Abstract
The Effects of Daecheongryong-tang on Transcription Factors and Adipogenic Genes during 3T3-L1 Differentiation
Lee Jun Moon, Cho Sung Woo*, Kang Kyung Hwa**, Lee Seung Yeon, Yu Sun Ae Dept.of Pediatrics, College of Oriental Medicine, Dong Eui University
*Dept.of Rehabilitation Medicine, College of Oriental Medicine, Dong Eui University
**Dept.of Oriental Physiology, College of Oriental Medicine, Dong Eui University
Objectives
The purpose of this study is to investigate the effects of Daecheongryong-tang (DCRT) on the adipogenesis in 3T3-L1 preadipocytes.
Methods
3T3-L1 preadipocytes were differentiated with adipogenic reagents by incubating for 2 days in the absence or presence of DCRT ranging 0.25 and 2%. The effect of DCRT on adipogenesis was examined by Oil red O staining, and the protein, RNA, and RT-PCR were measured.
Results
Our results showed that DCRT decreased the TG content by ORO staining. To elucidate the mechanism of the effects of DCRT on lowering TG content in 3T3-L1 adipocytes, we examined the DCRT modulate expressions of transcription factors to induce adipogenesis and adipogenic genes which is related to the regulation of accumulation of lipids.
As a result, the expression of SREBP1, C/EBPβ, C/EBPδ, C/EBPα, and PPARγ genes, which induce the adipose differentiation and adipose-specific aP2, adipsin, LPL, CD36, TGF-β and adiponectin genes which regulates fat formations, were decreased. In addition, DCRT reduced the expression of iNOS and IL-6 in 3T3-L1 adipocytes, resulting in inflammation.
Conclusions
DCRT could regulate transcript factor related to induction of adipose differentiation, inhibit the accumulation of lipids and expression of the adipogenic genes.
Key words : Daecheongryong-tang (DCRT), Adipocytes, Differentiation, Transcript factor, Adipogenic genes
대청룡탕이 지방세포 분화기전에 미치는 영향
이준문․조성우*․강경화**․이승연․유선애
동의대학교 한의과대학 소아과교실
*동의대학교 한의과대학 재활의학과교실
**동의대학교 한의과대학 생리학교실
■투 고 : 2010년 11월 26일, 수 정 : 2010년 12월 14일, 채 택 : 2010년 12월 16일
■교신저자 : 유선애, 울산광역시 남구 신정동 479-17 동의대부속울산한방병원 (Tel : 052-226-8107, E-mail : [email protected])
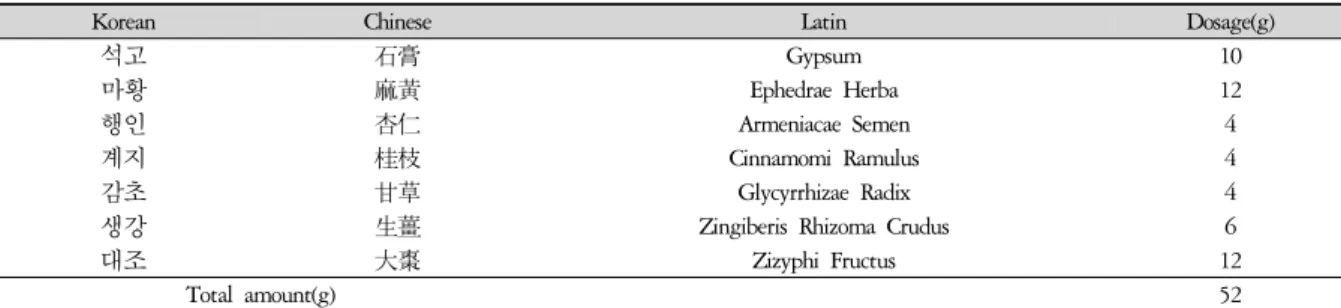
Korean Chinese Latin Dosage(g)
석고 石膏 Gypsum 10
마황 麻黃 Ephedrae Herba 12
행인 杏仁 Armeniacae Semen 4
계지 桂枝 Cinnamomi Ramulus 4
감초 甘草 Glycyrrhizae Radix 4
생강 生薑 Zingiberis Rhizoma Crudus 6
대조 大棗 Zizyphi Fructus 12
Total amount(g) 52
Table 1. The Composition of DCRT
Ⅰ. 緖 論
비만은 체내에서 요구되는 에너지의 양보다 많은 에 너지를 섭취하고 남은 에너지가 지방으로 바뀌어 체내 에 축적되어 발생되는 일종의 병리현상으로 체내의 지 방조직이 과다하게 증가된 상태를 말하며, 비만 그 자 체도 질병이 되지만 여러가지 성인병의 위험을 증가시 키는 요인이 되기도 한다1).
지방세포는 전구지방세포의 증식과 분화과정을 통 해 성숙되어 형성되는데, 전구지방세포에서 형태학적 으로나 생화학적으로 완벽히 성숙된 지방세포로 분화 되는 과정에는 인슐린 등을 포함한 호르몬 자극과 지 방세포 유전자의 조절부위에 중요한 전사인자가 활성 화되어야 한다2).
大靑龍湯은《傷寒論》3)에 “太陽中風 脈浮緊 發熱 惡寒 身疼痛 不汗出而煩躁者 大靑龍湯主之”, “傷寒 脈 浮緩 身不疼 但重 乍有輕時 無少陰證者 大靑龍湯發之”
라고 기재되어 있다. 임상적 응용은 流行性感冒, 肺炎, 氣管支哮喘, 流行性髓膜炎, 痲疹, 胸膜炎, 急性關節炎, 丹毒, 急性皮膚病性浮腫 등의 急性熱病 初期의 高熱에 表寒과 內熱, 煩躁가 보일 때 응용할 수 있다4).
이에 저자는 임상에서 호흡기 질환을 치료하기 위해 大靑龍湯을 복용하는 환자들에게 체중감량의 효과가 나타나 大靑龍湯이 비만조절에 대한 응용가능성과 근 거를 살펴보고자 하였다.
大靑龍湯에 대한 실험적 연구로는 호흡기 점액의 분비에 미치는 영향과 기도배상세포에서의 mucin분비 에 미치는 영향에 관한 연구가 보고되어 있었으나5, 6), 비만에 대한 연구는 찾아볼 수 없었다. 이에 저자는 대 청룡탕의 지방세포의 증식과 지방세포에서 분비되는 물질과 그 분화 기전 조절에 대하여 살펴보기 위하여, 3T3-L1 전구지방세포를 이용하여 지방세포 분화억제, 지방세포 분화 관련 전사인자와 유전자 발현조절 및
지방세포 분화관련 신호전달경로의 활성 조절에 관한 효과를 조사한 바 유의한 결과를 얻었기에 보고하는 바이다.
Ⅱ. 材料 및 方法
1. 재료
3T3-L1 mouse preadipocytes는 American Type Culture Collection (ATCC, USA)에서 분양받아 사용하였고, Dulbecco's Modified EaDCRT Media (DMEM), bovine calf serum (BCS), fetal bovine serum (FBS), penicillin-streptomycin mixture 및 dimethylsulfoxide(DMSO)는 GibcoBRL (USA)로 부터 구입하였다. Insulin, isobutylmethylxanthine(IBMX), dexamethasone (DEX), tetrazolium bromide salt(MTT), di- methylsulfoxide(DMSO), bovine serum albumin (BSA)는 Sigma (USA)로부터 구입하였다.
