유리지방산 유도 지방간세포에서 Isorhamnetin의 PPARα 활성화 작용을 통한 지질 축적 억제 효과
김추숙1․조은희2․최석영1․강민숙3․유리나1
1울산대학교 식품영양학과
2강원대학교 내분비내과
3농촌진흥청 식생활영양과
Inhibitory Effect of Isorhamnetin on Lipid Accumulation in Free Fatty Acid-Induced Steatotic Hepatocytes through the PPARα Pathway
Chu-Sook Kim
1, Eun-Hee Cho
2, Suck-Young Choe
1, Min-Sook Kang
3, and Rina Yu
11
Department of Food Science and Nutrition, University of Ulsan
2
Department of Internal Medicine, Kangwon National University
3
Food and Nutrition Division, National Institute of Agricultural Sciences
ABSTRACT Obesity-induced hepatic lipid accumulation is a crucial factor in the initiation and development of non- alcoholic fatty liver disease. In this study, we investigated the effect of isorhamnetin, a naturally occurring flavonoid in fruits and vegetables, on free fatty acid (FFA)-induced lipid accumulation in hepatocytes. Hepatocytes (Hepa1c1c) were exposed to FFA with or without isorhamnetin, and lipid accumulation was estimated by Oil Red O staining and biochemical measurement. The metabolic genes/proteins involved in lipid metabolism were measured by qRT-PCR and/or western blotting. Isorhamnetin significantly reduced lipid accumulation in FFA-induced hepatocytes, and this was accompanied by modulation of genes involved in lipid metabolism. Isorhamnetin up-regulated expression of fatty acid oxidation-related genes (peroxisome proliferator-activated receptor α (PPARα), carnitine palmitoyltransferase 1, and acyl-CoA oxidase), and down-regulated lipogenic gene expression (sterol regulatory element-binding protein-1, fatty acid synthase, and acetyl-CoA carboxylase). Alterations in the metabolic gene expression induced by isorhamnetin were blunted by PPARα antagonist. These findings indicate that the inhibitory action of isorhamnetin in FFA-induced lipid accumulation in hepatocytes is associated with the PPARα pathway. Isorhamnetin may be useful for protection against obesity-related hepatosteatosis.
Key words: obesity, fatty liver, hepatocyte, lipid metabolism, isorhamnetin
Received 26 April 2018; Accepted 15 June 2018
Corresponding author: Chu-Sook Kim, Department of Food Science and Nutrition, University of Ulsan, Ulsan 44610, Korea E-mail: [email protected], Phone: +82-52-259-2390
서 론
비알코올성 지방간질환(non-alcoholic fatty liver dis- eases, NAFLD)은 간에 중성지방이 축적된 단순 지방간증, 염증으로 발전한 지방간성 염증 및 간섬유화, 간경변증 등을 포함한다(1). 비알코올성 지방간질환은 만성적이고 점진적 인 과정을 통해 진행되기 때문에 초기 가역적인 상태에서 적절한 치료는 지방간 진행과정을 개선하거나 지연시킬 수 있다. 비알코올성 지방간질환의 병변과정에서 대사적으로 주목할 만한 특징은 간세포 내 중성지방의 축적이다(2). 간 세포 내 과량의 중성지방 축적은 간세포 내로 유리지방산 유입의 증가에 기인하는데, 고지방/고열량 식이, 인슐린 저
항성 상태에서 과도한 식이지방 유입의 증가 또는 지방조직 에서 지방분해의 증가로 유리지방산 유입이 증가한다(3). 이 때 유리지방산과 지질대사산물들은 지질세포독성을 야기하 고(4,5), 지방간성 염증 및 간섬유화를 가속화하는 주요인자 로 작용한다(6). 따라서 유리지방산 산화의 증가 또는 지질 합성의 감소 등의 간세포 내 지방산 대사 조절은 비알코올성 지방간질환을 예방하기 위한 전략일 수 있다.
