Apoptotic Effects of A Cisplatin and Eugenol Co-treatment of G361 Human Melanoma Cells
8
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
(7)
(8)
수치

관련 문서
Taken together, our results show that the activity of withaferin A to modulate multiple components in apoptotic response of human leukemia cells and raise the
Also, the cells were pre-treated with Chaga extracts(125 ㎍/㎖) for 4 h then incubated in the presence of cisplatin (20 μ M) for indicated time periods. Total RNA was then isolated
Results using flow cytometric analysis indicated that treatment with lapathoside A increased the number of apoptotic cells in pancreatic cancer cell lines.. These
S100A8 and S100A9 suppressed the cell survival, and this suppression was found to be associated with apoptosis because they increased the number of apoptotic cells in a dose-
response, both recovery and destruction are induced by redox and cell death signaling pathways to damaged cells and tissues. 4) Such chronic inflammatory response is
Our results indicated that HEABG induced apoptotic cell death and the effect is mediated through a signaling cascade of extrinsic as well as intrinsic caspase pathways..
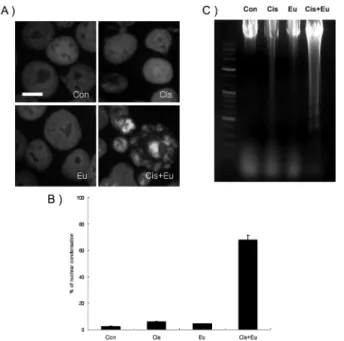
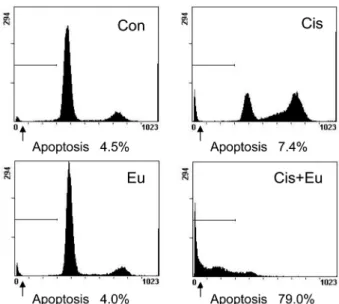
In our current study, we examined whether the cytotoxic effects of eugenol upon human melanoma G361 cells are associated with cell cycle arrest and apoptosis using a range