2. 시료의 준비
大靑龍湯(Daecheongryongtang, DCRT)은 傷寒論3)에 의거하여 한 첩의 분량은 Table 1과 같이 하였다. 약재 들은 광명당 (부산, 한국)에서 구입, 정선하여 사용하였 으며, 20첩 분량인 1040 g을 증류수로 水洗하고 2 ℓ의 증류수를 加하여 3시간 동안 가열 추출하였다. 추출된 용액을 원심분리를 통하여 상청액을 분리하고, 0.2 μm 여과지에서 여과하여 rotary evaporator로 200 ㎖가 되 도록 減壓濃縮하였다. 실온에서 냉각시킨 뒤 감압필터 를 사용하여 불순물을 제거하고 고압멸균하여 -20oC에 보관하였다가 실험직전 Phosphate buffered saline(PBS) 에 희석하여 사용하였다 (Table 1).
3. 세포배양 및 분화
지방세포의 분화에 관한 실험의 경우, 3T3-L1 전구지
Group Treatment
Normal group Undifferentiated cells
Control group Differentiated cells were treated with PBS
DCRT1 group Differentiated cells were treated with DCRT 1 mg/ml
DCRT2 group Differentiated cells were treated with DCRT 2 mg/ml
Table 2. The Experimental Groups
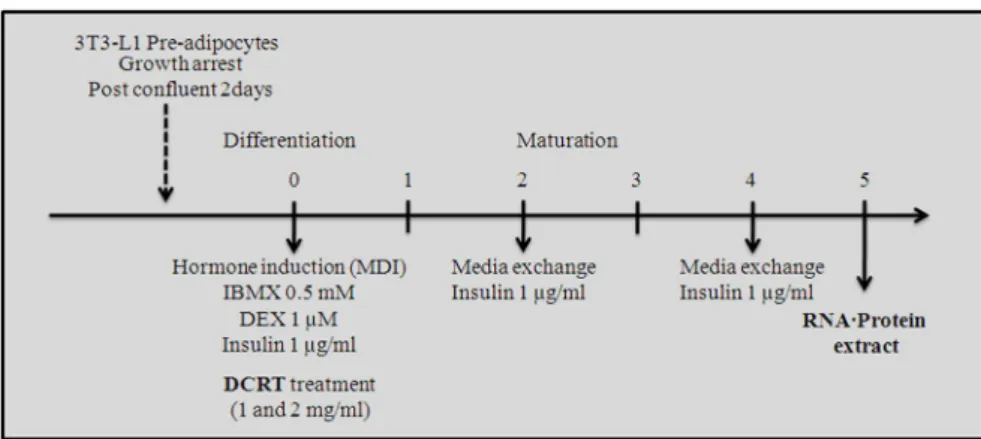
The 2-day post-confluent 3T3-L1 preadipocytes were treated with MDI (0.5 mM IBMX, 1 μM DEX and 1 μg/ml insulin) for 48 hrs.
Cells were then replaced with maintenance media containing 1 μg/ml insulin every 2 days for 5 days. To investigate effect of DCRT on adipogenesis, the differentiating 3T3-L1 cells were treated every 2 days with DCRT for 5 days.
Scheme 1. The experimental scheme of differentiation of 3T3-L1 preadipocytes
방세포는 5% CO2, 37℃ 배양기에서 DMEM(10% FBS, 100 unit/ml of penicillin G sodium, 100 μg/ml of streptomycin sulfate)배양하였다. 100% confluent 해졌을 때 0.5 mM IBMX, 1 μM DEX와 1 μg/ml insulin의 분화유도물질(MDI) 이 포함된 DMEM으로 교환하여 2~3일 동안 분화를 유도하였으며, 그 후 매 2일마다 1 μg/ml insulin이 포함된 DMEM으로 교환하였다. 분화 5 일 후 세포는 분석을 위해 사용되었다(Scheme 1). 3T3-L1 전구지방세포에 0.5 mM IBMX, 1 μM DEX와 1 μg/㎖ insulin을 처리하여 분화를 유도한 것을 control group으로, control group에 1 ㎎/㎖과 2 ㎎/㎖의 대청룡탕을 분화유도 2일 동안 처리 한 것을 실험군으로 하였다 (Table 2).
4. MTT Assay
3T3-L1 전구지방세포에 대한 대청룡탕의 독성 여부 를 알아보기 위하여 MTT assay를 시행하였다. 96 well plate(Corning, USA)에 1 × 104 cells/well의 농도로 세포 를 DMEM 배양액에 분주하여 24 시간동안 안정화시킨 후, DCRT를 농도별(0, 0.25, 0.5, 1 및 2 ㎎/㎖)로 처리하 여 1, 24 및 48 시간동안 반응시켰다. 배양액을 제거 한 후 5 ㎎/㎖의 MTT를 각 well에 넣고 잘 섞어 주었다.
최대 4 시간 37℃ incubator에서 배양한 후 tetrazolium bromide salt를 제거하고 각 well에 DMSO를 100 ㎕ 씩
분주하여 생성된 formazan이 잘 녹을 수 있게 충분히 흔들어 모두 녹인 후 ELISA(Molecular Devices, USA)를 사용하여 540 nm에서 흡광도를 측정하였으며, 3회의 측정으로 그에 대한 평균값과 표준 오차를 구하였다.
5. Oil Red O 염색 및 정량
세포내 지방구 생성을 확인하기 위하여 Oil red O 염색을 실시하였다. 배양된 세포는 phosphate-buffered saline(PBS)로 세척한 후 3.7% formalin으로 10 분간 고 정하고 deionized water로 세척하여 Oil red O를 처리한 후 실온에서 30 분간 염색하였다. 그 후 염색액을 제거 하고 deionized water로 3 회 세척하여 염색된 세포를 microscopic image(Olympus, Japan)로 관찰하였다. 또한 정량을 위해 DMSO를 가하여 지방을 추출한 후 micro- plate reader(Molecular Devices, USA)를 사용하여 490 nm 에서 흡광도를 측정하였다.
6. RNA 추출
Cell에 TRizol을 넣고 2분간 방치하였다가 세포를 떼 어낸 후 chloroform을 넣고 10 초간 강하게 vortexing하 고 4℃ 14,000 rpm에서 15 분간 원심분리하였다. 원심 분리 후, 상층액을 취하여 동량의 isopropanol을 혼합하 여 흔들어 준 후 실온에 10 분간 방치하였다. 그 후 4℃
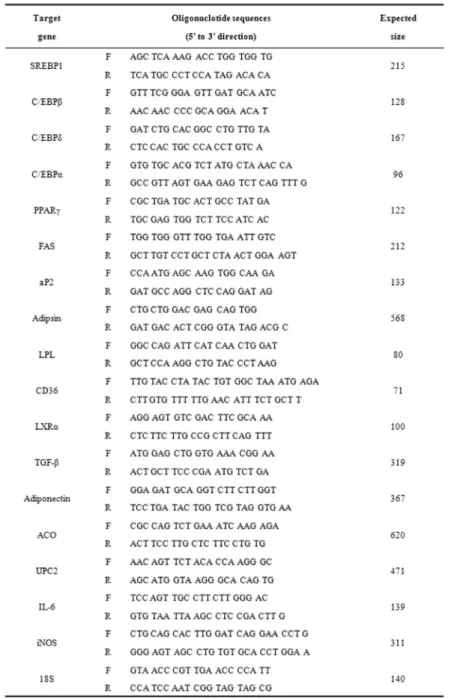
F: forward, R: reverse
Table 3. The Primers for RT-PCR
14,000 rpm에서 10 분간 원심분리하여 상층액을 제거 하고 75% ethanol을 넣고 섞어준 후 4℃ 7,500 rpm에서 5 분간 원심분리하였다. RNA pellet은 10 분간 실온에 서 건조시키고 DEPC(diethyl pyrocarbonate)-DW에 녹여 260 nm에서 흡광도를 측정하여 RNA 농도를 계산하였 다.
