pISSN 1598-642X eISSN 2234-7305
Aspergillus 속 곰팡이를 이용한 액체종국 제조 및 밀누룩의 품질특성
최정실·정석태·김주연·최지호·최한석·여수환*
농촌진흥청 국립농업과학원 농식품자원부 발효이용과
Received : October 7, 2011 / Revised : November 2, 2011 / Accepted : Decembe 3, 2011
Quality Characteristics of wheat Nuruk and Optimum Condition of Liquid Starters for Aspergillus sp.
Choi, Jeong-Sil, Seok-Tae Jung, Joo-Yeon Kim, Ji-Ho Choi, Han-Seok Choi, and Soo-Hwan Yeo*. Fer- mentation & Food Processing Division, Department of Agro-food Resource, NAAS, RDA, Suwon 441-853, Korea
− This study focus was primarily the development of liquid starters for Aspergillus oryzae and Aspergillus niger prepared with wheat bran as a low cost culture medium. For the preparation of the liquid media wheat bran was added at rates of 0, 5, 10, 15 and 20% and the Aspergillus sp. strains were then inoculated to these prepared broths. The results indicated that the more that wheat bran was contained in the medium, the more mycelia was produced for A. oryzae and A. niger. The highest enzyme activities were obtained with a 10~15%
adding rate of wheat bran for both strains. Changes in the enzyme activities of the liquid starters during vari- ous incubation times (0, 24, 48, 72 and 96 hrs), indicated that the highest enzyme activities were seen between 48 and 72 hrs of culture. In addition, a comparative study was carried out on the production of enzymes using wheat as a substrate in nuruk, with liquid starter made from fermented agents according to the same concentra- tions used with the wheat bran. The pH, acidity, amino acidity, reducing sugar content and enzyme activity (α- amylase, glucoamylase, acidic protease) of wheat nuruk made with liquid starter were compared with those of wheat nuruk made with solid starter. The results suggest that the liquid starter is superior in both cases.
Keywords: Aspergillus sp., mycelium, wheat nuruk, liquid starter, enzyme activity
서 론
오랜 세월 동안 탁·약주의 발효제로 사용된 누룩은 날곡 류를 거칠게 빻아 성형한 조곡으로서 자연계 미생물의 착생 에 의해 제조되어 왔다[24, 27]. 전통누룩 제조 시, 인위적으 로 균주를 접종하지 않아도 곰팡이, 효모, 유산균 등의 유용 미생물뿐만 아니라 잡균도 생육하여 위생적으로 좋지 않으 며, 제품의 품질 또한 균일하지 못하다[21 ,27]. 이러한 문제 점을 해결하기 위해, 증자한 곡류에 순수 배양한 누룩 곰팡 이를 인위적으로 접종·배양한 일본식 입국을 사용하게 되 었다[25]. 입국은 전분과 단백질분해 효소를 분비하고 유기산 과 향미 성분들을 생성함으로써 술덧의 주질을 결정짓는 중 요한 인자이다[12, 21, 23]. 이때 입국의 종균 조제는 주류 및 장류 등의 전통발효식품에서 전분 및 단백질분해 등에 필요 한 효소를 생산할 수 있는 곰팡이를 고체배지에 접종하여 포 자가 충분히 형성된 후, 건조시켜 제조한 것이다[9, 23]. 그 러나, 고체배지에 배양할 경우, 원료 처리 조작의 복잡성, 위 해 미생물의 오염, 제국실의 적절한 온·습도 및 저장관리
기술 등의 여러 가지 문제점이 보고되었다[6, 9, 13]. 액체배 양은 잡균번식에 비교적 안전하고 공정을 조절하거나 세포 외 효소, 균사체 및 포자 회수가 용이한 장점이 있다[2, 6].
한편, 전통누룩 제조에 가장 많이 사용하는 밀기울은 제분 과정에서 약 15~20% 정도 생산되며 영양학적 가치는 높지만 식품소재로서 활용의 예는 미흡하다. 그러나, 밀기울에서 약 40~50% 섬유소와 무기질 및 비타민 B가 함유된 것이 알려지 면서 식이섬유가 보강된 기능성 식품으로 이용되고 있다[14, 19]. 또한, 무기질과 비타민 B가 있는 밀기울은 천연배지의 좋 은 원료로서 효소 생산이 우수하다고 알려져 있다[3, 7].