간에서 지질대사를 조절하는 대표적인 전사인자로는 peroxisome proliferator-activated receptor α(PPARα) 와 sterol regulatory element-binding protein-1(SREBP- 1)이 있다. PPARα는 지방산 산화를 조절하는 반면에 SREBP- 1은 지질 합성을 조절한다(7,8). PPARα는 간세포에서 높은 수준으로 발현되며 지질대사와 관련된 유전자 발현을 조절 한다(9). PPARα 활성화는 지방산 수송단백질 및 long-chain acyl-CoA synthetase 발현을 유도하여 미토콘드리아 내로 의 지방산 수송을 증가시키고, acyl-CoA oxidase 1(ACOX1),
carnitine palmitoyl transferase 1(CPT-1)의 발현을 증가 시켜 지방산 산화를 증가시킨다(10). 지방간증 환자의 간에 서 PPARα의 발현 감소가 관찰되고(11), PPARα 항진제는 지방간증 상태를 개선한다고 알려져 있다(12). 한편 SREBP- 1의 활성화는 지질 합성 유전자인 acetyl CoA carboxylase (ACC), fatty acid synthase(FAS)의 발현을 증가시켜 지방 산과 중성지방 합성을 유도하고 지방간증의 진행을 촉진한 다(13). 따라서 이들 지질대사 관련 전사인자 조절작용을 가 진 식품성분은 지방간증의 예방과 치료에 유용할 것으로 생 각된다.
다양한 플라보노이드 성분인 quercetin, naringenin 등은 지방산 산화의 조절자인 PPARα와 CPT-1 발현을 증가시키 거나 SREBP-1을 매개한 지질 합성 경로를 차단함으로써 지 방간증을 억제하고 비알코올성 지방간질환을 개선한다(14, 15). 우리의 선행연구도 quercetin이 미토콘드리아 산화적 대사를 증가시킴으로써 비만으로 유도된 지방간 개선 효과 를 확인한 바 있다(16). Isorhamnetin은 식품 내에 널리 존 재하는 플라보노이드 성분으로 quercetin의 대사산물의 하 나이며, 강력한 항산화/항염증 활성을 나타내고 유전적 및 식이성 비만 쥐에서 인슐린 저항성과 같은 비만성 대사질환 을 개선한다(17). 또한, isorhamnetin의 투여는 혈중 지질 수준의 감소를 나타내고 간 내 중성지방 및 콜레스테롤의 감소를 가져온다고 보고되어 있으며(18), 이는 isorhamne- tin이 비알코올성 지방간질환을 개선할 가능성을 시사한다.
그러나 isorhamnetin의 지방간증 개선 효과가 PPARα 활성 화와 연관되어 있는지는 충분히 알려져 있지 않다. 본 연구 에서는 유리지방산으로 유도된 지방간세포 모델계에서 iso- rhamnetin의 지질 축적 억제 효과와 그 작용기전에 대해 조사하였다.
재료 및 방법
세포배양
Hepa1c1c cell은 10% fetal bovine serum과 10 mg/L penicillin-streptomycin, 2 mg/L gentamicin이 첨가된 Dulbecco’s modified Eagle’s medium(DMEM, Life Tech- nologies, Carlsbad, CA, USA) 배지에서 37°C, 5% CO2의 조건으로 배양하였다. 지질 축적을 유도하기 위해 Hepa1c1c cell은 4×105 cell/well의 세포수로 6-well plate에 분주하 고, 1 mM 유리지방산[2:1 ratio of oleate/palmitate cou- pled to free BSA(molar ratio, 10:1)]과 여러 농도의 iso- rhamnetin(Enzo, Farmingdale, NY, USA)을 24시간 동안 처리하였다. PPARα 저해제인 GW6471(Sigma-Aldrich Co., St. Louis, MO, USA)은 유리지방산과 isorhamnetin을 처리하기 1시간 전에 전처리하였다.
Oil Red O 염색
Hepa1c1c cell은 10% formalin(Sigma-Aldrich Co.)으
로 10분간 고정한 다음 60% isopropanol로 세척하였다. 그 리고 60% Oil Red O solution(Sigma-Aldrich Co.)을 첨가 하여 실온에서 1시간 동안 염색시킨 후, 광학현미경(Olym- pus CK2, Olympus Optical Co., Tokyo, Japan)을 이용하 여 염색 정도를 관찰하였다.
중성지방 함량 측정
세포 내 축적된 중성지방 함량은 아산셋트 중성지방 측정 용 kit(Asan Pharm. Co., Seoul, Korea)을 사용하여 분석하 였다. 단백질 정량(Pierce BCA protein assay kit, Thermo Scientific, Rockford, IL, USA)을 통해 시료 내 단백질 함량 을 측정하였으며, 결과값은 시료 내 중성지방 함량을 단백질 함량으로 나누어 표시하였다.