7. RT-PCR
각 1 mg의 RNA를 취하여 RT-PCR kit (Bioneer, Korea)를 사용하여 45℃에서 30 분, 94℃에서 5 분간
반응시킨 후 94℃에서 30 초간 denaturation시키고, 55-62℃에서 30 초간 annealing시킨 다음, 72℃에서 1 분간 extension시키는 cycle을 30-35 회 반복한 뒤, 마지 막 extension은 72℃에서 5분간 PCR machine (GeneAmp, PCR system 9700, USA)에서 수행하였다. 각 PCR prod- ucts는 0.5×TBE buffer (45 mM Tris-borate, 1mM EDTA) 를 사용하여 2% agarose gel에 loading하여 100 V 조건 에서 30 분간 전기영동한 후 etidium bromide 염색을 한 후 분석하였다. 각각의 primer는 Bioneer (Bioneer Co., Korea)에 의뢰하여 합성하였다(Table 3).
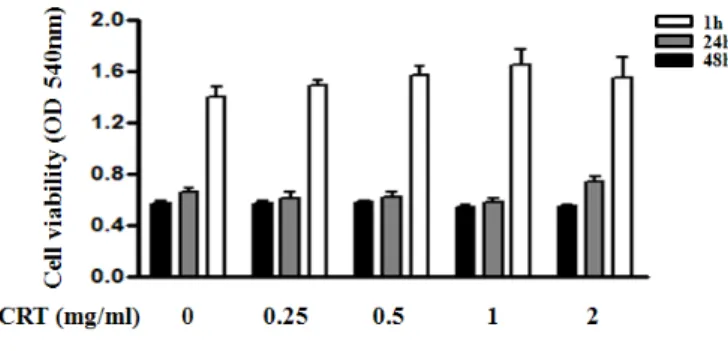
Fig. 1. Effects of DCRT on the cell viability in 3T3-L1 cells
3T3-L1 preadipocytes were treated with the indicated concentrations of DCRT for 1, 24 and 48 hrs. Cell viability was evaluated using a colorimetric assay based on MTT assay. Values are means ± SD. of three independent experiments.
8. Protein 추출
Cold PBS로 세척하여 수거한 세포에 protease cocktail 을 첨가한 RIPA lysis buffer (0.5 M Tris-HCl, pH 7.4, 1.5 M NaCl, 2.5% deoxylcholic acid, 10% NP-40, 10 mM EDTA)를 첨가하여 매 5 분간 vortex하여 얼음에 30 분 간 방치하였다. 그 후 12,000 rpm, 4℃에서 20 분간 원 심분리하여 상청액을 회수하여 단백질 양을 정량하 였다.
9. Western blot
각 lysate 50 ㎍을 취하여 SDS loading buffer (100 mM Tris-HCl, pH 6.8, 20% glycerol, 200 mM β-mercatoetha- nol, 10% SDS, 0.2%(w/v) bromophenol blue)와 섞어 10%
SDS-polyacrylamide gel에 loading하여 150 V에서 1 시간 30 분 동안 전기영동하여 단백질을 분리시켰다. 분리 된 단백질을 PVDF membrane (Amersham, Sweden)에 1 시간 transfer시킨 후 10% skim milk가 첨가된 TBST buffer (10 mM Tris-HCl, pH 8.0, 150 mM NaCl, 0.1%
Tween20)로 실온에서 1 시간 blocking시킨 후 1 차 anti- body를 1:500~1:2,000 희석하여 첨가한 후 반응시켰 다. TBST buffer로 3 회 세척한 후 2 차 antibody를 1:2,000으로 희석하여 1 시간 이상 반응시켰다. TBST buffer로 3 회 세척 후 ECL detection kit (Amersham, Sweden)를 이용하여 X-ray film에 노출시켜 현상하였다.
10. 통계분석
통계분석은 SPSS 12.0K for Windows 통계 프로그램 패키지를 사용하여 평균치 ± 표준편차로 나타내었고 유의수준은 p <.05로 하였다. 각 실험군 간의 통계학적 분석은 One way-ANOVA와 Dunnett's Multiple Comparison Test 검정을 실시하였다.
Ⅲ. 結 果
1. 지방세포 분화억제 효과
1) 지방전구세포 증식에 미치는 영향
3T3-L1 전구지방세포에 0, 0.25, 0.5, 1 및 2 ㎎/㎖의 DCRT을 1시간, 24시간과 48시간동안 처리하였다.
아무런 처리를 하지 않은 군에서 1시간동안 처리한 결과 0.57±0.03, 24시간동안 처리한 결과 0.65±0.08, 48 시간동안 처리한 결과 0.57±0.03의 흡광도를 나타내 었다.
0.25 ㎎/㎖의 DCRT를 처리한 군에서 1시간동안 처 리한 결과 0.57±0.05, 24시간동안 처리한 결과 0.61±0.11, 48시간동안 처리한 결과 0.57±0.05의 흡광 도를 나타내었다.
0.5 ㎎/㎖의 DCRT를 처리한 군에서 1시간동안 처리 한 결과 0.58±0.04, 24시간동안 처리한 결과 0.61±0.12, 48시간동안 처리한 결과 0.58±0.04의 흡광도를 나타내 었다.
1 ㎎/㎖의 DCRT를 처리한 군에서 1시간동안 처리한 결과 0.54±0.04, 24시간동안 처리한 결과 0.57±0.09, 48시 간동안 처리한 결과 0.54±0.04의 흡광도를 나타내었다.
2 ㎎/㎖의 DCRT를 처리한 군에서 1시간동안 처리한 결과 0.54±0.04, 24시간동안 처리한 결과 0.74±0.11, 48시 간동안 처리한 결과 0.54±0.04의 흡광도를 나타내어 대 청룡탕의 농도와 처리시간에 따라 세포의 증식이 저해 되지 않았음을 확인할 수 있었다(Fig. 1).
2) 지방세포의 형태학적 변화에 미치는 영향 100% 가득 찬 3T3-L1을 2 일 동안 분화를 유도하고 분화 양상을 현미경으로 관찰하여 세포 형태의 변화 를 관찰한 결과 전구지방세포의 세포밀도가 100%
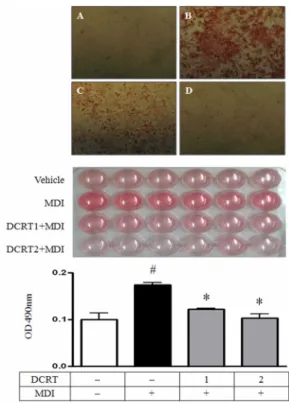
Fig. 2. Effect of DCRT on the microscopic morphological changes of 3T3-L1 preadipocytes differentiation The 2-day post-confluent 3T3-L1 preadipocytes(day 0) were treated with MDI (0.5 mM IBMX, 1 μM DEX and 1 μg/ml insulin) for 48 hrs in the absence or presence of indicated concentrations of DCRT. Cells were then replaced with maintenance media containing 1 μ g/ml insulin and DCRT for every 2 days. Accumulated lipid droplets in adipocytes were observed 1 day, 2 days, and 5 days after the initiation of differentiation and morphological changes associated with adipogenesis were photographed using Inverted Microscope (Carl Zeiss, Germany). (A) Undifferented cells, (B) Control was treated with MDI to induce differentiation, (C and D) MDI and 1 ㎎/㎖ and 2 ㎎/㎖ of DCRT were treated.