본 연구에서는 1,000년 전부터 우리 조상들이 발효제로 누룩을 사용하였고, 이 발효제는 밀과 밀 가공 부산물인 밀 기울을 이용하여 누룩 곰팡이를 쉽게 배양할 수 있는 액체 종국의 최적 배양조건 규명과 더불어 제조된 밀누룩의 품질 특성을 조사하였다.
재료 및 방법 사용 균주 및 액체배지 조성
본 실험에서 사용된 곰팡이는 국립농업과학원 발효이용과 에 보관중인 A. oryzae(RlB 1353)와 A. niger(RlB 1061)를
*Corresponding author
Tel: +82-31-299-0580, Fax: +82-31-299-0554 E-mail: [email protected]
사용하였다. 사용된 액체배지는 다음과 같이 제조하였다. 밀 기울을 분쇄하여 20-mesh sieve를 통과시킨 후, 증류수 100 mL에 대한 무게비율로 분쇄된 밀기울을 0, 5, 10, 15 및 20% (w/v)되게 첨가하여 121oC에서 25분간 고압 멸균하였 다. A. oryzae와 A. niger를 각각 1백금이 접종하고 30oC, 100 rpm으로 24시간 전배양한 후, 본배양 배지에 5% 접종 하여 48시간 배양하였다(Fig. 1-A).
밀누룩 제조 및 배양조건
밀누룩 제조는 밀을 깨끗이 씻어 3시간 침지하였으며 1시 간 물빼기를 한 후, 50 g씩 삼각 플라스크에 넣고 121oC에 서 25분간 고압 멸균하였다. 액체 배양액을 건조 통밀(수분 7.78±0.2%)의 무게비율로 1, 3, 5, 10%로 접종하여 30oC에 서 40시간 동안 배양시간 별(0, 15, 20, 36, 38, 40 hrs)로 경시적인 변화를 살펴보았다(Fig. 1-B).
균사체량 분석
액체종국의 최적 원료 배합비율과 배양조건을 구명하기 위해, 누룩곰팡이의 균사체량을 분석하였다. 50oC에서 미리 건조한 여과지(Adventec no. 2, Japan)를 이용하여 배양액을 여과하고 증류수로 3회 세척하여 가용성 물질을 제거하였다.
균사체를 50oC에서 항량이 될 때까지 건조하여 균사체량을 측정하였다.
액체종국의 효소활성 분석
액체종국이 가지는 효소활성은 배양액을 조효소액으로 하 여 일본 국세청소정분석법[1]에 따라 각각의 효소활성(α- amylase, glucoamylase, acidic protease)을 분석하였다.
밀누룩의 이화학적 특성분석
배양한 밀누룩 20 g에 물 100 mL을 넣고 실온에서 3시간 침출한 후, 이 여과액으로 pH, 산도, 아미노산도[17] 및 환 원당[16]을 분석하였다. pH는 pH meter(TGA, JP/HM-21P)
로 측정하였고 산도는 여과액 10 mL를 삼각 플라스크에 넣 고 혼합 지시약 1~2 방울을 떨어뜨려 0.1 N NaOH로 적정 한 값을 표시하였다. 아미노산도는 여과액 10 mL를 취하여 삼각 플라스크에 넣고 페놀프탈레인 지시약 3~4방울을 떨어 뜨려 혼합한 후, 0.1 N NaOH로 엷은 분홍색이 나올 때까지 적정한 다음, 중성포르말린 용액 5 mL를 넣어 원래 색으로 돌아오게 하고 0.1 N NaOH로 엷은 분홍색이 나올 때까지 적정하여 그 값을 표시하였다. 환원당은 여과액을 취하여 DNS(Dinitrosalicylic acid)법을 일부 변경하여 분석하였다 [16]. 즉, 25 mL 메스플라스크에 시료 당 용액 1 mL(시료희 석액 1 mL 중에 당 0.2~2 mg 함유)와 DNS시약 3 mL를 넣 고, 5분간 중탕·냉각시킨 후, 물을 가하여 25 mL로 채워 흡광도(550 nm)를 측정하였고, 환원당 함량은 glucose 표준 곡선으로 계산하였다.
밀누룩의 효소활성 분석
밀누룩 10 g에 염화나트륨 5 g를 증류수에 녹인 후, 0.2 M 초산 완충액(pH 5.0) 50 mL를 넣고 염화나트륨 용액 50 mL 를 가해 실온(15~20oC)에서 3시간 침출·여과한 액을 조효 소액으로 하여 일본 국세청소정분석법[1]에 따라 각각의 효 소활성(α-amylase, glucoamylase, acidic protease)을 분석하 였다.
