Correspondence to: Chang-Ju Kim, Department of Physiology, College of Medicine, Kyung Hee University, 1, Hoigi-dong, Dongdae- moon-gu, Seoul 130-701, Korea
Tel: +82-2-961-0407, E-mail: [email protected] Received November 15, 2009, Revised November 20, 2009, Accepted December 5, 2009
Effects of Schizandrae fructus on Lipopolysaccharide-Induced Cyclooxygenase-2 and Inducible Nitric Oxide Synthase Expressions in
Mouse BV2 Microglial Cells
*Department of Physiology, College of Medicine, Kyung Hee University,
†Department of Physical Education, Korea Military Academy, Seoul, Korea
Yun-Hee Sung*, Mal-Soon Shin*, Sang-Won Lee†, Chang-Ju Kim*
Schizandrae fructus is a medicinal fruit that has long been used for the treatment of dyspnea, cough, mouth dryness, amnesia, and nocturnal emission. Several studies have reported the anti-asthmatic, anti-hepatotoxic, anti-oxidative, and anti-diabetic effects of Schizandrae fructus; however little is known about the mechanisms by which the anti-inflammatory of this herb. In this study, we evaluated the effects of aqueous extract of Schizandrae fructus on lipopolysaccharide (LPS)-induced inflammation in mouse BV2 microglial cells. To accomplish this, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay, reverse trans- cription-polymerase chain reaction (RT-PCR), Western blot, prostaglandin E2 (PGE2) immunoassay, and nitric oxide (NO) detection were conducted. The results revealed that the aqueous extract of Schizandrae fructus suppressed PGE2 synthesis and NO production by inhibiting the LPS-stimulated enhancement of cyclooxygenase-2 (COX-2) and inducible nitric oxide synthase (iNOS) expression in mouse BV2 microglial cells. This extract also suppressed the expression of phosphorylated forms of mitogen-activated protein kinases (MAPKs). These results demonstrate that Schizandrae fructus may exert anti-inflammatory, and that these effects are associated with the suppression of COX-2 and iNOS expressions via down-regulation of MAPKs. (Korean J Str Res 2009;17:415
∼424)
Key Words: Schizandrae fructus, Mitogen-activated protein kinase, Nitric oxide, Prostaglandin E2
INTRODUCTION
Inflammation is a complex process that commences with a primary reaction in tissues. Once inflammation occurs in response to infection or injury in the central nervous system (CNS),
microglia, a resident macrophage-like population of brain cells, become activated. Activated microglia release various inflamma- tory mediators including tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), nitric oxide (NO), and prostanoids (Liu et al., 2003; Moon DO et al., 2007). Prostanoids, which regulate a number of physiopathological processes, are synthesized by two cyclooxygenase (COX) isoforms: COX-1 and COX-2. COX-1 is the constitutively expressed form during normal physiologic func- tions, whereas COX-2 increases prostaglandins (PGs) in response to inflammatory stimuli (Mitchell et al., 1995; Crofford et al., 2000).
NO is a neuromodulator in the CNS that acts as an important physiological messenger in many biological systems, including immunological, neuronal, and cardiovascular tissues (Bredt et al., 1994). NO is produced by three types of nitric oxide synthase (NOS): endothelial NOS (eNOS), neuronal NOS (nNOS), and inducible NOS (iNOS). Of these, iNOS catalyzes the oxidative deamination of L-arginine to produce NO, which is a potent pro-inflammatory mediator (Surh et al., 2001). These inflam- matory mediators can be activated by phosphorylated kinases, including mitogen-activated protein kinases (MAPKs) such as extracellular signal-regulated kinase (ERK), p38, and c-Jun NH2-terminal kinase (JNK). Many studies have reported that a key mediator in the inflammation response, lipopolysaccharide (LPS), activates these MAPKs in microglia, macrophages and other types of cells (Guha et al., 2001; Kim JH et al., 2006; Ha SK et al., 2008).
Schizandrae fructus is the fruit of Schisandra chinensis Baill, which has been classified into the family Magnoliaceae. This medicinal herb has traditionally been used to treat diseases such as gastrointestinal infection, diabetes mellitus, and hyperlipidemia in the Far East (Park JS et al., 2004). Additionally, several studies reported that Schizandrae fructus has anti-asthmatic, anti-hepa- totoxic, anti-oxidative, and anti-diabetic effects (Xu et al., 2008).
However, the mechanisms by which the anti-inflammatory of Schizandrae fructus occur have not been fully elucidated.
In the present study, we investigated the effects of aqueous extract of Schizandrae fructus on LPS-induced inflammation in mouse BV2 microglial cells. To accomplish this, 3-(4,5-dime- thylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay, reverse transcription-polymerase chain reaction (RT-PCR), Wes- tern blot, PGE2 immunoassay, and NO detection were con- ducted.