Fig. 3. Effects of DCRT on the adipogenesis and lipid accumulation of 3T3-L1 adipocytes
The 2-day post-confluent 3T3-L1 preadipocytes (day 0) were differentiated into adipocytes in the absence or presence of indicated concentrations of DCRT. Adipocytes were stained with Oil red O to determine the degree of adipocyte differentiation. Stained intracellular lipid was solubilized in isopropanol and absorbance was measured to evaluated lipid content. Values are means ± SD.
# p<.05 vs. undifferented cells, * p<.05 vs. control.
(A) Undifferented cells, (B) Control was treated with MDI to induce differentiation, (C and D) MDI and 1 ㎎/㎖ and 2 ㎎/㎖ of DCRT were treated.
Fig. 4. Effects of DCRT on the expression of SREBP1 in mRNA level
The 2-day post-confluent 3T3-L1 preadipocytes (day 0) were differentiated into adipocytes in the absence or presence of indicated concentrations of DCRT for 1 day. The mRNA levels were determined by RT-PCR as described in ‛Material and Methods’.
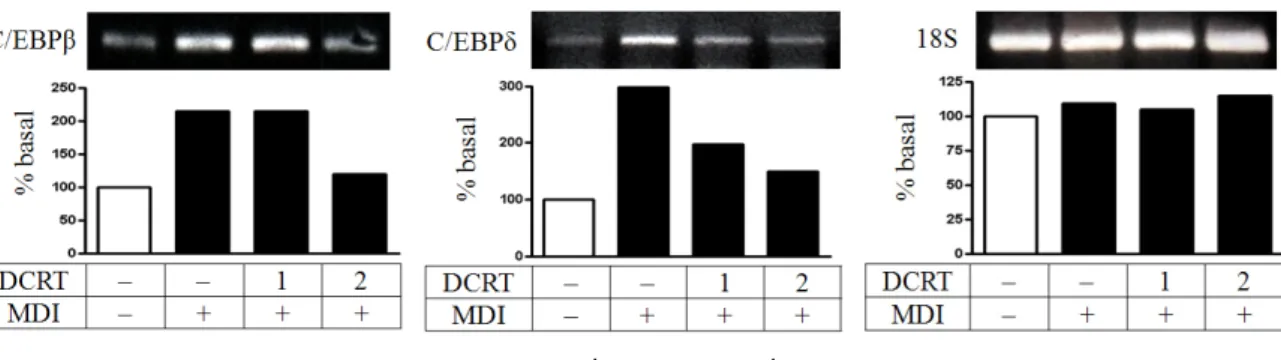
Fig. 5. Effects of DCRT on the expression of C/EBPβ and C/EBPδ in mRNA level
The 2-day post-confluent 3T3-L1 preadipocytes (day 0) were differentiated into adipocytes in the absence or presence of indicated concentrations of DCRT for 2 days. The mRNA levels were determined by RT-PCR as described in ‛Material and Methods’.
confluent된 세포형태는 길게 뻗은 섬유아세포의 형태 를 보여주었으나 (A), 지방세포로 분화 유도한 1 일, 2 일과 5 일째 세포의 형태는 시간이 지남에 따라 세포 모양이 둥글어지고 세포질 내 지방구들이 점차 많이 형성됨을 현미경으로 관찰할 수 있었다 (B). 반면 DCRT group은 농도에 의존적으로 세포내 지방구의 생성이 억제되는 것을 관찰할 수 있었다 (C and D, Fig.
2).
3) 지방세포 내 중성지방 생성에 미치는 영향 Oil red O를 이용하여 세포 염색을 시행하여 지방세 포의 형태를 관찰하고 세포 내 중성지방의 양을 정량 하였다. 관찰한 결과 control group에 비해 1 ㎎/㎖과 2
㎎/㎖의 DCRT group에서 지방세포 내 지방구의 형성 이 억제되어 있는 것으로 관찰되었다. 또한 지방세포 내 염색된 중성지방의 양을 정량한 결과 normal group 의 흡광도는 0.10±0.03로 나타났으며, control group에 서의 흡광도는 0.17±0.01로 나타났다. 1 ㎎/㎖과 2 ㎎/
㎖ DCRT group의 흡광도는 각각 0.12±0.01와 0.10±0.02로 지방생성을 유의하게 억제하였다 (Fig. 3).
2. 지방세포 분화 관련 전사인자 및 유전자 발현 조절 효과 1) 지방분화 초기 전사인자 발현에 미치는 영향
SREBP1의 발현 양상을 살펴보면 SREBP1 mRNA는 분화 1 일째 control group은 normal group에 비하여 발현이 증가하 였고, DCRT group은 control group에 비해 농도에 의존적으로 발현 감소를 보였다 (Fig. 4).
C/EBPβ와 C/EBPδ의 발현 양상을 살펴보면 C/EBPβ와 C/EBPδ mRNA는 분화 2 일째 control group은 normal group에 비하여 발현이 증가하였고, DCRT group은 control group에 비 해 농도에 의존적으로 발현 감소를 보였다 (Fig. 5).
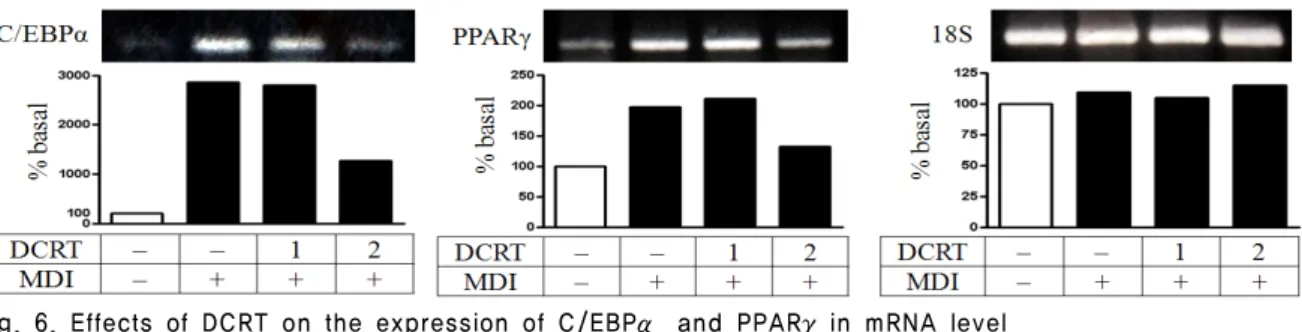
C/EBPα와 PPARγ의 발현 양상을 살펴보면 C/EBPα와 PPARγ mRNA는 분화 2 일째 control group은 normal group에 비하여 발현이 증가하였고, DCRT group은 C/EBPα mRNA는 control group에 비해 농도에 의존적으로 발현 감소를 보였으 며, PPARγ mRNA는 control group에 비해 2 ㎎/㎖농도에서 발현 감소를 보였다 (Fig. 6).