결과 및 고찰
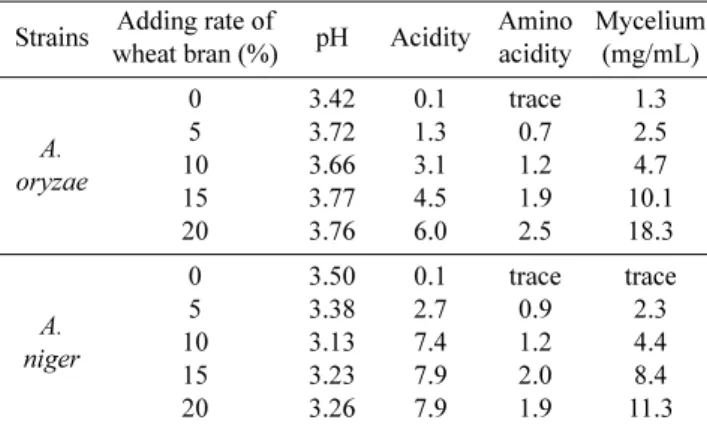
밀기울 첨가비율에 따른 액체종국의 배양학적 특성 술을 빚을 때 당화제로 사용하는 떡 누룩(병국)을 사용할 때마다 밤알 크기로 쪼개는 문제점을 해결하기 위해, 액체 종국의 최적조건을 검토하였다. 우선, 액체배지에 밀기울 농 도(0, 5, 10, 15, 20%) 별로 액체종국을 만들고, 여기에 Aspergillus sp. 곰팡이를 각각 접종한 후, 이들의 배양학적 특성을 Table 1에 나타내었다. pH의 경우, Aspergillus 속의 두 균주(A. oryzae, A. niger) 모두 pH 4.0 이하로 적정하였
Fig. 1. Wheat liquid starter and wheat nuruks manufactured in this study. Symbols : A; Wheat liquid starter, B; wheat nuruks inocu- lated with different inoculation rate of starter.
고, 산도와 아미노산도는 영양분이 풍부한 밀기울의 첨가량 이 많을수록 경시적으로 증가하는 것을 알 수 있었다. 특히, 밀기울 농도가 높을수록 A. niger는 A. oryzae보다 pH가 낮 았고, 산도는 높게 나타났다. 이것으로 보아 A. niger가 생 성하는 구연산이 배양액에 함유된 포도당으로부터 생성되는 것을 알 수 있었다[5]. 액체종국의 균사체 생성능을 살펴보 면, A. oryzae와 A. niger는 밀기울 농도가 높아질수록 균사 체 생성량도 증가하였다. Table 1에서 밀기울 농도가 15%
이상일 때, 균사체 생성량 또한 급격히 증가되었다. 이러한 결과는 서 등[22]의 결과와 유사한 것으로 Aspergillus 속 곰 팡이를 이용한 액체종국제조에서 밀기울이 영양원으로서 역 할을 하는 것으로 보인다.
밀기울 농도 및 배양시간에 따른 액체종국의 효소활성 변화 서로 다른 두 종류의 Aspergillus 속 곰팡이(A. oryzae, A.
niger)를 대상으로 밀기울 농도별 효소활성 결과를 Table 2 에 나타내었다. 각 효소의 활성은 밀기울 첨가비율에 따라 서로 다른 양상을 나타내었다. 황국균인 A. oryzae는 α- amylase가 15%에서, glucoamylase와 acidic protease는 10%
첨가구에서 가장 높은 효소 활성을 나타내었고, 흑국균(A.
niger)인 α-amylase는 15%, glucoamylase는 10%, acidic
protease는 5% 첨가구에서 효소활성이 가장 높은 결과를 보 였다. 따라서, 다양성과 차별성을 갖는 액체종국 제조는 사 용 균주 또는 효소 종류에 따라 첨가하는 밀기울 농도가 달 라지는 것을 알 수 있다. Table 1에 나타낸 균사체 생성량을 비교해 보면 밀기울 20% 첨가구에서 균사체 생성량은 많지 만 효소활성이 낮은 것으로 보아(Table 2) 균사체 생성량과 액체종국의 효소활성과는 비례하지 않는 것으로 나타났다.
