21
종묘장 넙치 치어에서의 Iridovirus 감염
김태중·장은진·김종수1·이재일*
전남대학교 수의과대학
1전남생물산업연구센터 (게재승인: 2006년 1월 15일)
Iridovirus infection of cultured juvenile flounder (Paralichthys olivaceus) in nursery
Tae-Jung Kim, Eun-Jin Jang, Jong-Su Kim
1, Jae-Il Lee*
College of Veterinary Medicine, Chonnam National University, Gwangju 500-757, Korea
1
Jeonnam Biotechnology Research Center, Hwasun 519-805, Korea
(Accepted: January 15, 2006)
Abstract : Iridovirus is an icosahedral cytoplasmic double-stranded DNA virus with a genome size of 170- 200kb. Outbreaks of fish iridovirus infection are characterized by their wide geographic distribution and broad host spectrum, especially in water temperatures of 25-27
oC Recently, the causative agent of high mortalities in flounder (
Paralichthys olivaceus) was identified as fish iridovirus in Korea. Iridoviral infection repeatedly occurs in the same area for long periods, suggesting the possibility of viral infection in nursery. To examine this, the existence of iridovirus in juvenile flounders was detected by PCR using virus-specific primers.
Antibodies induced against this virus were also examined using ELISA. As a result, juvenile fish in nursery were found to be previously infected with iridovirus, suggesting that proper prevention systems are required.
Key words : ELISA, iridovirus, PCR, recombinant protein
서 론
Iridovirus
는icosahedral cytoplasmic DNA virus
로서genome
의 크기가170-200 kb
인double-stranded DNA virus
이다[25].
이바이러스의큰특징인약50 kDa
의major capsid protein(MCP)
은바이러스감염을일으키는 주된polypeptide
이고, virion protein
의약45%
를구성한 다[23]. Iridoviridae family
는4
개의 속,
즉Iridovirus , Chloririvirus , Ranavirus ,
그리고Lymphocystivirus
로나뉘 는데,
최근iridovirus
사이의유전학적다양성을진단하는도구로자주사용되는
MCP
유전자서열분석법[23]
을이용하여분석해본결과
, fish iridovirus
는Ranavirus
속또는
Lymphocystivirus
속과는유전학적으로많이다 르다는것이밝혀졌다[4, 6].
Fish iridovirus
는넓은지역분포와숙주범위를가지며[10, 11],
보통18-20
oC
에서25-27
oC
로수온상승이되는시기에발병한다
.
이질병에걸린고기는특별한외부증 상없이,
비정상적이고무기력한유영을보이고, 2
개월 동안50-90%
의누적치사율을보인다[24].
조직병리학적소견의경우아가미
,
신장,
심장,
간,
비장에서비대 호염기세포가보인다[24]. Iridovirus
는자생어에서수입 어뿐아니라,
자연어에서양식어까지[5, 17],
그리고치어에서성어까지
[13]
널리퍼져어류산업에있어큰경제적손실을주고있다
.
우리나라의경우
1998
년8-10
월에남해안지역의양식돌돔
, Oplegnathus fasciatus
의대량폐사의원인이Red sea bream iridovirus(RSIV)
와유사한iridovirus
로밝혀진 이후[21],
다양한종류의양식어류에서iridovirus
감염*Corresponding author: Jae-Il Lee
College of Veterinary Medicine, Chonnam National University, Gwangju 500-757, Korea [Tel: +82-62-530-2854, Fax: +82-62-530-2857, E-mail: [email protected]]
증이발생하고있다
[3, 7, 9].
넙치( Paralichthys olivaceus )
의경우
, 2003
년양식넙치치어의대량폐사역시RSIV
와유사한
iridovirus
로밝혀졌다[3].
이감염증은수온23-26
oC
사이에서발생하였으며1
년생뿐아니라2-3
년생어류도폐사하여막대한손실을입혔다
.