2) 지방합성 유전자 발현에 미치는 영향
LXRα의 발현 양상을 살펴보면 LXRα mRNA는 분화
Fig. 6. Effects of DCRT on the expression of C/EBPα and PPARγin mRNA level
The 2-day post-confluent 3T3-L1 preadipocytes (day 0) were differentiated into adipocytes in the absence or presence of indicated concentrations of DCRT for 2 days. The mRNA levels were determined by RT-PCR as described in ‛Material and Methods’.
Fig. 7. Effects of DCRT on the expression of LXRα and FAS in mRNA level
The 2-day post-confluent 3T3-L1 preadipocytes (day 0) were differentiated into adipocytes in the absence or presence of indicated concentrations of DCRT for 5 days. The mRNA levels were determined by RT-PCR as described in ‛Material and Methods’.
Fig. 8. Effects of DCRT on the expression of aP2, adipsin, LPL, CD36, TGF-β, and adiponectin in mRNA level
The 2-day post-confluent 3T3-L1 preadipocytes (day 0) were differentiated into adipocytes in the absence or presence of indicated concentrations of DCRT for 5 days. The mRNA levels were determined by RT-PCR as described in ‛Material and Methods’.
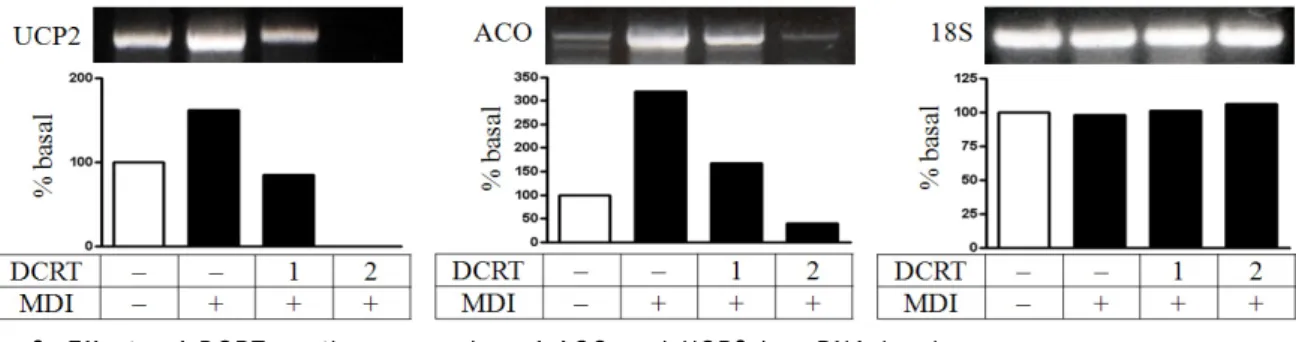
Fig. 9. Effects of DCRT on the expression of ACO and UCP2 in mRNA level
The 2-day post-confluent 3T3-L1 preadipocytes (day 0) were differentiated into adipocytes in the absence or presence of indicated concentrations of DCRT for 5 days. The mRNA levels were determined by RT-PCR as described in ‛Material and Methods’.
Fig. 10. Effects of DCRT on the expression of iNOS and IL-6 in mRNA level
The 2-day post-confluent 3T3-L1 preadipocytes (day 0) were differentiated into adipocytes in the absence or presence of indicated concentrations of DCRT for 5 days. The mRNA levels were determined by RT-PCR as described in ‛Material and Methods’.
5 일째 control group은 normal group에 비하여 발현이 증가하였고, DCRT group은 control group에 비해 2 ㎎/
㎖농도에서 발현 감소를 보였다 (Fig. 7).
FAS의 발현 양상을 살펴보면 FAS mRNA는 분화 5 일째 control group은 normal group에 비하여 발현이 증 가하였고, DCRT group은 control group에 비해 농도에 의존적으로 발현 감소를 보였다 (Fig. 7).
3) 지방세포 특이적 유전자 발현에 미치는 영향 aP2, adipsin, LPL, CD36, TGF-β, 그리고 adiponectin 의 발현 양상을 살펴보면 aP2, adipsin, LPL, CD36, TGF-β, 그리고 adiponectin mRNA의 경우 분화 5 일째 control group은 normal group에 비하여 발현이 증가하 였고, DCRT group은 control group에 비해 농도에 의존 적으로 발현 감소를 보였다 (Fig. 8).
4) 지방산화 관련 유전자의 발현에 미치는 영향 ACO와 UCP2 mRNA의 경우 분화 5 일째 normal group에 비해 control group에서 발현 증가를 보였으며, DCRT group에서 control group에 비해 발현 감소를 보
였다 (Fig. 9).
5) 염증 관련 유전자의 발현에 미치는 영향 3T3-L1 지방세포에서 발현이 증가하는 염증관련 유 전자인 iNOS와 IL-6에 미치는 영향을 알아보기 위하여 전구지방세포를 분화시킨 후 5 일째에 iNOS 및 IL-6 발현 정도를 측정하였다. iNOS와 IL-6 mRNA의 경우 분화 5 일째 normal group에 비해 control group에서 발 현 증가를 보였으며, DCRT group에서 control group에 비해 발현 감소를 보였다(Fig. 10).
3. 지방세포 분화 관련 신호전달 및 전사인자 조절 효과 1) ERK1/2 인산화 및 발현에 미치는 영향
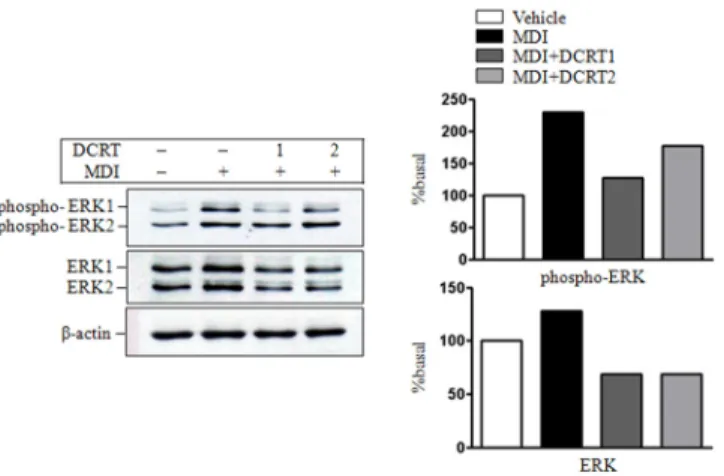
ERK1/2의 활성화에 미치는 영향을 알아보기 위하여 전구지방세포의 분화자극 10 분 후 ERK1/2의 단백질 발현과 인산화를 관찰하였다. ERK1/2 단백질 발현과 인산화 정도는 control group은 normal group에 비하여 발현 증가를 보였고 DCRT group은 control group에 비 해 발현 감소를 보였다(Fig. 11).
2) AKT의 인산화 및 발현에 미치는 영향
AKT의 활성화에 미치는 영향을 알아보기 위하여
Fig. 11. Effects of DCRT on the expression of phosphorylated ERK1/2 and ERK1/2 in protein level
The 2-day post-confluent 3T3-L1 preadipocytes (day 0) were differentiated into adipocytes in the absence or presence of indicated concentrations of DCRT for 10 mins. The protein levels of phosphorylated ERK and ERK were determined by western blotting as described in ‛Material and Methods’.