따라서 액체종국의 효소활성을 높이기 위해서는 밀기울의 최적농도를 15%로 첨가한 것이 가장 뛰어난 결과를 나타내 었다. 이와 반대로 밀기울을 많이 첨가하였을 경우, 액체종 국의 효소활성이 급격히 떨어지는 것에 대해서는 원료와 양 조미생물과의 상관관계에 대한 연구가 필요할 것으로 생각 된다. 전분을 분해하는 α-amylase는 전분의 α-1,4 glucoside 결합을 무작위로 가수분해하므로 glucose, maltose를 비롯하 여 다당류를 생산하는 세포외 amylase이다[11, 15]. 특히, A.
oryzae와 A. niger는 15% 밀기울을 첨가한 액체배지에서 최 고의 α-amylase활성을 나타내었다. 그러나, 이들 균간의 α- amylase차는 A. oryzae가 A. niger보다 2.6배의 높은 효소활 성을 가진다. 황국균인 A. oryzae는 많은 양의 세포외 α- amylase를 생산하는 균주로서 산업적으로 유용한 양조미생 물로 보고된 연구 결과도 있다[4].
전통누룩 제조시 가장 많이 사용하는 밀기울의 당화력은 액화효소와 당화효소뿐만 아니라 생전분 분해효소의 역할로 도 중요하다[8]. 생전분 분해력의 지표인 glucoamylase의 효 소 활성은 A. oryzae와 A. niger 모두 10% 밀기울배지에서 높은 활성을 가지고 있었다. 산성 protease활성은 A. oryzae 에서는 10% 밀기울배지에서 최고치(1.54 units/mL)를 보였 고, A. niger는 A. oryzae보다 밀기울 농도가 낮은 5% 배지 에서 가장 높았고(3.05 units/mL), 황국균보다 2배의 acidic protease를 가지고 있었다(Table 2).
배양시간에 따른 효소활성도의 변화
A. oryzae와 A. niger의 밀기울 액체배지에서 배양시간 별 당화효소 활성은 Table 3에 나타내었다. A. oryzae 효소활성 Table 1. Properties of liquid starters on different adding rate of
wheat bran.
Strains Adding rate of
wheat bran (%) pH Acidity Amino acidity
Mycelium (mg/mL)
A.
oryzae
0 5 10 15 20
3.42 3.72 3.66 3.77 3.76
0.1 1.3 3.1 4.5 6.0
trace 0.7 1.2 1.9 2.5
1.3 2.5 4.7 10.1 18.3
A.
niger
0 5 10 15 20
3.50 3.38 3.13 3.23 3.26
0.1 2.7 7.4 7.9 7.9
trace 0.9 1.2 2.0 1.9
trace 2.3 4.4 8.4 11.3
Table 2. Enzyme activities of liquid starter making from differ- ent adding rate of wheat bran.
Strains Adding rate of wheat bran (%)
Enzyme activity (units/mL) α-
amylase glucoamylase acidic protease
A. oryzae
0 5 10 15 20
0.22 1.40 6.64 15.80 0.85
0.09 0.75 1.65 1.56 0.40
0.07 0.80 1.54 0.91 0.11
A. niger
0 5 10 15 20
0.39 1.95 3.53 6.07 0.23
0.30 3.30 4.50 2.73 0.87
0.05 3.05 0.82 0.65 0.22
Table 3. Changes in enzyme activities of wheat bran liquid media during culture
Strains Culture time (hrs)
Enzyme activity (units/mL) α-amylase glucoamylase
A. oryzae
0 24 48 72 96
0.20 2.90 6.70 6.20 5.70
0.10 0.72 1.63 1.58 0.62
A. niger
0 24 48 72 96
0.35 1.90 3.80 4.50 3.30
0.25 2.90 4.70 4.20 1.50
은 경시적으로 증가하여 48시간에 최고치를 보였으며, A.
niger는 72시간 배양 시 가장 높은 활성을 나타내었다. 효소 활성에 따른 최적 배양시간은 곰팡이 종류에 따라 차이가 있 는 것을 알 수 있었다. A. oryzae의 α-amylase 활성은 A.
niger보다 효소 활성이 높은 것으로 알려져 있다[10]. 본 연 구에서도 양조용 황국균인 A. oryzae의 α-amylase 활성은 흑국균인 A. niger보다 24시간 더 일찍 효소 활성을 나타내 었으나 glucoamylase활성은 A. oryzae와 A. niger 두 균주 모두 48시간 배양이 최적이었다.
