caspase-9 immuno- reactivity was detected at any time during the experiment and appeared to be strong 3 day after the application of the stress, and then gradually decreased.
3. In the cold stress group, destruction of subman- dibular gland cells were not detected.
4. In the restraint stress group, caspase-9 immuno- reactivity appeared to be strong 3 day after the application of the stress, and then gradually decreased. And immunoreactivity of caspase-9 was hardly observed.
5. In the restraint stress group, destruction of submandibular gland cells were detected 5 days after the application of the stress. The arragement of cell was irregular and a lot of
acinar cells replaced by ductal cells at day 5. At day 7, the number of submandibular gland cell was markedly decreased.
The overall results indicate that submandibular gland cells were destroyed under restraint stress, and expression of caspase-9 was observed after the application of the stress. So, we suggest that change of mitochondria is involved in the pathway which submandibular gland cell apoptosis caused by stress is mediated. However, it is still unclear what mechanisms are involved in the relationship between caspase-8 (a key player of membrane receptor-induced apoptosis) and submandibular gland cell apoptosis under stress, and has to be elucidated in further studies. Thus, we will have to especially investigate mitochondrial changes during stress-induced apoptosis using transmission electron microscopy.
REFERENCES
1. Carson, D.A. and Ribeiro, J.M. : Apoptosis and disease. Lancet, 341: 1251-1254, 1993.
2. Thompson, C.B. : Apoptosis in the Pathogenesis and Treatment of disease. Science, 267:1456-1462, 1995.
3. Weiss, J.M. : Psychological factors in stress and diseases. Sci Am, 226:104-13, 1972.
4. Cohen, S. and Wills, T.A. : Stress, social support, and the buffering hypothesis. Psycho Bulletin, 98:310-57, 1985.
5. Selye, H. : Selye's guide to stress research. Vol. I, Canada, 1980, Van Nostrand Reinhold Ltd..
6. Park, H.K., Chun, Y.H., Lee, J.Y., Cho, H.G. and Hong, J.P. : Expression of Clusterin in the Salivary Gland under Restraint Stress. Kor J Stress Res, 6(2):33-44, 1998.
7. Chung, W.B., Jung, S.H., Chun, Y.H., Lee, J.Y. and Hong, J.P. : Caspase-3 expression in the submandibular gland of rat under restraint stress. J of Korean Academy of Oral Medicine, 25(3):265-278, 2000.
8. Blackwell : Oral mucosa in health and disease.
London, 1975, DOLBY AE..
9. Dobrosielski-Vergona, K. : Biology of the salivary glands. Boca Raton, 1993, CRC Press, Inc.
10. Lynch, M.A., Brightman, V.J. and Greenber, M.S. :
Burket's oral medicine. J.B. 9th ed, philadelphia, 1994, Lippincott Co..
11. Field, E.A., Longman, L.P., Bucknall, R. et al. : The establishment of a xerostomia clinic: a prospective study. Br J Oral Maxillofac Surg, 35:96-103, 1997.
12. Lewis, M.A. and Lamey, P.J. : Clinical oral medicine.
Oxford, 1993, Wright.
13. Black, P.H. : Psychoneuroimmunology : Brain and immunity. Sci. Am. Sci. Med., 2:16-25, 1995.
14. Auvenshine, R.C. : Psychoneuroimmuology and its relationship to the defferential diagnosis of temporomandibular disorders. Dental Clin. of Nor.
Am., 41(2):279-296, April 1997.
15. Chun, Y.H. and Hong, J.P. : Stress and Oral diseases.
Kor. Stress Res., 3(1):57-72, 1995.
16. Contran, R.S., Kumar, V., Robbins, S.L. and Schoen, F.J. : Robbins pathologic basis of disease. 5th ed, Philadelphia, 1994, W.B. Saunders Co..
17. Steven, A. and Lowe, J. : Pathology. London, 1995, Mosby. .
18. Tomei, L.D., Kiecolt-Glaser, J.K., Kennedy, S. and Glaser, R. : Psychological stress and phorbol ester inhibition of radiation-induced apoptosis in human peripheral blood leukocytes. Psychiatry Res, 33:59-71, 1990.
19. Sendo, F., Kato, T. and Yazawa, H. : Modulation of neutrophil apoptosis by psychological stress and glucocorticoid. Int J Immunopharmacol, 19(9-10):511- 6, 1997.
20. Koukalova, B., Kovarik, A., Fajkus, J. and Siroky, J.
: Chromatin fragmentation associated with apoptotic changes in tobacco cells exposed to cold stress.
FEBS LEtt, 414(2):289-92, 1997.
21. Kerr, J.F.R., Wyllie, A.H. and Currie, A.R. : Apoptosis : a basic biological phenomenon with wide-ranging implications in tissue kinetics. Br J Cancer, 26:239-257, 1972.