한편
iridovirus
감염증은이전발생지역에서지속적으로발병하는특징을보이는바
,
발병이입식전종묘장에서부터
iridovirus
에감염되었는지아니면양식장에서감염되었는지알아볼필요가있겠다고할수있다
. Iridovirus
감염을알아보기위한진단법으로cell culture [14], immunofluorescence assay [13], PCR [15-17]
등이연 구되었는데,
본실험에서는정확한감염의여부를알아보기위하여
RSIV MCP
에대한PCR
방법에의해직접MCP
유전자를 검출함과 함께, MCP
를대장균 발현system
을이용하여재조합단백질로발현하고이단백질에대한항체의존재여부를
ELISA
방법으로알아봄으로써간접적인진단을병행하여실시하여보았다
.
이 를통하여종묘장내넙치치어에서의iridovirus
감염상 황을조사함으로서, iridovirus
감염증에대한방역대책수립과백신적용의기초자료로사용하고자한다
.
재료 및 방법
시험어 및 DNA 준비
2005
년8
월 전북K
지역의 종묘장으로부터 넙치( Paralichthys olivaceus )
치어(2
개월령,
체장: 7-10 cm)
를20
개체를채집하여시험에사용하였다.
각개체별로비장과 간을적출한다음조직
1 g
당lysis buffer(50 mM Tris-HCl, pH 8; 20 mM NaCl, 2% SDS, 10 mM EDTA, pH 8; proteinase K 100
µg/m) [24]
를1 ml
씩 첨가하고미세수술용가위로조직을세절한뒤
56
oC
에서3
시간동 안용해시켰다. DNA
추출은Sambrook
등[19]
에준하 여phenol/chloroform
방법을실시하였다. Genomic DNA
는
TE buffer(10mM Tris-HCl; 1mM EDTA, pH 7.5)
에녹여
50 ng/
µl
의농도로맞춘후, PCR template
로사용하 였다.
PCR 반응(polymerase chain reaction)
Primer
는GenBank
에 등록되어 있는RSIV MCP (AY310918) [23]
를이용해서Bam HI site(underlined)
를포 함하는5’-primer(5'-TATAGGATCCATGTCTGCGA TCT CAGGTCG-3')
와Hind III site(underlined)
를포함하는3’- primer(5'-TGTGAAGCTTTTACAGGATAGGGAAGC CTG-3')
를제작하여사용하였다. PCR
조건은2
µl DNA
와
10X reaction buffer 5
µl
의 각primer(50 pM) 1
µl, 10 mM dNTPs(
각2.5 mM) 5
µl, 5U Ex Taq
TMDNA
polymerase(Takara, Japan) 1
µl
을혼합한 후,
총50
µl
이되도록
DW
를첨가시켰다.
이를PTC-100(MJ Research, USA)
을이용해95
oC 5
분간incubation
시킨후95
oC/40s denaturation, 55
oC/40s annealing, 72
oC/2 min extension
을30
회반복수행한후72
oC
에서5
분간다시extension
을실시하였다
.
증폭된반응생성물은1.0%(w/v) agarose gel
에서전기영동한후
DNA
크기를비교하였다. PCR
로증폭된생성물을
1.0% agarose gel
에서gel-cut
후PCR purification kit(Qiagen, Germany)
를이용하여순수분리 하고이를pGEM-T easy vector(Promega, USA)
에삽입하여
cloning
하였다.
이construct
를E. coli JM109
에형 질전환시켜배양시킨후, QIAprep Spin Mini Kit (Qiagen, Germany)
을이용하여DNA plasmid
를추출하였다.
준비된
construct plasmid
내에원하는크기의insert
가삽입되었는지 알아보기 위하여 제한효소
Eco RI (Promega,
USA)
를이용하여restriction study
를하였다.
그리고이construct
를ABI Prism 3100 Genetic Analyzer (Applied
Biosystems, USA)
를이용하여염기서열을분석하였다.
분석된염기서열을
GenBank
에등록된RSIV
유전자들과염기서열을비교분석하였다
.
재조합 단백질 발현용 construct
재조합단백질의생산 및순수분리는김등
[1]
및Kim and Lee [8]
에준하여실시하였다.
즉, pGEM-T easy
에
cloning
된capsid
유전자를2
가지제한효소( Bam HI + Hind III)
로처리한다음양쪽끝을sticky ends
로만들 었다.
여기에단백질에hexahistidine-tag
를fusion
시키는기능이있는대장균발현용
vector
인pRSET(Invitrogen, USA) [22]
를같은효소로( Bam HI + Hind III)
처리하여T4 DNA ligase(Promega, USA)
를이용하여insert
와vector
를
ligation
하였다.