MATERIALS AND METHODS
1. Cell culture
Mouse BV2 microglial cells were cultured in Dulbecco's modified Eagle’s medium (DMEM; Gibco BRL, Grand Island, NY, USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Gibco BRL) at 37oC in 5% CO2-95% O2 in a humidified cell incubator. The cells were then plated onto culture
dishes at a density of 2×104 cells/cm2 at 24 h prior to drug treatment.
2. Preparation of the aqueous extract of Schi- zandrae fructus
To obtain the aqueous extract of Schizandrae fructus, 50 g of Schizandrae fructus were added to distilled water, which was then heated at 80oC for 2 h. The sample was then concentrated by rotary evaporation and lyophilized. The resulting powder, which weighed 14.15 g (a yield of 28.3%), was diluted to the required concentrations using autoclaved distilled water and filtered through a 0.22μm syringe filter prior to use.
3. MTT cytotoxicity assay
The cytotoxicity of Schizandrae fructus was determined according to a previously described method (Chang HK et al., 2005).
Briefly, mouse BV2 microglial cells were cultured in 96-well plates containing a final volume of 100μl of culture medium per well. The cells were then treated with the aqueous extract of Schizandrae fructus at concentrations of 1μg/ml, 10μg/ml, 100 μg/ml, and 1,000μg/ml for 24 h. The cells in the control group were left untreated. Next, 10μl of the MTT labeling reagent containing 5 mg/ml MTT in phosphate-buffered saline were added to each well, after which the plates were incubated for 2 h. Subsequently, 100μl of solubulization solution con- taining 10% sodium dodecyl sulfate (SDS) in 0.01 M hydrochloric acid was added to each well, and the cells were incubated for another 12 h. The absorbance was then measured using a microtiter plate reader (Bio-Rad, Hercules, CA, USA) at a test wavelength of 595 nm with a reference wavelength of 690 nm.
The optical density (O.D.) was calculated as the difference between the absorbance at the reference wavelength and the test wavelength. The percent viability was calculated as follows: (O.D.
of drug-treated sample/control O.D.)×100.
4. RNA isolation and RT-PCR
To evaluate the expression of COX-2 and iNOS mRNA, RT-PCR was conducted using a previously described method (Chang HK et al., 2005). Briefly, the total RNA was isolated from Mouse BV2 microglial cells using RNAzolTMB (TEL-TEST, Friendswood, TX, USA). Next, 2μg of RNA and 2μl of 50 nM
random hexamers (Promega, Madison, WI, USA) were added, after which the mixture was heated at 65oC for 10 min. One μl of 10 units AMV reverse transcriptase (Promega), 5μl of 2.5 mM dNTP (Promega), 1μl of 40 units RNasin (Promega), and 8μl of 5×AMV RT buffer (Promega) were then added to the mixture, and the final volume was brought to 40μl volume with diethylpyrocarbonate (DEPC)-treated water. The reaction mixture was then incubated at 42oC for 2 h.
PCR amplification was performed in a reaction mixture with a final volume of 40μl that contained 1μl of the appropriate cDNA, 0.5μl of each set of primers at a concentration of 10 pM, 4μl of 10×RT buffer, 1μl of 2.5 mM dNTP, and 0.2 units of Taq DNA polymerase (Takara, Shiga, Japan). For COX-2, the primer sequences were 5’-CCAGATGCTATCTTT- GGGGAGAC-3’ (a 23-mer sense oligonucleotide) and 5’-CTT- GCATTGATGGTGGCTG-3’ (a 19-mer anti-sense oligonucleo- tide). For iNOS, the primer sequences were 5’-CAAGAGTTTGA- CCAGAGGACC-3’ (a 21-mer sense oligonucleotide) and 5’-TG- GAACCACTCGTACTTGGGA-3’ (a 21-mer anti-sense oligonu- cleotide). For cyclophilin, the internal control used in the study, the primer sequences were 5'-ACCCCACCGTGTTCTTCGAC-3’ (a 20-mer sense oligonucleotide) and 5'-CATTTGCCATGGACAA- GATG-3’ (a 20-mer anti-sense oligonucleotide).
For COX-2 and iNOS, PCR was conducted by subjecting the samples to the following conditions in a PTC-0150 MiniCycler (Bio-Rad): initial denaturation at 94oC for 5 min, followed by 30 amplification cycles consisting of denaturation at 94oC for 45 sec, annealing at 60oC for 45 sec, and extension at 72oC for 45 sec, with an additional extension step at the end of the procedure at 72oC for 10 min. For cyclophilin, PCR was conducted by subjecting the samples to the following conditions: initial denaturation at 94oC for 5 min, followed by 25 amplification cycles consisting of denaturation at 94oC for 30 sec, annealing at 58oC for 45 sec, and extension at 72oC for 45 sec, with an additional extension step at the end of the procedure at 72oC for 10 min. The final amount of RT-PCR product for each of the mRNA species was calculated densitometrically using Molecular AnalystTM version 1.4.1 (Bio-Rad).