액체종국 접종량에 따른 일반성분의 특성
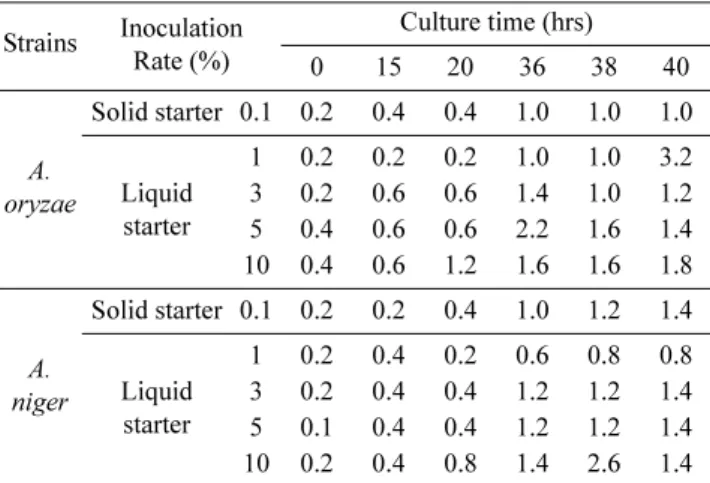
A. oryzae와 A. niger의 액체종국 배양은 10~15% 밀기울 배지와 48~72시간의 배양 최적조건이 규명하였다. 그러나 액체종국의 최적 접종량을 결정하기 위해, 기존의 고체종국 을 대조구로 하여 배양시간 별로 조사하였다. 액체종국을 접 종한 밀누룩의 균주별 배양시간에 따른 산도의 특성은 Table 4에 나타내었다. 전체적으로는 균주 접종 후, 배양시간이 경 과됨에 따라 산도가 높아지는 경향이었다. 대체적으로 20시
간까지는 두 균주간의 변화는 거의 없었지만 배양시간이 늘 어남에 따라 A. oryzae는 5% 이상, A. niger는 3% 이상의 접종량에서 양호하였다. 아미노산도는 접종 후, 급격히 증가 하였고 배양 36 시간째 거의 최고를 나타내었지만 배양시간 이 경과됨에 따라 아미노산도가 3.0이하로 낮아지는 것으로 보아 향후, 주류 제조시 느끼한 맛은 없는 것으로 사료된다 (Table 5). 대조구인 고체종국과 액체종국을 비교해 보면, 두 균주에서 액체종국을 접종한 밀누룩이 고체종국보다 산도나 아미노산도가 높은 것을 알 수 있다. 이것은 고체배지보다 액체배지에 접한 곰팡이 면적이 큰 것으로 인해 생육속도가 증가된 것으로 여겨진다.
밀누룩의 액체종국 접종량별 환원당 생성 특성
배양시간에 따른 균주별 환원당 생성량은 A. oryzae가 배 양 36시간째 3~10% 접종량에서 환원당이 가장 많이 생성 되었다. 따라서, 액체종국의 접종량이 증가할수록 생성된 amylase작용으로 환원당이 증가하는 것을 알 수 있다[20].
A. niger의 경우는 배양 15시간 이후에 접종량에 따른 큰 변 화가 없었지만, 5% 액체종국이 가장 높았다. 결론적으로 액 체종국의 접종량이 많아질수록 당화효소에 의해 전분질이 분 해되어 환원당이 많이 생성된 것으로 이 등[13]의 보고와 유 사한 결과를 얻었다. 대조구인 고체종국과 비교해 보면, A.
oryzae는 배양 15시간 이후부터 액체종국의 접종량이 많아질 수록 환원당 생성량이 높은 것을 알 수 있었다. 고체종국을 38시간 배양한 결과, 환원당 함량이 가장 높았고 액체종국의 환원당은 36시간째 가장 많이 생성하였다. A. niger는 배양 15시간 이후부터 고체종국보다 액체종국에서 환원당이 많이 생성되었다. 따라서, 시간과 인력 등의 경제적 가치뿐만 아 니라 잡균의 오염도가 높은 고체종국보다는 액체종국을 사 용하는 것이 여러 가지 측면에서도 유리하다고 여겨진다.