Western blot 분석
Hepa1c1c cell을 lysis buffer(150 mM NaCl, 50 mM Tris-HCl, 1 mM EDTA, 50 mM NaF, 10 mM Na4P2O7, 1% IGEPAL, 2 mM Na3VO4, 0.25% protease inhibitor cocktail, and 1% phosphatase inhibitor cocktail)로 용해 시킨 후 단백질 정량을 시행하였다. 10 μg의 단백질을 11%
SDS-PAGE에서 전기영동 한 다음 nitrocellulose mem- brane에 2시간 동안 전이시켰다. Membrane은 1시간 동안 blocking 한 후 fibroblast growth factor 21(FGF21; 1:
1,000, ab171941; Abcam, Cambridge, MA, USA), β-actin (1:5,000, A5441; Sigma-Aldrich Co.)에 대한 항체로 검 출하였다.
유전자 발현 측정
Hepa1c1c cell은 TRI reagent(Life Technologies) 용 액을 이용하여 total RNA를 분리하였다. 분리된 total RNA 를 정량한 후 제조회사의 방법에 준하여 역전사 반응은 oli- go dT primer와 AMV reverscriptase를 이용하여 0.5 μg 의 RNA에서 cDNA를 합성하였다. 이 cDNA를 template로 사용하여 각각의 primer와 TaKaRa SYBR premix Ex Taq kit(TaKaRa Bio Inc., Mountain View, CA, USA)을 첨가하 고 TaKaRa Real time PCR 기기(TaKaRa Bio Inc., Otsu, Japan)를 이용하여 증폭시킨 후 정량 소프트웨어를 사용하 여 분석하였다. PCR 조건은 95°C에서 10초 반응 후, 95°C 에서 5초와 60°C에서 30초를 40회 반복하는 cycle로 수행 하였다. Real time PCR로 증폭된 산물은 comparative cy- cle threshold(Ct)법을 이용하여 정량하였으며, 각 시료는 18s의 발현량으로 보정하였다. PCR에 사용한 각 유전자의 primer는 Table 1에 제시하였다.
통계처리
본 실험 결과는 Prism 5 software(GraphPad, San Diego, CA, USA)를 이용하여 평균과 오차를 나타내었고, 일원배치 분산분석(one-way ANOVA test)을 실시한 후 Dunnett
Table 1. Mouse primers used in qRT-PCR analyses
Gene Forward primer (5’→3’) Reverse primer (5’→3’)
PPARα ACOX1 CPT-1 SREBP-1
ACCα FAS FGF21
18s
ACGCTCCCGACCCATCTTTAG AAGATGGATCCTAAGCCAGCTGAA
TGGCTTCAGAGCCAGTGGAG GGAGACATCGCAAACAAGCTGA
GAAGTCAGAGCCACGGCACA AGCACTGCCTTCGGTTCAGTC ACACTGAAGCCCACCTGGAGA TTCCGATAACGAACGAGACTCT
TCCATAAATCGGCACCAGGAA CAGCTTACCACAAAGCCAGCTACTC
AGCGATGGTGGCTGTCATTC CAGACTGCAGGCCAGATCCA GGCAATCTCAGTTCAAGCCAGTC
AAGAGCTGTGGAGGCCACTTG CTGCAGGCCTCAGGATCAAAG TGGCTGAACGCCACTTGTC
A B
Fig. 1. Effect of isorhamnetin on lipid accumulation in hepatocytes. Hepa1c1c cells exposed to FFAs (1 mM) were treated with
or without isorhamnetin (10∼25 μM) for 24 h. (A) Oil Red O staining of lipid droplets photographed at 200× magnification.(B) Intracellular triglyceride content in hepa1c1c cells. Data are presented as the mean±SEM. ***
P<0.001 compared with FFA treatment
by Dunnett multiple range test. FFA, free fatty acid; Isor, isorhamnetin; TG, triglycerides.multiple range test로
P
<0.05 수준에서 유의차를 검증하 였다.결 과
간세포에서 유리지방산으로 유도된 지질 축적에 대한 iso- rhamnetin의 저해 효과
간세포(Hepa1c1c cell)에 유리지방산(1 mM, 2:1 ratio of oleate/palmitate)과 함께 isorhamnetin을 처리하여 세 포 내 지질 축적의 변화를 관찰하였다. 세포 내 지질의 축적 정도는 지질성분에 특이적으로 반응하는 Oil Red O 염색을 통해서 측정하였고, 대조군에 비해 유리지방산 처리군에서 지질 축적이 현저히 증가함을 확인하였다. 이때 isorham- netin 처리군에서는 유리지방산으로 유도된 지질 축적이 유 의적으로 억제되었다(Fig. 1A). 이와 유사하게 간세포 내 중성지방 함량은 유리지방산 처리군에 비해 isorhamnetin 처리군에서 농도 의존적으로 감소하였다(Fig. 1B).