5. Western blot analysis
The protein expression was determined according to a pre-
viously described method (Chang HK et al., 2005). Briefly, mouse BV2 microglial cells were lysed in ice-cold whole cell lysate buffer containing 50 mM HEPES (pH 7.5), 150 mM NaCl, 10%
glycerol, 1% Triton X-100, 1.5 mM magnesium chloride hexa- hydrate, 1 mM ethyleneglycol-bis-(β-aminoethyl ether)-N,N'- tetraacetic acid (EGTA), 1 mM phenylmethylsulfonyl fluoride (PMSF), 2μg/ml leupeptin, 1μg/ml pepstatin, 1 mM sodium ortho vanadate, and 100 mM sodium fluoride, after which the mixture was incubated for 30 min at 4oC. The cell debris was then removed by microcentrifugation followed by quick freezing of the supernatant. The protein concentration was measured using a Bio-Rad colorimetric protein assay kit (Bio-Rad). Next, 40μg aliquots of protein were separated on SDS-polyacrylamide gels and then transferred onto a nitrocellulose membrane (Schleicher &
Schuell GmbH, Dassel, Germany) that was subsequently probed with the following primary antibodies: goat COX-2 (1:1,000;
Santa Cruz Biotech, CA, USA), rabbit iNOS (1:1,000; Santa Cruz Biotech), mouse p-ERK (1:1,000; Santa Cruz Biotech), mouse p-p38 (1:1,000; Santa Cruz Biotech), and rabbit p-JNK (1:2,000; Cell Signaling Technology, Beverly, MA, USA).
Horseradish peroxidase-conjugated anti-goat antibody (1:4,000;
Santa Cruz Biotech) for COX-2, anti-mouse antibody (1:2,000;
Santa Cruz Biotech) for p-ERK and p-p38, and anti-rabbit antibody (1:2,000; Santa Cruz Biotech) for iNOS and p-JNK were used as the secondary antibodies. The bands were detected on x-ray film using the enhanced chemiluminescence (ECL) detec- tion system (Amersham Pharmacia Biothech GmbH, Freiburg, Germany). For quantitative analysis, the bands were measured using an Image-ProⓇ computer-assisted image analysis system (Media Cyberbetics Inc., Silver Spring, MD, USA).
6. Measurement of PGE2 synthesis
PGE2 synthesis was assessed using a commercially available PGE2 competitive enzyme immunoassay kit (Amersham Phar- macia Biotech. Inc., Piscataway, NJ, USA) according to a previously described method (Chang HK et al., 2005). Briefly, 100μl of the supernatant of the culture medium and the standards was placed in different wells in the goat anti-mouse IgG-coated microtiter plate provided in the kit. Mouse anti-PGE2
antibody and peroxidase-conjugated PGE2 were then added to each well, after which the plate was incubated at room
Fig. 1. Cytotoxic effect of Schizandrae fructus. Mouse BV2 microglial cells were stained with 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) and then analyzed by microtiter plate reader. (A) Control group, (B) 1μg/ml Schizandrae fructus-treated group, (C) 10μg/ml Schizandrae fructus-treated group, (D) 100μg/ml Schizandrae fructus-treated group, (E) 1,000μg/ml Schizandrae fructus-treated group. The results are presented as the mean±standard error of the mean (S.E.M.). *p<0.05 compared to the control group.
temperature with shaking for 2 h. Next, the wells were drained and washed, and 3,3',5,5'-tetramethylbenzidine/hydrogen peroxide solution was added. The plates were then incubated at room temperature with shaking for 30 min. Finally, the absorbance of the content of each well was measured at a wavelength of 450 nm.
7. Determination of NO production
To determine the effect of the aqueous extract of Schizandrae fructus on NO production, the amount of nitrite in the super- natant was measured using a commercially available NO detec- tion kit (iNtRON, INC., Seoul, Korea) as previously described (Chang HK et al., 2005). Briefly, 100μl of culture supernatant were added to each well, followed by the addition of 50μl of N1 buffer. The plate was then incubated at room temperature for 10 min, after which N2 buffer was added and the plate was incubated at room temperature for 10 min. The absorbance of the content of each well was measured at a wavelength of 540 nm and the nitrite concentration was calculated from a nitrite standard curve.
8. Statistical analysis
Each experiment was repeated at least five times. The results are presented as the mean±standard error of the mean (S.E.M.).
All data were analyzed by one-way ANOVA followed by Duncan's post-hoc test using SPSS. The differences were consi- dered statistically significant at p<0.05.
RESULTS
1. Effect of the aqueous extract of Schizandrae fructus on the viability of mouse BV2 micro- glial cells
Cells cultured in Schizandrae fructus-free media were used as controls. The viability of cells incubated with the aqueous extract of Schizandrae fructus at concentrations of 1μg/ml, 10μg/ml, 100 μg/ml, and 1,000μg/ml for 24 h was 112.82±3.53%, 108.80±
2.31%, 99.52±2.42%, and 95.65±2.24% of the control value, respectively.
These results indicate that the aqueous extract of Schizandrae fructus showed no cytotoxicity at concentrations below 1,000μg/ml
(Fig. 1). Therefore, 10μg/ml and 100μg/ml were used for all subsequent experiments.