배양시간에 다른 액체종국의 효소활성
사용된 누룩 곰팡이(A. oryzae, A. niger)의 효소 활성변화 Table 4. Changes in acidity of wheat nuruk on different inocula-
tion rate of liquid starter Strains Inoculation
Rate (%)
Culture time (hrs)
0 15 20 36 38 40
A.
oryzae
Solid starter 0.1 0.2 0.4 0.4 1.0 1.0 1.0
Liquid starter
1 0.2 0.2 0.2 1.0 1.0 3.2 3 0.2 0.6 0.6 1.4 1.0 1.2 5 0.4 0.6 0.6 2.2 1.6 1.4 10 0.4 0.6 1.2 1.6 1.6 1.8
A.
niger
Solid starter 0.1 0.2 0.2 0.4 1.0 1.2 1.4
Liquid starter
1 0.2 0.4 0.2 0.6 0.8 0.8 3 0.2 0.4 0.4 1.2 1.2 1.4 5 0.1 0.4 0.4 1.2 1.2 1.4 10 0.2 0.4 0.8 1.4 2.6 1.4 Table 5. Changes in amino acidity of wheat nuruk on different inoculation rate of liquid starter
Strains Inoculation Rate (%)
Culture time (hrs)
0 15 20 36 38 40
A.
oryzae
Solid starter 0.1 0.2 0.4 1.0 3.8 4.2 4.4
Liquid starter
1 0.4 0.4 0.6 2.6 3.0 2.2 3 0.2 0.4 0.8 2.6 1.8 2.2 5 0.4 0.6 1.0 3.4 3.0 2.2 10 0.6 0.8 1.4 3.6 3.4 3.0
A.
niger
Solid starter 0.1 0.2 0.2 0.2 0.8 0.6 0.6
Liquid starter
1 0.2 0.2 0.4 0.8 0.8 1.0 3 0.2 0.4 0.6 2.2 1.8 2.4 5 0.2 0.4 0.6 2.0 1.8 2.0 10 0.2 0.6 0.8 2.8 3.2 2.8
Table 6. Changes in reducing sugar contents of wheat nuruk on inoculation rate of liquid starter
Strains Inoculation Rate (%)
Culture time (hrs)
0 15 20 36 38 40
A.
oryzae
Solid starter 0.1 1.6 4.1 8.2 13.4 14.0 9.0
Liquid starter
1 5.4 6.1 9.6 18.0 28.5 16.4 3 1.1 6.0 13.6 19.3 14.2 5.4 5 2.2 10.3 12.6 22.0 14.1 13.9 10 1.8 12.7 19.3 20.3 13.1 16.7
A.
niger
Solid starter 0.1 0.6 3.7 4.9 10.3 10.8 9.7
Liquid starter
1 0.6 7.9 7.4 13.3 16.3 14.5 3 0.6 11.5 16.3 5.4 5.0 5.5 5 0.9 14.5 14.3 23.9 23.3 23.4 10 1.0 14.7 8.4 11.1 11.9 11.8
는 20시간 이후부터 급격히 증가하는 것을 볼 수가 있었다 (Fig. 2). 이것으로 보아 배양 20시간까지는 누룩 곰팡이의 포자가 발아하여 밀 입자에 균사가 착생하는 시기라고 생각 된다. 특히, 전분분해 효소인 α-amylase는 전분을 무작위로 끊는 역할을 하고 glucoamylase는 전분의 비환원당 말단에 서 분해하여 포도당을 만드는데 관여하는 효소이다[26]. Fig.
2-A에 나타낸 것처럼 A. oryzae 액체종국의 접종량에 따른 α-amylase활성 변화는 큰 차이가 없었다. 이와 반대로, A.
niger의 α-amylase 활성변화는 액체종국의 접종 농도에 따 라 효소활성에 차이가 있었고, 배양 20~36시간까지는 효소 활성이 급격히 증가하는 양상을 보였다(Fig. 2-B). 누룩 곰 팡이(A. oryzae, A. niger)의 접종량이 많을수록 α-amylase 효소활성이 높아지는 것은 배양기간 중 누룩 곰팡이의 포자 가 발아하여 균사체가 많이 생장한 것으로 여겨진다[18].
2종류의 누룩 곰팡이(A. oryzae, A. niger)로 만든 액체종 국의 glucoamylase 효소활성은 배양 38시간에 가장 높은 효 Fig. 2. Changes in enzyme activity of wheat nuruk on different inoculation rate of liquid starter. Symbols: A; α-amylase activity of wheat nuruk, B; α-amylase activity of wheat nuruk, C; Glucoamylase activity of wheat nuruk, D; Glucoamylase activity of wheat nuruk, E;
Acidic protease activity of wheat nuruk, F; Acidic protease activity of wheat nuruk.