Fig. 12. Effects of DCRT on the expression of phosphorylated AKT and AKT in protein level
The 2-day post-confluent 3T3-L1 preadipocytes (day 0) were differentiated into adipocytes in the absence or presence of indicated concentrations of DCRT for 36 hrs. The protein levels of phosphorylated AKT and AKT were determined by western blotting as described in ‛Material and Methods’.
Fig. 13. Effects of DCRT on the expression of C/EBPα in protein level
The 2-day post-confluent 3T3-L1 preadipocytes (day 0) were differentiated into adipocytes in the absence or presence of indicated concentrations of DCRT for 5 days. The protein levels of C/EBPα were determined by western blotting as described in ‛Material and Methods’.
전구지방세포의 분화자극 36 시간 후 AKT의 단백질 발현과 인산화를 관찰하였다. AKT 단백질 발현과 인 산화 정도는 control group은 normal group에 비하여 약 간의 발현 증가를 보였고, DCRT group은 control group 에 비해 약간의 감소를 보였으나 현저한 발현 감소를 보이지 않았다(Fig. 12).
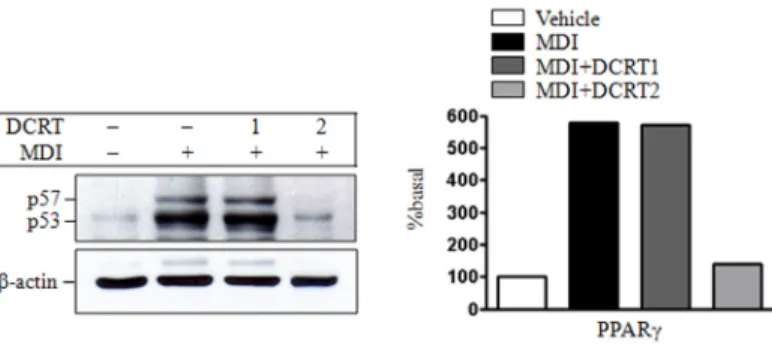
3) C/EBPα와 PPARγ의 발현에 미치는 영향 지방세포 분화관련 주요 조절 전사인자인 C/EBPα와 PPARγ의 활성화에 미치는 영향을 알아보기 위하여 전 구지방세포의 분화자극 5 일 후 C/EBPα와 PPARγ 의 단백질 발현을 관찰하였다. C/EBPα 단백질 발현은 con-
Fig. 14. Effects of DCRT on the expression of and PPARγin protein level
The 2-day post-confluent 3T3-L1 preadipocytes (day 0) were differentiated into adipocytes in the absence or presence of indicated concentrations of DCRT for 5 days. The protein levels of PPARγ were determined by western blotting as described in ‛Material and Methods’.
trol group은 normal group에 비하여 발현 증가를 보였 고, DCRT group은 control group에 비해 농도에 의존적 으로 발현 감소를 보였다(Fig. 13). PPARγ 단백질 발현 은 control group은 normal group에 비하여 발현 증가를 보였고 DCRT group은 control group에 비해 2 ㎎/㎖ 농 도에서 현저한 발현 감소를 보였다(Fig. 14).
Ⅳ. 考 察
비만은 섭취 에너지량이 소비 에너지량보다 많아서 나머지 에너지가 체지방으로 조직에 침착되어 대사장애 를 수반하는 질환이다. 한의학에서는 肥人, 肉人, 肥貴人 등으로 표현하였으며, 膏粱厚味의 과다섭취, 久坐나 久 臥 등의 운동부족, 內傷七情 등으로 인한 氣滯, 痰濁, 水濕, 瘀血 등이 연관되나, 근본적으로는 臟腑의 氣運이 虛하여 運化無力해져 濕痰이 발생하고, 동시에 水液代 射失調가 되어 비만의 원인이 된다고 하였다7, 8).
大靑龍湯은《傷寒論》3)에 “太陽中風 脈浮緊 發熱 惡寒 身疼痛 不汗出而煩躁者 大靑龍湯主之”, “傷寒 脈浮緩 身不疼 但重 乍有輕時 無少陰證者 大靑龍湯 發之”라고 기재되어 있다. 임상적 응용은 流行性感冒, 肺炎, 氣管支哮喘, 流行性髓膜炎, 痲疹, 胸膜炎, 急性 關節炎, 丹毒, 急性皮膚病性浮腫 등의 急性熱病 初期 의 高熱에 表寒과 內熱, 煩躁가 보일 때 응용할 수 있 다4).
이에 저자는 임상에서 호흡기 질환인 喘證과 鼻炎 등의 환자에게 大靑龍湯을 투여하던 중 체중감량의 효 과가 나타나, 비만환자 중 호흡기 질환을 겸한 환자에 게 응용해 보고 있던 중 大靑龍湯이 비만에 어떤 효과 가 있는지 실험해 보고자 하였다.
裵9)는 “傷寒 脈浮緩 身不疼 但重 乍有輕時 無少陰 證者 大靑龍湯發之” 병증을 體表에 外邪가 있고 體內 에 裏熱이 있으며 表邪의 鬱閉로 인하여 水濕의 邪氣 가 체표에 鬱閉된 것으로 보았다. 浮脈은 主表하여 病 邪와 病位가 體表에 있음을 나타내고, 緩脈은 主濕과 主水氣를 나타내므로, 본조의 기본 病機가 “浮緩의 相 合” 즉 水濕의 邪氣가 體表에 閉鬱한 病症이라고 하였 다. 그리고 三焦는 水道이고, 膀胱은 水府로 정상적인 상태에서는 腠理, 皮毛가 膀胱과 三焦의 氣化로 敷布 되는 津氣의 濡養을 받게 되는데, 表氣가 鬱閉하고 腠 理가 不開하여 玄府가 不通하게 되면 體表로 布敷되 는 津氣가 凝聚하여 水濕의 邪氣가 되고 肌腠와 皮膚 에 鬱閉하게 되므로 “身疼, 但重”의 병증이 나타나므 로, 水濕의 邪氣로 인해 沈重하게 느껴지는 것을 의미 한다고 하였다. 이 경우 “其在皮者 汗而發之”의 원칙 에 따라 發散, 發越, 發汗의 치법을 사용할 수 있다고 하였다9). 이에 한의학적으로 濕痰의 발생과 동시에 水 液代射失調가 비만으로 진행되는 하나의 病機가 되므 로, 大靑龍湯이 體表로 鬱閉된 水濕의 邪氣를 發表하 고 內熱을 식혀 식욕을 조절하는 작용으로 비만에 응 용될 수 있으리라 생각되어, 3T3-L1 전구지방세포를 이용하여 지방세포의 증식과 분화에 미치는 영향에 대 해 알아보고자 실험하였다.
비만은 열량의 과다 섭취와 소비의 불균형으로 발생 되는 대사성 질환으로, 지방조직의 증가 상태로 정의 되며 만성적인 염증 반응을 동반한다10). 최근에는 지방 조직이 지방을 저장하는 단순한 에너지 저장고의 역할 이외에 신체 생리 대사적인 변화를 일으키는 다양한 adipokine이라 불리는 물질들을 분비하는 내분비 역할 을 담당하여 체내의 에너지 항상성을 조절하는데 중요 한 역할을 하는 것으로 보고되고 있다. 비만의 경우 지
Scheme 2. Transcription factor activation, regulatory pathway, and adipocyte specific expressed genes during adipogenesis
방세포에서 염증반응의 표지가 되고 있는 물질들을 생 산하고 분비하는 것으로 알려져 있다11). 따라서 지방세 포의 증식과 지방세포에서 분비되는 물질들에 대한 이 해와 그 생체내 분화 조절 기전에 대한 이해가 비만 및 그로 인한 여러 질병들을 효과적으로 치료할 수 있 을 것으로 사료된다.