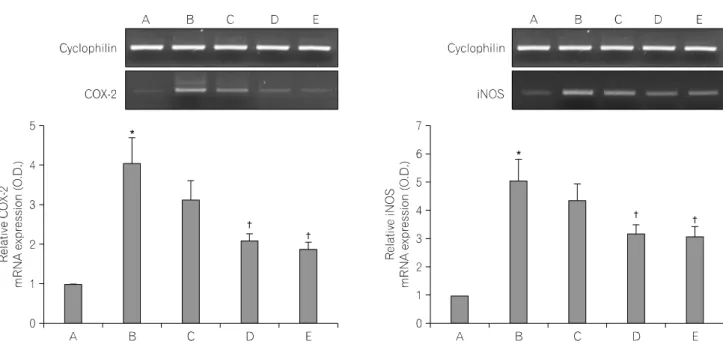
2. Effects of the aqueous extract of Schizan- drae fructus on mRNA expression of COX-2 and iNOS
RT-PCR analysis of the mRNA levels of COX-2 and iNOS was conducted to estimate the relative level of expression of these genes. For the present study, the mRNA levels of COX-2 and iNOS in the control were set as 1.00.
The mRNA level of COX-2 was markedly increased to 4.05±
0.63 following treatment with 100 ng/ml LPS for 24 h. However, the mRNA level of COX-2 was only 3.12±0.47, 2.07±0.19, and 1.88±0.15 in cells that were treated with the aqueous extract of Schizandrae fructus at 10μg/ml, 100μg/ml, and 500μM acetyl- salicylic acid (ASA), respectively, for 1 h prior to treatment with 100 ng/ml LPS for 24 h.
The mRNA level of iNOS following treatment with 100 ng/ml LPS for 24 h was markedly increased to 5.03±0.78. However, the mRNA level of iNOS was only 4.35±0.59, 3.15±0.34, and 3.06±
0.34 in cells that were treated for 1 h with the aqueous extract of Schizandrae fructus at 10μg/ml, 100μg/ml, and 500μM ASA, respectively, prior to treatment with 100 ng/ml LPS for 24
Fig. 2. RT-PCR analysis of the mRNA levels of cyclooxygenase-2 (COX-2) and inducible nitric oxide synthase (iNOS). Mouse BV2 microglial cells were pre-treated for 1 h with Schizandrae fructus at concentrations of 10μg/ml and 100μg/ml, after which they were treated with 100 ng/ml lipopolysaccharide (LPS) for 24 h. The RNA was then extracted and subjected to RT-PCR analysis of COX-2 and iNOS. (A) Control group, (B) LPS-treated group, (C) LPS- and 10μg/ml Schizandrae fructus pre-treated group, (D) LPS- and 100μg/ml Schizandrae fructus pre-treated group, (E) LPS- and 500μM acetylsalicylic acid (ASA) pre-treated group. Cyclophilin was used as the internal control. The results are presented as the mean±standard error of the mean (S.E.M.). *p<0.05 compared to the control group, †p<0.05 compared to the LPS-treated group.
h (Fig. 2).
3. Effects of the aqueous extract of Schizan- drae fructus on the protein expression of COX-2 and iNOS
Analysis of the levels of COX-2 and iNOS protein was conducted to provide an estimate of the relative level of expression of these proteins. In the present study, the protein levels of COX-2 and iNOS in the control cells were set as 1.00.
The level of COX-2 protein increased markedly to 2.65±0.31 following treatment with 100 ng/ml LPS for 24 h. However, the level of COX-2 protein was only 2.75±0.36, 1.89±0.23, and 1.49±0.21 in cells that were treated for 1 h with the aqueous extract of Schizandrae fructus at 10μg/ml, 100μg/ml, and 500 μM ASA, respectively, prior to treatment with 100 ng/ml LPS for 24 h.
The level of iNOS protein following treatment with 100 ng/ml LPS for 24 h increased to 2.30±0.17; however, this level was only 2.03±0.17, 1.48±0.10, and 1.59±0.10 in cells that were treated for 1 h with the aqueous extract of Schizandrae fructus at 10μg/ml,
100μg/ml, and 500μM ASA, respectively, prior to treatment with 100 ng/ml LPS for 24 h (Fig. 3).
4. Effect of the aqueous extract of Schizandrae fructus on PGE2 synthesis
The PGE2 immunoassay revealed that LPS treatment for 24 h increased the amount of PGE2 in the culture medium from 23.05±0.75 pg/ml to 80.80±16.36 pg/ml. However, the levels of PGE2 were only 83.45±5.65 pg/ml, 55.13±6.40 pg/ml, and 41.03±0.95 pg/ml in cells that were treated with the aqueous extract of Schizandrae fructus for 1 h at 10μg/ml, 100μg/ml, and 500μM ASA, respectively, prior to treatment with 100 ng/ml LPS for 24 h (Fig. 4).
5. Effect of the aqueous extract of Schizandrae fructus on NO production
In the NO detection assay, treatment with LPS for 24 h led to an increase in nitrate from 0.90±0.06μM to 15.02±1.34μM.