22. Gottlieb, R.A., Nordberg, J., Skowronski, E. and Babior, B.M. : Apoptosis induced in Jurkat cells by several agents is preceded by intracellular acidification. Proc. Natl. Acad. Sci., 93:654-658, 1996.
23. Hockenbery, D.M., Oltvai, X.N., Yin, X.M., Milliman, C.L. and Korsmeyer, S.J. : Bcl-2 functions in an antioxidant pathway to prevent apoptosis. Cell, 75:241-251, 1993.
24. Lazebnik, Y.A., Kaufmann, S.H., Desnoyers, S., Poirier, G.G. and Earnshaw, W.C. : Cleavage of poly(ADP-ribose) polymerase by a proteinase with properties like ICE. Nature, 371:346-347, 1994.
25. Martin, S., Reutelingsperger, C., McGahon, A. et al. : Early redistribution of plasma membrane phosphatidylserine is a general feature of apoptosis regardless of the initiating stimulus : Inhibition by overexpression of Bcl-2 and Abl. J. Exp. Med., 182:1545-1556, 1995.
26. Wyllie, A.H., Morris, R.G., Smith, A.L. and Dunlop, D.
: Chromatin cleavage in apoptosis : Association with condensed chromatin morphology and dependence on macromolecular synthesis. J. Pathol., 142:66-77, 1984.
27. Zamzami, N., Susin, S.A., Marchetti, P. et al. : Mitochondrial control of nuclear apoptosis. J. Exp.
Med., 183:1533-1544, 1996.
28. Horvitz, H.R., Shaham, S., and Hengartner, M.O. : The genetics of programmed cell death in the nematode Caenorhabditis elegans. Cold Spring Harb.
Symp. Quant. Biol., 59:377-385, 1994.
29. Jacobson, M.D., Weil, M., and Raff, M.C. : Programmed cell death in animal development. Cell, 88:347-354, 1997.
30. Vaux, D.L. : CED-4-the third horseman of apoptosis.
Cell, 90:389-390, 1997.
31. Zou, H., Henzel, W.J., Liu, X., Lutschg, A., and Wang, X. : Apaf-1, a human protein homologous to C.
elegans CED-4, participates in cytochrome c-dependent activation of caspase-3. Cell, 90:405-413, 1997.
32. Reed, J.C. : Bcl-2 and the regulation of programmed cell death. J. Cell. Biol., 124:1-6, 1994.
33. Cohen, G.M. : Caspases: the executioners of apoptosis.
Biochem. J., 326:1-16, 1997.
34. Salvesen, G.S., and Dixit, V.M. : Caspases:
intracellular signaling by proteolysis. Cell., 91:443-446, 1997.
35. Nicholson, D. W. : Caspase structure, proteolytic substrates, and function during apoptotic cell death.
Cell Death Differ., 6:1028-1042, 1999.
36. Muzio, M., Stockwell, B.R., Stennicke, H.R., Salvesen, G.S., and Dixit, V. M. : An induced proximity model for caspase-8 activation. J. Biol. Chem., 273:2926-2930, 1998.
37. Srinivasula, S.M., Ahmad, M., Fernandes-Alnemri, T., and Alnemri, E.S. : Autoactivation of caspase-9 by Apaf-1-mediated oligomerization. Mol. Cell., 1:949-957, 1998.
38. Nicholson, W.D., Ali, A., Thornberry, N.A. et al. : Identification and inhibition of the ICE/CED-3 protease necessory for mammalian apoptosis. Nature.
376:37-43, 1995.
39. Li, P., Nijhawan, D., Budihardjo, I. et al. : Cytochrome c and dATP-Dependent Formation of Apaf-1/
Caspase-9 Complex Initiates an Apoptotic Protease Cascade. Cell, 91:479-489, November, 1997.
40. Nicholson, W.D., and Thornberry, N.A. : Caspases:
killer proteases. Trends Biochem. Sci., 257:299-306, 1997.
41. Liu, X., Zou, H., Slaughter, C. and Wang, X. : DFF, a heterodimeric protein that functions downstream of caspase-3 to trigger DNA fragmentation during apoptosis. Cell, 89:175-184, 1997.
42. Shawn, B.B., Marion, M., Kelvin, C. and Gerald, M.C.
: Protein complexs activate dictinct caspase cascades in death receptor and stress-induced apoptosis. Exp Cell Res., 256:27-33, 2000.
43. Li, H., Zhu, H, Xu, C. J. and Yuan, J. : Cleavage of BID by Caspase 8 Mediates the Mitochondrial Damage in the Fas Pathway of Apoptosis. Cell, 94:491-501, August, 1998.