소활성을 보인 후, 감소하는 경향을 보였다(Fig. 2-C, D). 두 효소의 활성을 비교해보면, glucoamylase는 α-amylase보다 2배 이상 높은 효소 활성을 가지고 있었다(Fig. 2).
2종류의 누룩 곰팡이에 대한 acidic protease는 Fig. 1-E, F에 나타내었다. 전반적으로 고체종국과 액체종국간에 acidic protease차이는 없었으나 배양 20시간부터 효소활성이 급격 히 증가하였다. 특히, A. oryzae는 접종 후 38시간(Fig. 2-E) 에, A. niger는 접종 후, 36시간에 최고조에 달하였다(Fig. 2- F). 따라서, acidic protease에 의해 분해된 아미노산은 배양 기간 중 효모의 영양원으로 이용되어 고급 알코올 및 에스 테르 등의 향기성분으로 분해된다. 그 결과, 각 지역에서 생 산된 주류의 차별성이 부각된 것으로 여겨진다[5].
요 약
고역가 액체종국 제조 및 배양기술을 개발하기 위해, Aspergillus 속의 곰팡이(A. oryzae, A. niger)를 이용한 밀기 울 첨가율 별(0, 5, 10, 15 및 20%) 배지를 이용한 액체종 국을 제조하여 이들의 품질 특성을 조사하였다. 액체종국의 일반성분 및 효소활성(glucoamylase, acidic protease)을 조 사한 결과, 배지에 밀기울 첨가량이 많을수록 균사체량이 많 아졌을 뿐만 아니라 산도, 아미노산도도 증가하는 것을 알 수 있었고 액체종국에 첨가하는 밀기울 농도가 달라짐에 따 라 각 균주의 효소활성이 바뀌는 것을 알 수 있었다. 또한, 효소활성에 따른 최적 배양시간은 곰팡이 종류에 따라 차이 가 있었고 A. oryzae와 A. niger 곰팡이의 액체종국 배양은 10~15% 밀기울을 첨가하여 48~72시간 배양하는 최적조건 을 규명하였다. 기존에 사용되었던 고체종국과 비교하여 밀 입국제조에 종국으로써 적합한지를 구명하고자 A. oryzae와 A. niger를 사용한 고체종국을 접종한 밀누룩을 대조구로 하 여 밀기울 10% 액체배지로 만든 액체종국의 접종량 별(1, 3, 5, 10%) 밀누룩의 배양시간 별(0, 15, 20, 36, 38, 40 hrs) 및 이화학적 특성과 효소활성(α-amylase, glucoamylase, acidic protease)변화를 비교하였다. 결론적으로 시간과 인력 등의 경제적 가치가 많이 투입되는 고체종국 보다는 누룩 제 조시 액체종국을 사용하는 것이 여러 가지 측면에서도 유리 하다고 여겨진다.
Acknowledgement
This study was carried out with the support of “Research Program for Agricultural Science & Technology Develop- ment (Project No. PJ007396)”, National Academy of Agri- cultural Science, Rural Development Administration, Repub- lic of Korea.
REFERENCES
1. Brewing Society of Japan. 1993. Regulation for analysis of alcoholic beverages, pp. 221-228.
2. Chandran, S., A. Sumantha, G. Szakacs, A. Pandey. 2005.
Comparative evaluation of neutral protease production by Aspergillus oryzae in submerged and solid-state fermenta- tion. Process Biochemistry 40: 2689-2694.
3. Chung, M. J. 1977. Studies on the proteolytic enzyme pro- duced by Rhizopus japonicus S-62. Korean J. Food Sci.
Technol. 9: 31-35.
4. Ham, Y. T. 1996. The characteristics of Aspergillus oryzae and application it to genetic engineering. Mycology Society News 8: 34-48.
5. Ji, I. S. 1994. New technology in manufacturing of distilled alcohols (So-ju and Ju-jeong). Korean Journal of Micro- biology and Biotechnology. pp. 53-62.
6. Kang, S. G., I. B. Park, and S. T. Jung. 1997. Characteristics of fermented hot pepper soybean paste (Kochujang) pre- pared by liquid beni-koji. Korean J. Food Sci. Technol. 29:
82-89.