However, the NO levels were only 11.46±0.90μM, 9.96±0.49 μM, and 9.28±0.53μM when cells were treated for 1 h with
Fig. 3. Western blot analysis of the protein levels of cyclooxygenase-2 (COX-2) and inducible nitric oxide synthase (iNOS). Mouse BV2 microglial cells were pre-treated for 1 h with Schizandrae fructus at concentrations of 10μg/ml and 100μg/ml, after which they were treated with 100 ng/ml lipopolysaccharide (LPS) for 24 h. The protein was extracted and subjected to Western blot analysis of COX-2 and iNOS. (A) Control group, (B) LPS-treated group, (C) LPS- and 10μg/ml Schizandrae fructus pre-treated group, (D) LPS- and 100μg/ml Schizandrae fructus pre-treated group, (E) LPS- and 500μM acetylsalicylic acid (ASA) pre-treated group. Actin was used as the internal control. The results are presented as the mean±standard error of the mean (S.E.M.). *p<0.05 compared to the control group, †p<0.05 compared to the LPS-treated group.
Fig. 4. Measurement of prostaglandin E2 (PGE2) synthesis in mouse BV2 microglial cells. The cells were pre-treated for 1 h with Schizandrae fructus at concentrations of 10μg/ml and 100μg/ml, and then treated with 100 ng/ml lipopolysaccharide (LPS) for 24 h. PGE2 was evaluated by enzyme-linked immunosorbent assay (ELISA). (A) Control group, (B) LPS-treated group, (C) LPS- and 10μg/ml Schizandrae fructus pre-treated group, (D) LPS- and 100μg/ml Schizandrae fructus pre-treated group, (E) LPS- and 500μM acetylsalicylic acid (ASA) pre-treated group. The results are presented as the mean±standard error of the mean (S.E.M.).
*p<0.05 compared to the control group, †p<0.05 compared to the LPS-treated group.
Fig. 5. Measurement of nitric oxide (NO) production in mouse BV2 microglial cells. The cells were pre-treated for 1 h with Schizandrae fructus at concentrations of 10μg/ml and 100μg/ml, after which they were treated with 100 ng/ml lipopolysaccharide (LPS) for 24 h. NO production was then evaluated by microtiter plate reader. (A) Control group, (B) LPS-treated group, (C) LPS- and 10μg/ml Schizandrae fructus pre-treated group, (D) LPS- and 100μg/ml Schizandrae fructus pre-treated group, (E) LPS- and 500μM acetylsalicylic acid (ASA) pre-treated group. The results are presented as the mean±standard error of the mean (S.E.M.).
*p<0.05 compared to the control group, †p<0.05 compared to the LPS-treated group.
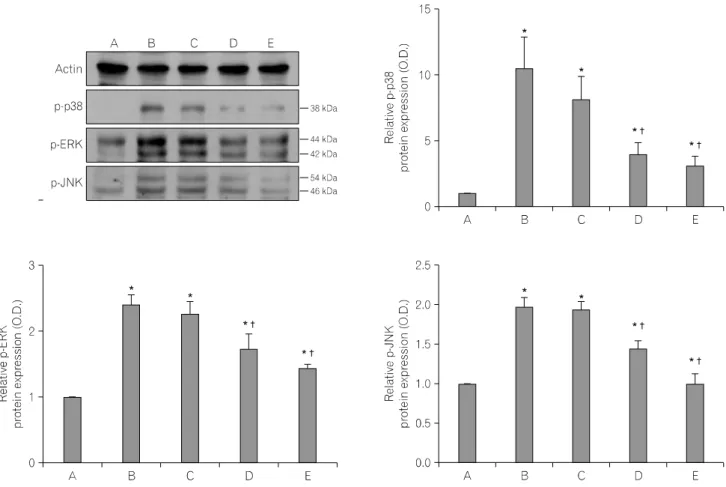
Fig. 6. Effects of the aqueous extract of Schizandrae fructus on phosphorylated mitogen-activated protein kinases (MAPKs). Mouse BV2 microglial cells were pre-treated for 1 h with Schizandrae fructus at concentrations of 10μg/ml and 100μg/ml, after which they were treated with 100 ng/ml lipopolysaccharide (LPS) for 30 min. (A) Control group, (B) LPS-treated group, (C) LPS- and 10μg/ml Schizandrae fructus pre-treated group, (D) LPS- and 100μg/ml Schizandrae fructus pre-treated group, (E) LPS- and 500μM acetylsalicylic acid (ASA) pre-treated group. Actin was used as the internal control.
the aqueous extract of Schizandrae fructus at 10μg/ml, 100μg/ml, and 500μM ASA, respectively, prior to treatment with 100 ng/ml LPS for 24 h (Fig. 5).
6. Effects of the aqueous extract of Schizan- drae fructus on phosphorylated MAPKs in mouse BV2 microglial cells
Analysis of the levels of p-p38, p-ERK, and p-JNK protein was conducted to provide an estimate of the relative level of expression of these proteins. In the present study, the protein levels of p-p38, p-ERK, and p-JNK in the control cells were set as 1.00.