본 실험에서 사용한 3T3-L1 세포주는 분화된 3T3-L1 adipocytes가 in vivo adipocytes의 생화학적 형태학적 성 질들과 사실상 동일하다는 보고12)에 따라 지방 대사를 연구하는데 적절하다고 판단되어 3T3-L1 전구지방세 포를 분화시켜 사용하였다. 그리고 실험은 지방세포 분화억제, 지방세포 분화 관련 전사인자와 유전자 발 현조절 및 지방세포 분화 관련 신호전달경로의 활성 조절에 관한 실험으로 크게 세 방법으로 나누어 진행 하였다 (Scheme 2).
지방세포 분화 억제 효과에 관한 실험 결과는 다음 과 같다.
3T3-L1 전구지방세포의 증식반응에서 DCRT 농도 와 처리시간에 따라 세포의 증식이 저해되지 않아 2
㎎/㎖까지의 DCRT가 3T3-L1 전구지방세포에 독성을 나타내지 않음을 알 수 있었다 (Fig. 1).
전구지방세포의 분화과정에서 형태학적 변화를 관 찰한 결과 전구지방세포의 세포밀도가 100% confluent 된 세포형태는 길게 뻗은 섬유아세포의 형태를 보여주 었으나(Fig. 2A), 분화 유도 후 1 일, 2 일과 5 일째 세포 의 형태는 시간이 지남에 따라 세포 모양이 둥글어지 고 세포질 내 지방구들이 점차 많이 형성되었고 (Fig.
2B). 반면 DCRT group은 농도에 비례하여 세포내 지 방구의 생성이 억제되는 것을 관찰할 수 있었다 (Fig.
2C, D).
분화된 지방세포 내 중성지방 생성에 대한 영향을
관찰하기 위해 Oil red O를 이용하여 염색을 시행하여 지방세포의 형태를 관찰하고 세포내 중성지방의 양을 관찰한 결과 control group에 비해 1 ㎎/㎖과 2 ㎎/㎖의 DCRT group에서 지방세포의 형성이 억제되어 있는 것으로 관찰되었다 (Fig. 3).
이러한 결과를 통해 DCRT이 지방세포 분화를 억제 하는 효과가 있음을 알 수 있었다.
지방세포 분화 관련 전사인자와 유전자 발현조절에 대한 실험결과는 다음과 같다. 전구지방세포에서 형태 학적, 생화학적으로 완전히 성숙된 지방세포로 분화되 는 과정은 호르몬, cytokine 그리고 전사인자 등의 여러 인자들의 상호조절을 통해 이루어지는데13), SREBP1은 전구지방세포 분화초기에 매우 빠르게 유도되어 PPAR γ와 함께 지방세포 분화를 촉진시키는 역할을 하며 지 방대사에 관련된 많은 유전자를의 발현을 증가시킴으 로써 지방대사를 촉진한다14, 15).
전구지방세포가 adipogenetic signal을 받으면 전사인 자 C/EBPβ와 C/EBPδ의 발현이 유도되고 C/EBPβ와 C/EBPδ 발현의 일시적인 증가는 PPARγ 발현 증가에 선행해서 일어나며, 그 후 C/EBPβ와 C/EBPδ 발현의 감소는 C/EBPα의 유도를 수반하며 분화를 진행한다16).
C/EBPα는 여러 지방조직 특이성 유전자들이 발현되 기 전에 증가하여 에너지 항상성을 조절하며17), PPARγ 는 adipogenesis의 주요 조절자로서 leptin, fatty acid syn- thase(FAS) 및 fatty acid binding protein(aP2)등의 adipo- genic gene들의 발현을 조절한다. PPAR는 핵내 수용체 이고, 스테로이드 호르몬 수용체나 활성형 지용성 비 타민 수용체와 유사한 구조를 가지며 α, β, γ 등의 여 러 아형이 알려져 있는데, 이 중 PPARγ는 지방분화에 관여하여 지방대사 관여 유전자들의 발현을 조절하는 것으로 알려져 있다18).
지방세포 분화 초기에 조절 작용하는 전사인자의 발 현 양상을 관찰한 결과 DCRT는 분화 1 일째 SREBP1 mRNA, 분화 2 일째 C/EBPβ와 C/EBPδ mRNA와 C/EBPα와 PPARγ mRNA의 발현 증가를 감소시켰다 (Fig. 4, 5, 6).
이러한 전사인자들의 발현양상은 대청룡탕이 지방 세포의 분화과정에서 지질 생성에 미치는 영향과 일치 하여 분화 관련 전사인자들의 발현을 조절하여 지방합 성을 조절하고 있음을 알 수 있었다.
지방합성에 관련된 유전자의 발현에 대한 영향을 살 펴보기 위해 지방합성에 핵심역할을 하는 핵수용체로 지방생성과 축적에 관여하는 LXRα과 지방산 합성효소 인 FAS의 발현양상을 관찰한 결과 DCRT는 분화 5 일 째 LXRα mRNA와 FAS mRNA의 발현증가를 감소시켰 다 (Fig. 7).
전구지방세포가 증식과 분화과정을 거쳐 지방세포 로 되는 과정에서 지질이 축적되고 지방세포 특이적인 유전자가 발현되는데 이에 대한 영향을 살펴보기 위해 aP2, adipsin, LPL, CD36, TGF-β, 그리고 adiponectin의 발현 양상을 관찰한 결과 DCRT는 분화 5 일째 aP2, adipsin, LPL, CD36, TGF-β, 그리고 adiponectin mRNA 의 발현을 감소시켰다 (Fig. 8).
3T3-L1 지방세포에서 발현되어 지방산화를 조절한 다고 알려진 유전자의 발현에 미치는 영향을 알아보 기 위하여 분화 5 일째에 지방산화 효소인 ACO와 에 너지 소비 단백질인 UCP2의 발현 정도를 관찰한 결과 DCRT는 분화 5 일째 ACO와 UCP2 mRNA의 발현을 감소시켰다 (Fig. 9). 이는 대청룡탕이 지방산화를 통해 에너지 소비를 증가시키는 효과는 없는 것으로 사료 된다.
3T3-L1 지방세포에서 발현이 증가하는 염증관련 유 전자에 미치는 영향을 알아보기 위하여 분화 5 일째에 iNOS 및 IL-6 발현 정도를 관찰한 결과 DCRT는 iNOS 및 IL-6 mRNA의 발현을 감소시켰다 (Fig. 10).
지방세포 분화관련 신호전달 및 전사인자 조절에 대 한 실험결과는 다음과 같다. 지방세포 분화과정에서 mitotic clonal expansion이라 불리는 증식단계는 인슐린 과 같은 분화자극에 의해 시작되는데 이 과정에서 ERK pathway의 활성화가 일어난다고 알려져 있다. 게 다가 PPARγ발현은 mitotic clonal expansion 단계에서 일어나지 않으며 ERK 활성이 약화되고 마지막 분화가 진행되는 동안 점차 증가한다. 따라서 ERK 활성은 지 방세포 분화의 조절자인 C/EBP α, β, δ와 PPARγ의 발
현에 중요한 역할을 한다19). ERK1/2의 활성화에 미치 는 영향을 알아보기 위하여 전구지방세포의 분화자극 10 분 후 ERK1/2의 단백질 발현과 인산화를 관찰한 결 과 DCRT는 ERK1/2 인산화를 억제하였다 (Fig. 11).