이construct
를JM109
에서 증균후insert
존재및open reading frame
의shift
여부를sequencing
을통해재확인하였다
.
확인된construct
는단백질발현용대장균
BL21(DE3)pLysS
에형질전환시켜IPTG
를첨가하여단백질을발현하고
hexahistidine
에결합하는Ni
2+- charged resin
을 이용하는Probond(Invitrogen, USA) purification kit
를이용하여denaturing condition
에서단백 질을순수분리 하였다.
순수분리된 단백질은ELISA study
에서iridovirus capsid protein
에대한항체의존재여부를알아보기위한항원으로사용되었다
.
ELISA
ELISA
는Seferian
과Martinez [20]
에준하여실시하였 다.
즉96-well plates(Maxisorp, Nunc, Denmark)
에재조 합capsid
단백질을(1
µl/well/90
µl) coating
하고1% skim
milk
가 들어있는PBS
로blocking
한 다음PBST
(PBS + 0.05% Tween-20, pH 7.4)
로3
회washing
하였다.
치어로부터혈액을채취하여총
20
개의혈청시료(1
차항체
)
를준비하고이를1 : 100
으로희석하여사용하였다
. 2
차항체로는mouse anti-flounder immunoglobulin monoclonal antibody(
경상대학교수의과대학정태성교수
)
를1 : 100
으로희석하여사용하였고3
차항체로는goat anti-mouse IgG horseradish peroxidase-conjugated(Pierce, USA)
를1 : 500
으로희석하여사용하였다.
각반응은모 두90
µl
의항체액을사용하였고반응은모두37
oC
에서1
시간동안 반응하였고PBST
로3
회washing
하였다.
Negative control
로는각단계의항체를하나씩제거한시료및기질용액만을첨가한
well
의값을이용하여이 들의평균값을이용하였다. 3
차항체의존재를알아보기위한발색반응으로는
ABST
기질용액[0.1 M citric acid buffer, pH 4.0 10 ml; ABTS stock solution(ABTS 100 mg in 4.5 ml DW) 250
µl; H
2O
250
µl]
을사용하였고반응15
분후흡광도405 nm
에서ELISA reader(Multiskan EX, Thermo LabSystems, USA)
를이용하여발색정도를측정 하였다.
결과는 평균±
표준편차(SD)
로 나타내었고Student’s t -test
를이용하여검증하였으며, p < 0.01
을통 계적으로유의하다고판단하였다.
결 과
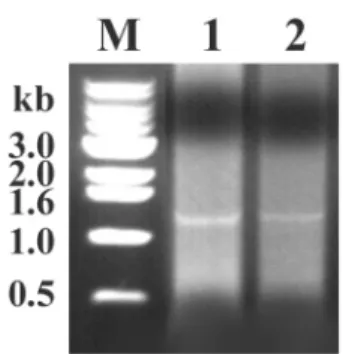
PCR 반응
각넙치의비장및간에서분리된
DNA
를template
로 이용하여RSIV MCP-specific primer
를이용하여PCR
을실시한결과
,
모든시료에서예상되는크기의(1362 bp)
반응생성물이증폭되었고
20
개의시료중1
번시료의 결과를그림1
에나타내었다.
이PCR
생성물이조사대상인
iridovirus
의capsid
유전자인지를알아보기위한sequencing
을위하여cloning vector
에cloning
하였다.
염기서열분석(sequencing)
넙치의비장으로부터
(Fig. 1, Lane 1) PCR
에의해증 폭된1362 bp
의 반응생성물을pGEM-T easy cloning vector
에cloning
하였다.
이후plasmid DNA
를miniprep
하여
construct
내에원하는insert
가삽입되었는지를알아 보기위하여제한효소Eco RI
를이용하여insert
삽입여부를알아보았다
(Fig. 2).
그리고이plasmid
를이용하여sequencing
을실시하였다.
이결과를GenBank
에등록된RSIV
의capsid
유전자서열과homology
를비교한결과,
넙치에서 얻어진
PCR
생성물은RSIV
의MCP cDNA
gene
과99.49%
상동성을보였다(1355 bp/1362 bp).