The level of p-p38 protein increased markedly to 10.42±2.40 following treatment with 100 ng/ml LPS for 24 h. However, the
level of p-p38 protein was only 8.07±1.78, 3.96±0.88, and 3.12±0.69 in cells that were treated for 1 h with the aqueous extract of Schizandrae fructus at 10μg/ml, 100μg/ml, and 500 μM ASA, respectively, prior to treatment with 100 ng/ml LPS for 24 h.
The level of p-ERK protein following treatment with 100 ng/ml LPS for 24 h increased to 2.41±0.15; however, this level was only 2.26±0.20, 1.73±0.22, and 1.43±0.07 in cells that were treated for 1 h with the aqueous extract of Schizandrae fructus at 10μg/ml, 100μg/ml, and 500μM ASA, respectively, prior to treatment with 100 ng/ml LPS for 24 h.
The level of p-JNK protein following treatment with 100 ng/ml LPS for 24 h increased to 1.99±0.11; however, this level was only 1.95±0.10, 1.44±0.11, and 1.00±0.13 in cells that were
treated for 1 h with the aqueous extract of Schizandrae fructus at 10μg/ml, 100μg/ml, and 500μM ASA, respectively, prior to treatment with 100 ng/ml LPS for 24 h (Fig. 6).
DISCUSSION
Inflammation activates astrocytes, microglia, and macrophages, which release diverse inflammatory mediators such as TNF-α, prostanoids, interleukins, and NO (Kota et al., 2006). The levels of their presence in tissues indicate the types and intensities of inflammation. Of these, microglial cells are involved in both normal physiology and maintenance as well as pathophysiology by playing a pivotal role in immune response of the central nervous system (Nikodemova et al., 2006). So, many researchers have an interest in relation between inflammation and microglia.
Overexpression of COX-2 activity in vivo has been reported in chronic inflammatory conditions such as rheumatoid arthritis (Kang RY et al., 1996; Crofford et al., 2000). Increased COX-2 expression also results in the increase in prostaglandin production that is associated with inflammation. Thus, it is well known that specific COX-2 inhibitors attenuate the symptoms of inflamma- tion (Crofford et al., 2000). The results of the present study indicated that the aqueous extract of Schizandrae fructus inhibited LPS-stimulated enhancement of COX-2 enzyme activity and PGE2 synthesis in mouse BV2 microglial cells.
During macrophage activation, iNOS mediates production of NO, which causes cell injury through the generation of reactive radicals such as peroxynitrite (Lee AK et al., 2003). The formation of large amounts of NO by iNOS accounts for the cytotoxicity of activated macrophages, and NO plays an important role in inflammation and host-defense response (Nathan et al., 1994).
Szabo reported that agents that inhibit the activity of iNOS may be useful in the development of therapies that focus on many inflammatory processes (Szabo, 1995). In the present study, we showed that the aqueous extract of Schizandrae fructus inhibited LPS-stimulated enhancement of iNOS enzyme activity and NO production in mouse BV2 microglial cells. So, we suggested that Schizandrae fructus might help as factor inhibiting COX-2 and iNOS expression in inflammatory response.
Co-regulation of COX-2 and iNOS has been demonstrated in several inflammatory conditions (Salvemini et al., 1995; Yun KJ
et al., 2008). The results of this study showed that LPS treatment enhanced the expression of COX-2 and iNOS, which resulted in increased PGE2 synthesis and NO production in mouse BV2 microglial cells. NO modulates the activity of COX-2 in a cyclic guanosine monophosphate (cGMP)-independent manner, and NO plays a critical role in the release of PGE2 via the direct activation of COX-2 (Salvemini et al., 1995). COX also affects NOS activity through COX metabolites. Prostanoids may regulate iNOS expression and NO production during the inflammatory response in vivo (Posadas et al., 2000). Additionally, immunosuppressive drugs such as cyclosporine, tacrolimus, methotrexate, and azathi- oprine reduce PGs and NO production (Dusting et al., 1999;
Sands, 2000). Inhibition of these enzyme activities attenuates the symptoms of inflammation.
MAPKs play a critical role in the regulation of cell growth and differentiation, particularly in response to cytokines and stressors (Johnson et al., 2002; Kim JH et al., 2006). p38 MAPK mediates inflammatory responses in various cell types, JNK is essential mediators of relevant proinflammatory functions in microglia, and ERK plays a key role in the down-regulation of iNOS in interferon-γ-stimulated microglia (Chen CC et al., 1999; Park JS et al., 2005; Waetzig et al., 2005). Phosphorylation of MAPKs induces activation of MAPKs, which then regulates the expression of COX-2 or iNOS (Kim JH et al., 2006; Pan et al., 2006). Pan et al.(2006) demonstrated that LPS-induced activation of ERK increased the protein levels of iNOS and COX-2. Furthermore, Kim JH et al.(2006) reported that blockage of p38 MAPK activation in LPS-stimulated raw 264.7 cells reduced inflamma- tion through inhibition of NO production and iNOS expression.