Isorhamnetin에 의한 지질대사 관련 유전자 발현의 변화
Isorhamnetin의 지질 축적 저해 효과에 대한 기전을 알아 보기 위해 세포 내 지방산 합성 및 산화와 관련된 유전자 발현을 조사하였다. PPARα는 간 내 지방산 산화에 관여하 는 핵전사인자이다. PPARα 표적 유전자의 대부분은 지방산산화를 조절하는 중요한 효소이므로 간 내 지질 축적에 영향 을 미칠 수 있다. PPARα 유전자 발현 수준은 isorhamnetin 처리군에서 농도 의존적으로 증가하였다(Fig. 2A). 지방산 산화를 촉진하는 PPARα 표적 유전자인 ACOX1과 CPT-1 발현 수준 역시 isorhamnetin 처리군에서 현저히 증가하였 다(Fig. 2A).
한편 지질 합성 전사인자인 SREBP-1 유전자 발현 수준 은 유리지방산 처리군에 비해 isorhamnetin 처리군에서 현 저히 저해되었다(Fig. 2B). SREBP-1에 의해 활성화되는 지질 합성 유전자인 ACC와 FAS의 유전자 발현 수준은 유 리지방산 처리군에서 증가하였고, 이러한 증가는 isorham- netin 처리군에서 농도 의존적으로 감소하였다(Fig. 2B).
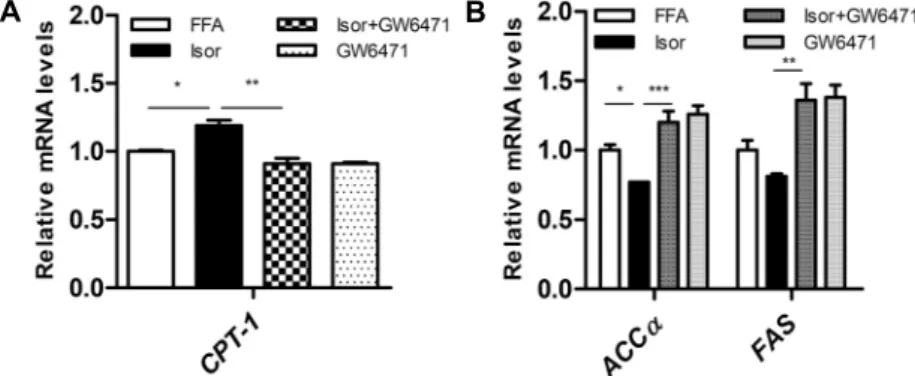
간세포에서 isorhamnetin 작용에 대한 PPARα저해제의 효과
Isorhamnetin에 의한 대사적 변화가 지질대사를 조절하 는 전사인자인 PPARα와 관련이 있는지를 조사하기 위해 간세포에 isorhamnetin과 함께 PPARα 저해제인 GW6471 을 처리하였다. 그 결과 isorhamnetin 처리군에서 증가한 지방산 산화 관련 유전자인 CPT-1 발현은 isorhamnetin+
GW6471 처리군에서 부분적으로 감소하였다(Fig. 3A). 또 한, isorhamnetin 처리군에서 감소한 지방산 합성 관련 유 전자 ACCα와 FAS의 발현은 isorhamnetin+GW6471 처리
A B
Fig. 2. Effect of isorhamnetin on expression of gene involved in lipid metabolism in hepatocytes. Hepa1c1c cells exposed to FFAs
(1 mM) were treated with or without isorhamnetin (10∼25 μM) for 24 h. (A) Relative mRNA expression levels of PPARα, ACOX1, and CPT-1 in hepa1c1c cells. (B) Relative mRNA expression levels of SREBP-1, ACCα, and FAS in hepa1c1c cells. Data are presented as the mean±SEM. *P<0.05 compared with FFA treatment by Dunnett multiple range test. FFA, free fatty acid; Isor,