이로 써입식전종묘장내의치어에이미iridovirus
가감염이되어있음을알수있었다
.
ELISA
Iridovirus
의capsid protein
을재조합단백질로 발현한 결과,
형질전환이전의대장균시료인control(Fig. 3, lane 1)
에서는나타나지않는50 kDa
크기의단백질(Fig. 3, Lane 2)
이발현되었으며,
순수분리결과,
다른단백질의 오염이없음을확인하였다(Fig. 3, Lane 3).
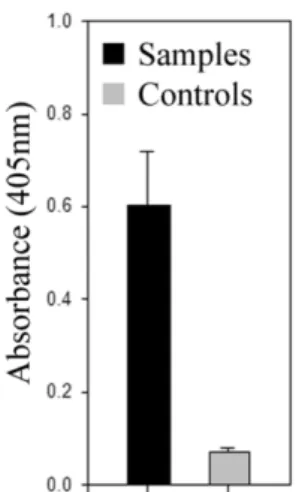
이를항원으로사용하여
iridovirus
의감염여부를간접적으로알아본
ELISA
실험 결과,
실험군전체의시료들은(Fig. 4, black;
평균OD = 0.6, SD = 0.11) negative control(Fig. 4, grey;
평균OD = 0.07, SD = 0.008)
에비해,
통계적으로 높은수치를나타내었다( p < 0.01).
이로써종묘장치어 에서의iridovirus
감염을재확인할수있었다.
고 찰
Iridovirus
는한국내여러지역과다양한어종에전파Fig. 1. The major capsid protein (MCP) gene of fish iridovirus was amplified using MCP-specific primers. Genomic DNA from spleen (lane 1) and liver (lane 2) of juvenile olive founder was used as a template. M: 1 kb size marker.
Fig. 2. Iridovirus capsid gene (1.3 kb) was cloned into the
pGEM-T easy (3 kb) cloning vector as described in the
Materials and methods. Clone containing iridovirus capsid
gene was digested with a restriction enzyme (
EcoRI),
generating two (3 kb+1.3 kb) restriction fragments. M: 1 kb
size marker.
되어있다
[2]. Iridovirus
감염증은특별한외부증상없이치사율이높고
[12, 24],
여러종류의어류및다양한연령에서
[5, 13, 17, 18]
발생하기때문에한국내양식어류산업에큰피해를주고있다
. Iridovirus
감염발생은수온과상당히관련되었기에
[24]
우리나라에서는5-8
월내 에많이발생한다.
이질병에대한방역과백신접종계획을세우기위해
서는어류성장단계중어느시기에주로
virus
에노출되어감염되는지알아야하기에본실험을실시해본결 과
,
전북K
지역종묘장으로부터유래되는치어에는이미
iridovirus
가감염이되어있는것으로나타났다.
넙치와돌돔을주로양식하는서남해안일대에서의
iridovirus
의감염증에의한대량폐사의원인으로수온이높은서 남해안일대바닷물의오염을들수있고
,
오염된바닷 물재순환으로인한종묘장 치어들의감염,
또한입식 후바닷물을통한이웃양식장으로의iridovirus
의전파를들수있겠다
.
이러한iridovirus
의확산에대해현재 까지뚜렷한방역대책이없는것이사실이며,
이러한이류로
iridovirus
감염에대한대책은거의전무하다고할수있다
.
하지만종묘장내넙치치어에서철저한바닷물 소독등으로iridovirus
의감염을막고,
감염여부를본 실험에서확립된방법들을이용하여확인하며,
양식장으로의입식전에양식장의청소와철저한소독을한다 면
virus
증식을막을수있을것으로예상된다.
또한,
서 남해안일대의바닷물이이미오염되었기에,
입수전치어상태에서의백신접종을 통해서만백신효과를 기대할 수있겠다
.
본실험에서는한정된지역
,
한정된어종과수를가지고했기때문에
, iridovirus
의전체방역과백신에적용 할수는없겠지만,
이를기초자료로활용하여iridovirus
감염과전파를막을수있는보다체계적인대책을수 립하는데기여하고자한다
.
감사의 글
본실험을위해단클론항체를제공해주신경상대학 교수의과대학정태성교수님께감사의말씀을전합니다