The results of the present study demonstrated that the upregula- tion of phosphorylated forms of MAPKs in the mouse BV2 microglial cells, including p38, ERK, and JNK, by LPS was decreased in response to pre-treatment with the aqueous extract of Schizandrae fructus. As a result, the protein levels of COX-2 and iNOS were also inhibited. These results suggest that Schizandrae fructus inhibits MAPKs activation, which results in the suppression of COX-2 and iNOS expressions.
Natural products have are often useful as alternative medicines with very little side effects. As a result, many available anti- infective, anti-cancer, immune suppressive, and anti-inflammatory drugs have been derived from natural products (Buss et al., 2004;
Butler, 2004). Schizandrae fructus is a medicinal fruit that has long been used for the treatment of dyspnea, cough, mouth dryness, amnesia, and nocturnal emission (Zhu et al., 1999). Schizandrae fructus contains five lignans, schizandrin, gomisin A, gomisin N, gomisin C, and schizandrin C, of which schizandrin has been reported to exert an anti-inflammatory effect on carrageenan- induced paw edema and LPS-treated macrophage cells (Guo et al., 2008).
In this study, we demonstrated that the aqueous extract of Schizandrae fructus exerted anti-inflammatory, and that these effects are relation with suppression of COX-2 and iNOS expression via blockage of the activation of MAPKs. Based on the present results, Schizandrae fructus can be considered as an anti-inflammatory agent for the treatment of brain inflammation.
ACKNOWLEDGEMENTS
This work was supported by 2nd Excellent Research Article Scholarship from the Graduate School of Kyung Hee University at 2007.
REFERENCES
Bredt DS, Snyder SH (1994) Nitric Oxide: a physiologic messenger molecule. Annu. Rev. Biochem. 63:175-195.
Buss DB, Butle MS (2004) A new model for utilising chemical diversity from natural sources. Drug Dev. Res. 62:362-370.
Butler MS (2004) The role of natural product chemistry in drug discovery. J. Nat. Prod. 67:2141-2153.
Chang HK, Yang HY, Lee TH et al. (2005) Armeniacae semen extract suppresses lipopolysaccharide-induced expressions of cyclooxyge- nase-2 and inducible nitric oxide synthase in mouse BV2 micro- glial cells. Biol. Pharm. Bull. 28:449-454.
Chen CC, Wang JK (1999) p38 but not p44/42 mitogen-activated protein kinase is required for nitric oxide synthase induction mediated by lipopolysaccharide in raw 264.7 macrophages. Mol.
Pharmacol. 55:481-488.
Crofford LJ, Lipsky PE, Brooks P et al. (2000) Basic biology and clinical application of specific cyclooxygenase-2 inhibitors. Art- hritis Rheum. 43:4-13.
Dusting GJ, Akita K, Hickey H et al. (1999) Cyclosporin A and tacrolimus (FK506) suppress expression of inducible nitric oxide synthase in vitro by different mechanisms. Br. J. Pharmacol.
128:337-344.
Guha M, Mackman N (2001) LPS induction of gene expression in
human monocytes. Cell Signal. 13:85-94.
Guo LY, Hung TM, Bae KH et al. (2008) Anti-inflammatory effects of schisandrin isolated from the fruit of Schisandra chinensis Baill. Eur. J. Pharmacol. 591:293-299.
Ha SK, Lee P, Park JA et al. (2008) Apigenin inhibits the production of NO and PGE2 in microglia and inhibits neuronal cell death in a middle cerebral artery occlusion-induced focal ischemia mice model. Neurochem. Int. 52:878-886.
Johnson GL, Lapadat RM (2002) Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases.
Science 298:1911-1912.
Kang RY, Freire-Moar J, Sigal E et al. (1996) Expression of cyclooxgenase-2 in human and an animal model of rheumatoid arthritis. Br. J. Rheumatol. 35:711-718.
Kim JH, Kim DH, Baek SH et al. (2006) Rengyolone inhibits inducible nitric oxide synthase expression and nitric oxide production by down-regulation of NF-κB and p38 MAP kinase activity in LPS-stimulated raw 264.7 cells. Biochem. Pharmacol.
71:1198-1205.
Kota RS, Rutledge JC, Gohil K et al. (2006) Regulation of gene expression in raw 264.7 macrophage cell line by interferon-γ.
Biochem. Biophys. Res. Commun. 342:1137-1146.
Lee AK, Sung SH, Kim YC et al. (2003) Inhibition of lipopoly- saccharide-inducible nitric oxide synthase, TNF-α and COX-2 expression by sauchinone effects on I-κBα phosphorylation, C/EBP and AP-1 activation. Br. J. Pharmacol. 139:11-20.
Liu B, Hong JS (2003) Role of microglial in inflammation-mediated neurodegenerative diseases: mechanisms and strategies for thera- peutic intervention. J. Pharmacol. Exp. Ther. 304:1-7.