isorhamnetin; PPARα, peroxisome proliferator-activated receptor α; ACOX1, acyl-CoA oxidase 1; CPT-1, carnitine palmitoyltransferase 1; SREBP-1, sterol regulatory element-binding protein 1; ACCα, acetyl-CoA carboxylase α; FAS, fatty acid synthase.A B
Fig. 3. Effect of PPARα antagonist on isorhamnetin action in hepatocytes. Hepa1c1c cells exposed to FFAs were treated with
or without isorhamnetin (10∼25 μM) for 24 h. GW6471 (10 μM) was added 1 h prior to the treatment with isorhamnetin and FFAs. (A) Relative mRNA expression levels of fatty acid oxidative PPARα target gene (CPT-1) in hepa1c1c cells. (B) Relative mRNA expression levels of lipogenic genes (ACCα and FAS) in hepa1c1c cells. Data are presented as the mean±SEM. *P<0.05,
**
P<0.01,
***P<0.001 compared with isorhamnetin treatment by Dunnett multiple range test. FFA, free fatty acid; Isor, isorhamnetin;
CPT-1, carnitine palmitoyltransferase 1; ACCα, acetyl-CoA carboxylase α; FAS, fatty acid synthase.
군에서 유의적으로 증가하였다(Fig. 3B).
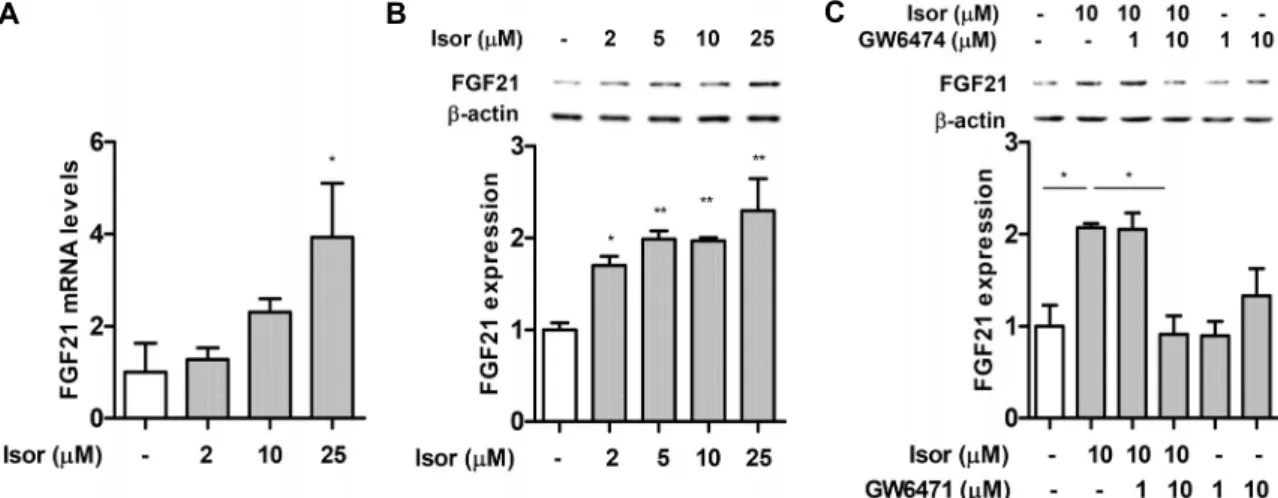
간세포에서 FGF21 발현에 대한 isorhamnetin의 효과
FGF21은 주로 간에서 분비되며(hepatokine) 당/지질의 항상성에 관여하는 대사성 호르몬이다(19,20). FGF21은 지질 합성을 저해함으로써 지방간 질환을 개선한다(21). 또 한, FGF21은 PPARα에 의해 발현이 증가한다(22). 따라서 본 연구에서는 isorhamnetin이 FGF21 발현을 증가시키는 지를 확인하였다. Isorhamnetin 처리 시 FGF21 유전자 발 현 및 단백질 발현 수준은 농도 의존적으로 증가하였다(Fig.4A, 4B). 한편 isorhamnetin 처리군에서 증가된 FGF21 발 현은 isorhamnetin+GW6471 처리군에서 감소하였다(Fig.
4C).