44. Kischkel, F.C., Hellbardt, S., Behrmann, I. et al. : Cytotoxicity-dependent APO-1 (Fas/CD95)- associated proteins form a death-inducing signaling complex (DISC) with the receptor. EMBO J., 14:5579-5588, 1995.
45. Duan, H., Orth, K., Chinnaiyan, A.M. et al. : ICE-LAP6, a novel member of the ICE/Ced-3 gene family, is activated by the cytotoxic T cell protease granzyme. B. J. Biol. Chem., 271:16720-16724, 1996.
46. Srinivasula, S.M., Fernandes-Alnemri, T., Zangrilli, J.
et al. : The Ced-3/interleukin 1beta converting enzyme-like homolog Mch6 and the lamin-cleaving enzyme Mch2alpha are substrates for the apoptotic mediator CPP32. J. Biol. Chem., 271:27099-27106, 1996.
47. Vander Heiden, M.G., Chandel, N.S., Williamson, E.K., Schmacker, P.T., and Thompson, C.B. : Bcl-xL regulates the membrane potential and volume homeostasis of mitochondria. Cell, 91:627-637, 1997.
48. Liu, X., Kim, C.N., Yang, J., Jemmerson, R., and Wang, X. : Induction of apoptotic program in cell free extracts: requirement for dATP and cytochrome C.
Cell, 86:147-157, 1996.
49. Susin S.A., Lorenxo H.K., Zamzami, N. et al. : Mitochondrial release of caspase-2 and -9 during the apoptotic process. J Exp Med., 189:381-394, 1999.
50. Susin, S.A., Lorenxo, H.K., Zamzami, N. et al. : Molecular characterization of mitochondrial apoptosis-inducing factor. Nature, 397:441-446, 1999, 51. Earnshaw, W.C: Apoptosis : a cellular poison
cupboard. Nature, 397:387-389, 1999.
52. Hu, Y., Benedict, M.A., Wu, D., Inohara, N. and Nunez, G. : Bcl-XL interacts with Apaf-1 and inhibits Apaf-1-dependent caspase-9 activation.
Proc. Natl. Acad. Sci. USA, 95:4386-4391, 1998.
53. Pan, G., O'Rourke, K. and Dixit, V.M. : Caspase-9, Bcl-XL, and Apaf-1 form a ternary complex. J. Biol.
Chem., 273:5841-5845, 1998.
54. Merry, D.E. and Korsmeyer, S.J. : Bcl-2 gene family in the nervous system. Annu. Rev. Neurosci., 20:245-267, 1997.
55. Huang, D.C.S., Adams, J.M. and Cory, S. : The conserved N-terminal BH4 domain of Bcl-2 homologues is essential for inhibition of apoptosis and interaction with CED-4. EMBO J., 17:1029-1039, 1998.
56. Damu, T., Jill, M.L. and Vincent, J.K. : Caspase-8 activation and bid cleavage contribute to MCF7 cellular execution in a caspase-3-dependent manner during staurosporine-mediated apoptosis. J. Biol.
Chem., 275(13): 9303-9307, 2000.
Corresponding Author : Jung-Pyo Hong, Professor, Department of Oral Diagnosis & Oral Medicine, School of Dentistry, Kyung Hee University, 1 Hoegi-Dong, Dongdaemun-Ku, Seoul 130-701, Korea
국문초록
구속과 한냉스트레스에 의한 흰쥐 악하선 조직에서의 Caspase-9 변화에 관한 면역조직화학적 연구
경희대학교 치과대학 구강내과학 교실
안 은 영․문 경 환․조 재 오․전 양 현․홍 정 표
세포자사는 더 이상 기능을 하지 못하거나 이상기능을 나타내는 세포를 염증 반응 없이 제거하여 생체의 항상 성을 유지시키기 위한 기전이다. 세포자사가 과도하거나 부족할 경우 인체는 질병에 이환 될 수 있으며, 이는 스 트레스에 의해서도 유발될 수 있다. 이에 본인은 구속과 한냉 스트레스시 흰쥐 악하선 세포의 세포자사 과정을 구명하기 위하여 세포자사 과정 중 중요하게 작용하는 단백 분해효소인 caspase-9 의 발현을 관찰하고자 하였다.
웅성 흰쥐 (Sprague-Dawley, 8주) 33마리를 하루 5분간 한냉 스트레스를 부여한 군과 실험 전 기간에 걸쳐 구속 스트레스를 가한 군으로 나누고 실험 후 각 군별로 즉일, 1일, 3일, 5일, 7일에 희생시켰으며 실험동물의 악하 선을 절취하여 동결절편을 제작하였고 caspase-9에 대하여 ABC 면역조직화학법을 시행하여 관찰하였다.