AKT 활성화는 insulin signaling pathway에 관련되어 지방세포 분화과정을 조절하며20), AKT의 지속적인 활 성은 지방세포의 자발적인 분화를 일으키고21), PPATγ 의 활성을 직접적으로 조절하여 지방분화를 촉진한다 고22) 알려져 있다. AKT의 활성화에 미치는 영향을 알 아보기 위하여 전구지방세포의 분화자극 36 시간 후 AKT의 발현과 인산화를 관찰한 결과 DCRT는 약간의 발현 감소를 보였으나 현저한 발현 감소를 보이지 않 았다 (Fig. 12).
또한 지방세포 분화관련 주요 조절 전사인자인 C/EBPα와 PPARγ의 활성화에 미치는 영향을 알아보기 위하여 분화자극 5 일 후 C/EBPα와 PPARγ 단백질 발 현을 관찰한 결과 DCRT는 C/EBPα와 PPARγ 단백질 발현을 감소시켰다 (Fig. 13, 14).
본 연구결과를 종합해 볼 때 대청룡탕은 지방세포분 화를 유의하게 억제하는 것으로 나타났는데, 이러한 효과는 SREBP1, C/EBPβ와 C/EBPδ, 그리고 C/EBPα, PPARγ 지방분화 전사인자와 ERK1/2와 AKT 등의 신 호전달의 조절을 통해 전구지방세포의 지방세포로의 분화를 초기에 억제하여 LXRα와 FAS 지방합성효소와 aP2, adipsin, LPL, CD36, TGF-β와 adiponectin 같은 지 방세포 특이적 유전자의 발현과 iNOS와 IL-6 염증관련 유전자를 억제하는 것으로 사료된다.
Ⅴ. 結 論
대청룡탕이 3T3-L1 세포에서 지방세포 분화기전 에 미치는 영향을 관찰한 결과 다음과 같은 결론을 얻었다.
1. 대청룡탕은 농도에 따라 지방분화와 중성지방의 축 적을 유의하게 억제하였다.
2. 대청룡탕은 지방분화 전사인자인 SREBP1, C/EBPβ 와 C/EBPδ, C/EBPα, PPARγ mRNA의 발현을 억제 하였다.
3. 대청룡탕은 지방합성에 관여하는 LXRα와 FAS mRNA와 지방특이 단백질인 aP2, adipsin, LPL, CD36, TGF-β와 adiponectin mRNA의 발현을 억제
하였다.
4. 대청룡탕은 지방세포의 분화산물 중 염증관련 iNOS 와 IL-6 mRNA의 발현을 감소시켰다.
5. 대청룡탕은 지방세포의 산화를 유도하는 ACO와 UCP2 mRNA의 발현을 증가시키지는 못했다.
6. 대청룡탕은 지방세포의 분화조절과 관련된 신호전 달경로인 ERK1/2와 AKT의 활성화를 억제하였다.
Ⅵ. 參 考 文 獻
1. 대한비만학회. 임상비만학. 서울: 고려의학. 2001:19-29.
2. Ahima RS, Flier JS. Adipose tissue as an endocrine organ.
Trends in Endocrinology & Metabolism. 2000;11(8):
327-32.
3. 張仲景. 傷寒論. 北京:人民衛生出版社. 1990.
4. 丹波元簡. 傷寒論輯義. 北京:人民衛生出版社. 1989.
5. 박완열. 대청룡탕 및 <석실비록> 급치법방이 호흡기 점액의 분비에 미치는 영향. 서울:동국대 석사학위논 문. 2006.
6. 정영재. 대청룡탕 등 수종의 처방이 기도배상세포에서 의 mucin분비에 미치는 영향에 관한 연구. 서울:동국대 박사학위논문. 2006.
7. 김동환, 이경섭. 비만관리센터 치료 성과에 대한 연구.
대한한방부인과학회지. 2001;14(1):319-27.
8. 신동길, 김덕곤, 이진용. 비만아 9례에 대한 곽향정기산 가미방의 치료효과에 대한 임상적 고찰. 대한한방소아 과학회지. 2001;15(1)183-94.
9. 裵永淸. 傷寒論臨床應用五十年. 北京:學菀出版社.
1995:66-8.
10. Spiegelman BM, Flier JS. Adipogenesis and obesity: round- ing out the big picture. Cell. 1996;87(3):377-89.
11. Ahima RS, Flier JS. Adipose tissue as an endocrine organ.
Trends in Endocrinology & Metabolism. 2000;11(8):
327-32.
12. MacDougald OA, Hwang C, Fan H, Lane MD. Regulated expression of the obese gene product (leptin) in white adipose tissue and 3T3-L1 adipocytes. Preceedings of the National Academy of Sciences. 1995;92(20):9034-7.
13. Gregoire FM, Smas CM, Sul HS. Understanding adipocyte differentiation. Physiological Reviews. 1998;78:783-809.
14. JB KIM, Spiegelman BM. ADD1/SREBP1 promotes
adipocyte differentiation and gene expression linked to fatty acid metabolism. Genes Dev. 1996;10:1096-107.
15. JB KIM, Spotts GD, Halvorsen YD, Shih HM, Ellenberger T, Towle HC, Spiegelman BM. Dual DNA binding specif- icity of ADD1/SREBP1 controlled by a single amino acid in the basic helix-loop-helix domain. Mol. Cell. Biol.
1995;15:2582-8.
16. Camp HS, Ren D, Leff T. Adipogenesis and fat-cell function in obesity and diabetes. Trends in Molecular Medicine. 2002;8(9):442-7.
17. Long SD, Pekala PH. Lipid mediators of insulin resistance:
ceramide signalling down-regulates GLUT4 gene tran- scription in 3T3-L1 adipocytes. Biochemical Journal. 1996;
319:179-84.
18. Hwang CS, Loftus TM, Mandrup S, Lane MD. Adipocyte differentiation and leptin expression. Annual review of cell and developmental biology. 1997;13:231-59.
19. Bost F, Aouadi M, Caron L, Binétruy B. The role of MAPKs in adipocyte differentiation and obesity. Biochimie.
2005;87(1):51-6.
20. Sakaue H, Ogawa W, Matsumoto M, Kuroda S, Takata M, Sugimoto T, Spiegelman BM, Kasuga M. Posttranscrip- tional control of adipocyte differentiation through activa- tion of phosphoinositide 3-kinase. J Biol Chem. 1998;273 (44):28945-52.
21. Kohn AD, Summers SA, Birnbaum MJ, Roth RA.
Expression of a constitutively active Akt Ser/Thr kinase in 3T3-L1 adipocytes stimulates glucose uptake and glu- cose transporter 4 translocation. J Biol Chem. 1996;271 (49):31372-8.
22. Dowell P, Otto TC, Adi S, Lane MD. Convergence of peroxisome proliferator-activated receptor gamma and Foxo1 signaling pathways. J Biol Chem. 2003;278(46):
45485-91.