고 찰
지방간증은 지질대사의 불균형으로 인해 간세포 내에 과
잉의 중성지방이 축적된(중량의 5% 이상) 상태로서 비만이 매우 중요한 원인인자로 알려져 있다(23). 이는 과도한 식이 지방의 섭취 및 지방조직에서의 지방 분해 증가로 인한 간세 포 내로 유리지방산 유입 증가, 그리고 간세포 내에서 지질 합성의 증가 및 지방산 산화의 감소 등에 기인한다(3,16).
또한, 유리지방산으로 유도된 간세포 내 지방 축적은 식이조 절 및 치료를 통해 가역적으로 회복이 가능하기 때문에 지방 산 대사 조절은 초기 비알코올성 지방간질환의 예방에 중요 하다. 실제로 많은 연구들은 지방간증과 비알코올성 지방간 질환의 진행을 차단하기 위한 식품 유래 활성성분에 대해 보고하고 있다(24,25). 예를 들면 감귤류 성분인 naringe- nin은 PPARα 활성화를 유도함으로써 간세포 내 지질대사 를 개선한다(15). 또한, 채소와 과일에 존재하는 quercetin 은 간에서 PPARα 발현을 증가시킴으로써 간에서의 지방산 산화를 높여 고지방식이로 유도된 비만쥐의 간 내 지질 축적 을 감소시킨다(26). Isorhamnetin은 quercetin의 대사산물 로 알려져 있으며, 본 연구에서는 isorhamnetin이 유리지방
A B C
Fig. 4. Effect of isorhamnetin on FGF21 expression in hepatocytes. Hepa1c1c cells were incubated with various concentrations
of isorhamnetin for 16 h. (A) Relative mRNA expression levels of FGF21 were determined by qRT-PCR. (B) FGF21 protein was measured by western blotting. Data are presented as the mean±SEM. *P<0.05,
**P<0.01 compared with the control by Dunnett
multiple range test. (C) FGF21 protein in hepa1c1c cells in the presence of isorhamnetin and/or PPARα antagonist GW6471. Data are presented as the mean±SEM. *P<0.05 compared with isorhamnetin treatment by Dunnett multiple range test. Isor, isorhamnetin;
FGF21, fibroblast growth factor 21.
산으로 유도된 지방간세포에서 농도 의존적으로 중성지방 함량을 감소시킴을 확인하였다. 이러한 isorhamnetin의 효 과는 PPARα 항진제인 bezafibrate(20 μM) 및 fenofibrate (100 μM)와 유사한 수준인 점으로 보아(27,28), isorham- netin은 비알코올성 지방간 예방에 효과적인 식이성분이 될 수 있을 것으로 기대된다.
PPARα는 지질대사에서 중요한 역할을 담당하고(10,29), 비알코올성 지방간질환 개선 및 치료의 효과적인 타깃으로 간주되고 있다(30,31). 본 연구에서는 지방간을 모방한 간 세포 모델계에서 isorhamnetin이 PPARα 발현 및 활성화에 미치는 영향을 조사하였다. 그 결과 유리지방산으로 유도된 간세포에서 isorhamnetin이 PPARα 발현을 농도 의존적으 로 증가시켰다. PPARα 활성화는 지방산 수송단백질 및 long-chain acyl-CoA synthetase 발현을 유도하여 미토 콘드리아 내로의 지방산 수송을 증가시키고, ACOX1 및 CPT-1의 발현을 증가시켜 지방산 산화를 증가시킨다. 본 연구에서 isorhamnetin은 PPARα 발현뿐만 아니라 PPARα 타깃 효소인 ACOX1, CPT-1 발현을 유의적으로 증가시켰 다. 반면 PPARα 저해제(GW6417)는 isorhamnetin으로 유 도된 지방산 산화 유전자 발현의 증가를 억제하였다. 유사하 게 PPARα 결손쥐에서 지방산 산화 감소에 기인하여 심각한 지방간증이 나타났고(11), 간세포 선택적인 PPARα 결손은 지방간증을 촉진하였다(32). 또한, PPARα 항진제(fenofi- brate)는 지방산 산화를 증가시킴으로써 비알코올성 지방간 질환을 개선하였다(33). 따라서 이들 결과는 isorhamnetin 이 PPARα 활성화 및 지방산 산화 유전자 발현의 증가를 통해 유리지방산으로 유도된 간세포 내 지질 축적을 개선할 수 있음을 시사한다.