1. 한냉 스트레스 군에서는 전 실험군에서 caspase-9이 발현되었으며, 3일군까지는 증가되었으나 점차 감소되었 다.
2. 한냉 스트레스 군에서는 악하선 선포세포의 파괴양상이 관찰되지 않았다.
3. 구속 스트레스 군에서 caspase-9의 발현은 구속 3일군까지 증가되었으나 5일군부터 감소되기 시작하여 7일군 에서는 거의 발현되지 않았다.
4. 구속 스트레스 군에서는 5일군에서 세포의 파괴가 진행되어 세포의 배열이 불규칙해지고 많은 선포세포가 도 관세포로 대치되었으며, 7일군에서는 보다 현저하여 세포의 수가 매우 감소되었다.
본 연구에서 구속스트레스시 세포의 파괴가 진행되고 caspase-9의 발현이 관찰되었던 바 구속 스트레스에 의
한 악하선의 세포자사와 사립체의 변화가 관련이 있다고 사료된다.
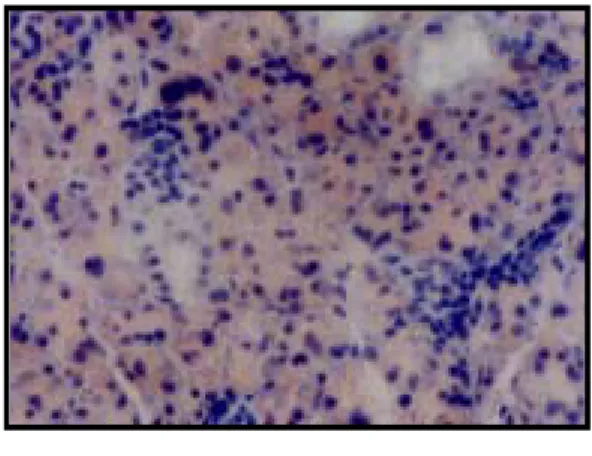
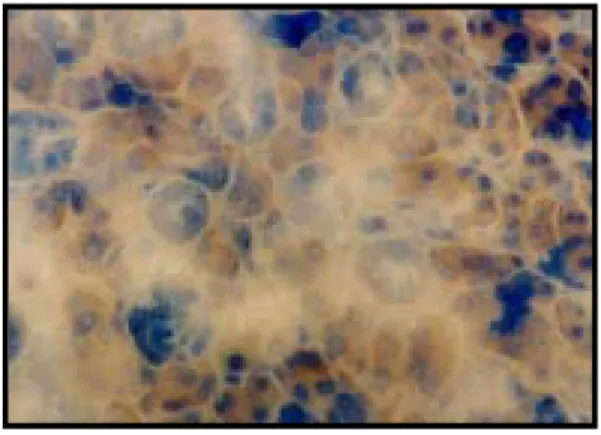
EXPLANATION OF FIGURES
Fig. 1. Immunohistchemistry microscopy of caspase-9 in the submandibular gland of the control group (X200).
Fig. 2. Immunohistochemistry microscopy of caspase-9 in the submandibular gland of the rat under cold stress immediatly (X200) after the application of the stress.
Fig. 3. Immunohistochemistry microscopy of caspase-9 in the submandibular gland of the rat under cold stress 1 day (X200) after the application of the stress.
Fig. 4. Immunohistochemistry microscopy of caspase-9 in the submandibular gland of the rat under cold stress 3 days (X200) after the application of the stress.
Fig. 5. Immunohistochemistry microscopy of caspase-9 in the submandibular gland of the rat under cold stress 5 days (X200) after the application of the stress.
Fig. 6. Immunohistochemistry microscopy of caspase-9 in the submandibular gland of the rat under cold stress 7 days (X200) after the application of the stress.
Fig. 7. Immunohistochemistry microscopy of caspase-9 in the submandibular gland of the rat of the restraint stress group immediatly (X200) after the application of the stress. .
Fig. 8. Immunohistochemistry microscopy of caspase-9 in the submandibular gland of the rat under restraint stress 1 day (X200) after the application of the stress.
Fig. 9. Immunohistochemistry microscopy of caspase-9 in the submandibular gland of the rat under restraint stress 3 days (X200) after the application of the stress.
Fig. 10. Immunohistochemistry microscopy of caspase-9 in the submandibular gland of the rat under restraint stress 5 days (X200) after the application of the stress.
Fig. 11. Immunohistochemistry microscopy of caspase-9 in the submandibular gland of the rat under restraint stress 7 days (X200) after the application of the stress.
Fig. 1. Fig. 2.
Fig. 3. Fig. 4.
Fig. 5. Fig. 6.
Fig. 7. Fig. 8.
Fig. 9. Fig. 10.
Fig. 11.