7. Kim, K., I. H. Park, and Y. I. Sunwoo. 1991. α-Amylase production of Bacillus natto IAM 1212 in the wheat bran medium. Korean J. Biotechnol. Bioeng. 6: 143-146.
8. Kim, H. S., J. S. Hyun, J. Kim, H. P. Ha, and T. S. Yu. 1998.
Enzymological characteristics and identification of useful fungi isolated from traditional Korean nuruk. Kor. J. Appl.
Microbiol. Biotechnol. 26: 456-464.
9. Kwon, Y. A., and J. K. Chun. 1988. Microcomputer-con- trolled koji incubation system and its application to barley koji manufacture. Korean J. Food Sci. Technol. 20: 326-330.
10. Lee, D. Y. 1967. Studies on the industrialization of the Korean kokja. (I) It’s isolation and physiological charac- teristics of mold from kokja. Kor. Jour. Microbiol. 5: 51-54.
11. Lee, S. H., H. J. Jung, S. H. Yeo, H. S. Kim, and T. S. Yu.
2004. Characteristics of α-amylase of, a new species, Aspergillus coreanus NR 15-1. Korean J. Biotechnol. Bioeng.
19: 301-307.
12. Lee, S. K., and T. S. Lee. 1976. Studies on the effect of seed koji for the soysauce qualities. J. Korean Agricultural Chemical Society 19: 155-161.
13. Lee, T. S., S. O. Park, and S. S. Kung. 1984. Changes of chemical composition during the aging of liquid koji kochujang. Korean J. Food Sci. Technol. 16: 1-6.
14. Lee, Y. T. 2003. Quality characteristics of high-fiber breads added with domestic wheat bran. J. Korean Soc. Agric.
Chem. Biotechnol. 46: 323-328.
15. Mabel, S. H., M. Rodriguez, N. P. Guerra, and R. P. Roses.
2006. Amylase production by Aspergillus niger in submerged cultivation on two wastes from food industries. Journal of Food Engineering 73: 93-100.
16. Miller, G. L. 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal Chem 31: 426-428.
17. National Tax Service Liquors Licence Aid Center. 2009.
Liquors analysis code. pp. 41-42.
18. Oh, K. C., T. J. Yu, and T. Y. Kim. 1988. Studies on degree of kojic mycelial penetration and some characteristics of kojic enzyme in moromi. Kor. J. Mycol. 16: 175-184.
19. Park, Y. H., and B. H. Park. 2000. Changes in nutritional components of toha-jeot with wheat bran during fermenta- tion. Korean J. of Human Ecology 3: 77-89.
20. Rani, G., P. Gigras, H. Mohapatra, V. K. Goswami, and B.
Chauhan. 2003. Microbial α-amylase: a biotechnological perspective. Process Biochemistry 38: 1599-1616.
21. Rha, K. Y. 1989. Seed mold important in brewing. Korean J.
Food & Nutrition 1: 108-110.
22. Seo, J. B., B. R. Jin, S. C. Shin, H. Y. Park, B. Y. Lee, C. K.
Lee, and S. K. Kang. 1995. Development of liquid culture media for the mass production of Beauveria bassiana blastospore. Korean J. Seric. Sci. 37: 172-175.
23. So, M. H. 1993. Characteristics of koji molds isolated from koji-starters for brewing in Korea and Japan. Korean J. Food
& Nutrition 6: 1-7.
24. So, M. H. 1999. Characteristics of a modified nuruk made by inoculation of traditional nuruk microorganisms. Korean J. Food & Nutrition 12: 219-225.
25. So, M. H., and Y. S. Lee. 2009. Effects of culture condition of Rhizopus sp. ZB9 on the production of saccharifying amylase during the preparation of rice koji. Korean J. Food
& Nutri 22: 644-649.
26. Yumiko, Y., T. Susuki, K. Takamine, H. Tamaki, K. Ito, and Y. Sameshima. 2010. Characterization of glucoamylase and α-amylase from Monascus anka: Enhanced production of α- amylase in red koji. Journal of Bioscience and Bioengineer- ing 110: 670-674.
27. Yu, T. S., J. Kim, H. S. Kim, J. S. Hyun, H. P. Ha, and M. G.
Park. 1998. Bibliographical study on microorganisms of traditional Korean nuruk (since 1945). J. Korean Soc. Food Sci. Nutr. 27: 789-799.