최근 PPARα에 의해 전사 조절되는 지질대사 관련 단백질 FGF21이 주목을 끌고 있다. FGF21은 주로 간에서 발현되
는 분비 단백질이며, 많은 연구 보고들을 통해서 FGF21이 당/지질대사 항상성에 있어 중요한 대사 조절자임이 밝혀졌다 (19,20). PPARα는 FGF21 gene promoter에 직접 결합하 여 전사를 유도하고 FGF21 발현을 조절한다. PPARα 항진제 (Wy 14643)는 FGF21 발현을 증가시켰고(22), PPARα 결 손쥐에서 FGF21 발현이 감소하였다(22,34). 한편 PPARα 리간드 활성을 나타내는 curcumin은 간세포에서 FGF21 발 현을 유도하며(35), 식이로 유도된 비만쥐에서 지방간 개선 과 에너지 소비를 증가시켰다(36). 본 연구에서 isorham- netin은 유전자 및 단백질 수준에서 FGF21 발현의 증가를 유도하였고, PPARα 저해제 GW6471은 isorhamnetin에 의 한 FGF21 단백질 발현 증가를 억제하였다. 이러한 결과는 isorhamnetin이 PPARα 활성화를 통해 FGF21 발현을 매개 할 가능성을 시사한다. 흥미롭게도 FGF21은 간세포에서 지 질 합성 조절자인 SREBP-1과도 관련이 있다(37). FGF21 은 유리지방산으로 유도된 간세포에서 SREBP-1의 분해와 핵 내 이동의 저해 및 타깃 유전자 발현을 감소시킴으로써 지질 합성과 중성지방 축적을 감소시킨다(37). 본 연구에서 는 isorhamnetin이 농도 의존적으로 SREBP-1과 그 down- stream 유전자 ACCα, FAS 발현을 유리지방산으로 유도된 간세포에서 감소시켰다. 또한, PPARα 저해제(GW6417)는 isorhamnetin으로 유도된 지질 합성 유전자 발현의 감소를 차단하였다. 이러한 결과들은 isorhamnetin에 의한 FGF21 발현의 증가가 SREBP-1을 감소시킴으로써 지질 합성 유전 자의 발현을 감소시킬 가능성을 시사한다. 즉 isorhamnetin 에 의해 유도된 FGF21 발현은 PPARα와 SREBP-1 간의 cross-talk에 관여하여 지방간증을 조절하는 데 기여할 것 으로 생각된다.
종합하자면 isorhamnetin은 간세포 내 지질 산화/합성계를 조절하여 지질 축적을 억제하고, 이러한 조절작용은 PPARα
경로와 연관된 것으로 생각된다. Isorhamnetin은 비만과 관 련된 비알코올성 지방간 개선과 예방을 위한 유용한 식이성 분으로 활용이 가능할 것으로 기대된다. 향후 isorhamne- tin을 비만성 지방간질환 예방/개선을 위한 기능성 식품 및 생약제제 성분으로 활용하기 위해서는 isorhamnetin의 생체 이용률을 감안한
in vivo
연구가 더 필요할 것으로 사료된다.요 약
지질대사 불균형에 따른 간세포 내 중성지방의 축적은 비만 성 지방간증뿐만 아니라 지방간염으로의 이행에 중요한 요 인으로 작용한다. 본 연구에 사용된 isorhamnetin은 채소, 과일 등에 존재하며 항산화/항염증 활성 및 비만성 대사질환 개선에 효과가 있다고 알려져 있다. 본 연구에서는 유리지방 산으로 유도된 지방간세포에서 isorhamnetin의 지질 축적 억제 효과와 그 작용기전에 대해 조사하였다. 간세포(Hepa 1c1c cell)에 유리지방산(1 mM, 2:1 ratio of oleate/pal- mitate)과 함께 isorhamnetin을 처리한 후 세포 내 지질 축 적의 정도를 확인하기 위해 Oil Red O 염색과 중성지방 함량 을 측정하였다. 또한, 지질대사와 관련된 유전자 및 단백질 발현을 qPT-PCR 또는 western blotting으로 측정하였다.