Mitchell JA, Larkin S, Williams TJ (1995) Cyclooxygenase-2:
regulation and relevance in inflammation. Biochem. Pharmacol.
50:1535-1542.
Moon DO, Choi YH, Kim ND et al. (2007) Anti-inflammatory effects of β-lapachone in lipopolysaccharide-stimulated BV2 microglia. Int. Immunopharmacol. 7:506-514.
Nathan C, Xie QW (1994) Nitric oxide synthases: roles, tolls, and controls. Cell 78:915-918.
Nikodemova M, Duncan ID, Watters JJ (2006) Minocycline exerts inhibitory effects on multiple mitogen-activated protein kinases and IκBα degradation in a stimulus-specific manner in microglia. J. Neurochem 96:314-323.
Pan MH, Lai CS, Wang YJ et al. (2006) Acacetin suppressed LPS-induced up-expression of iNOS and COX-2 in murine macrophages and TPA-induced tumor promotion in mice.
Biochem. Pharmacol. 72:1293-1303.
Park JS, Woo MS, Kim SY et al. (2005) Repression of interferon-γ- induced inducible nitric oxide synthase (iNOS) gene expression in microglia by sodium butyrate is mediated through specific
= 국문초록 =
오미자는 호흡곤란, 기침, 목마름, 기억상실, 그리고 야뇨증 등에 약효를 가지고 있어 치료를 위해 오랫동안 사용된 생약제이다. 천식, 항산화, 간독성, 당뇨 등의 다양한 약효가 보고되고 있지만 항염증과 항진통의 기전에 관한 연구 는 미흡한 실정이다. 따라서 본 연구에서는 생쥐 BV2 소교세포에서 lipopolysaccharide (LPS)로 유도한 염증반응 시 오 미자추출물의 효과를 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) 분석법, 역전사 중합효소 연쇄반 응(RT-PCR), Western blot, prostaglandin E2 (PGE2) 면역분석, 그리고 산화질소(NO) 검출법을 이용하여 조사하였다. 오미 자 추출물은 LPS로 자극된 BV2 소교세포에서 증가된 cyclooxygenase-2 (COX-2)와 inducible nitric oxide synthase (iNOS)의
발현을 억제시킴으로써 PGE2 합성과 NO 생성을 억제하였다. 또한 오미자 추출물은 인산화된 mitogen- activated
protein kinases (MAPKs) 발현도 억제시켰다. 이러한 결과는 오미자가 MAPKs의 하향조절을 통해 COX-2와 iNOS 발현 을 억제시켜 항염증과 항진통효과를 나타낼 수 있음을 나타낸다.
중심단어: 오미자, Mitogen-activated protein kinases, 산화질소, 프로스타글란딘2 inhibition of ERK signaling pathways. J. Neuroimmunol. 168:
56-64.
Park SH, Han JH (2004) A study of medicinal plants for applications in functional foods I. Effects of Schizandrae fructus on the regional cerebral blood flow and blood pressure in rats. J. Korean Soc. Food Sci. Nur. 33:34-40.
Posadas I, Terencio MC, Guillén I et al. (2000) Co-regulation between cyclo-oxygenase-2 and inducible nitric oxide synthase expression in the time-course of murine inflammation. Naunyn Schmiedebergs Arch. Pharmacol. 361:98-106.
Salvemini D, Settle SL, Masferrer JL et al. (1995) Regulation of prostaglandin production by nitric oxide; a in vivo analysis. Br.
J. Pharmacol. 114:1171-1178.
Sands BE (2000) Therapy of inflammatory bowel disease. Gastroen- terology 118:S68-S82.
Surh YJ, Chun KS, Cha HH et al. (2001) Molecular mechanisms underlying chemopreventive activities of anti-inflammatory phy- tochemicals: down-regulation of COX-2 and iNOS through
suppression of NF-κB activation. Mutat. Res. 480-481:243-268.
Szabo C (1995) Alternations in nitric oxide production in various forms of circulatory shock. New Horiz. 3:2-32.
Waetzig V, Czeloth K, Hidding U et al. (2005) c-Jun N-terminal kinases (JNKs) mediate pro-inflammatory actions of microglia.
Glia 50:235-246.
Xu M, Wang G, Xie H et al. (2008) Pharmacokinetic comparisons of schizandrin after oral administration of schizandrin monomer, Fructus Schisandrae aqueous extract and Sheng-Mai-San to rats. J.
Ethnopharmacol. 115:483-488.
Yun KJ, Koh DJ, Kim SH et al. (2008) Anti-inflammatory effects of sinapic acid through the suppression of inducible nitric oxide synthase, cyclooxygase-2, and proinflammatory cytokines expre- ssions via nuclear factor-κB inactivation. J. Agric. Food Chem.
56:10265-10272.
Zhu M, Lin KF, Yeung RY et al. (1999) Evaluation of the protective effects of Schisandra chinensis on Phase I drug metabolism using a CCl4 intoxication model. J. Ethnopharmacol. 67:61-68.