본 연구 결과 isorhamnetin은 유리지방산으로 유도된 간세 포에서 농도 의존적으로 세포 내 지질 함량을 감소시켰고, 이와 함께 간의 지방산 산화를 촉진하는 주요 인자인 PPAR α 및 그 타깃 분자(CPT-1, ACOX1) 발현 증가, 간세포 지질 합성의 주요 전사인자인 SREBP-1의 발현 억제 및 down- stream 타깃인 FAS와 ACC의 발현 감소가 관찰되었다. 나 아가 PPARα 활성 저해제인 GW6471 처리는 isorhamnetin 에 의해 유도된 지방대사 관련 유전자 발현 변화를 차단하였 다. 이러한 결과들은 isorhamnetin이 간세포 내 지질 산화/
합성계를 조절하여 지질 축적을 억제하며, 이러한 조절작용 이 PPARα 경로와 연관되어 있음을 의미하는 것으로 생각된 다. Isorhamnetin의 지질대사 조절작용은 비알코올성 지방 간질환의 예방과 치료에 유용할 것으로 기대된다.
감사의 글
이 논문은 2017년도 정부(교육부)의 재원으로 한국연구재 단의 지원을 받아 수행된 기초연구사업임(NRF-2017R1D 1A1B03033004).
REFERENCES
1. Farrell GC, Larter CZ. 2006. Nonalcoholic fatty liver dis- ease: from steatosis to cirrhosis. Hepatology 43: S99-S112.
2. de Alwis NM, Day CP. 2008. Non-alcoholic fatty liver dis- ease: the mist gradually clears. J Hepatol 48: S104-112.
3. Musso G, Gambino R, Cassader M. 2009. Recent insights into hepatic lipid metabolism in non-alcoholic fatty liver dis- ease (NAFLD). Prog Lipid Res 48: 1-26.
4. Ibrahim SH, Kohli R, Gores GJ. 2011. Mechanisms of lipo- toxicity in NAFLD and clinical implications. J Pediatr Gas-
troenterol Nutr 53: 131-140.
5. Pan MH, Lai CS, Tsai ML, Ho CT. 2014. Chemoprevention of nonalcoholic fatty liver disease by dietary natural com- pounds. Mol Nutr Food Res 58: 147-171.
6. Chavez-Tapia NC, Rosso N, Tiribelli C. 2012. Effect of intra- cellular lipid accumulation in a new model of non-alcoholic fatty liver disease. BMC Gastroenterol 12: 20.
7. Rawson RB. 2003. The SREBP pathway-insights from in- sigs and insects. Nat Rev Mol Cell Biol 4: 631-640.
8. Liang G, Yang J, Horton JD, Hammer RE, Goldstein JL, Brown MS. 2002. Diminished hepatic response to fasting/
refeeding and liver X receptor agonists in mice with selective deficiency of sterol regulatory element-binding protein-1c.
J Biol Chem 277: 9520-9528.
9. Zheng L, Lv GC, Sheng J, Yang YD. 2010. Effect of miRNA- 10b in regulating cellular steatosis level by targeting PPAR- α expression, a novel mechanism for the pathogenesis of NAFLD. J Gastroenterol Hepatol 25: 156-163.
10. Pawlak M, Lefebvre P, Staels B. 2015. Molecular mecha- nism of PPARα action and its impact on lipid metabolism, inflammation and fibrosis in non-alcoholic fatty liver disease.
J Hepatol 62: 720-733.
11. Gao Q, Jia Y, Yang G, Zhang X, Boddu PC, Petersen B, Narsingam S, Zhu YJ, Thimmapaya B, Kanwar YS, Reddy JK. 2015. PPARα-deficient ob/ob obese mice become more obese and manifest severe hepatic steatosis due to decreased fatty acid oxidation. Am J Pathol 185: 1396-1408.
12. Sahebkar A, Chew GT, Watts GF. 2014. New peroxisome proliferator-activated receptor agonists: potential treatments for atherogenic dyslipidemia and non-alcoholic fatty liver disease. Expert Opin Pharmacother 15: 493-503.
13. Repa JJ, Liang G, Ou J, Bashmakov Y, Lobaccaro JM, Shimomura I, Shan B, Brown MS, Goldstein JL, Mangels- dorf DJ. 2000. Regulation of mouse sterol regulatory ele- ment-binding protein-1c gene (SREBP-1c) by oxysterol re- ceptors, LXRalpha and LXRbeta. Genes Dev 14: 2819-2830.
14. Panchal SK, Poudyal H, Brown L. 2012. Quercetin amelio- rates cardiovascular, hepatic, and metabolic changes in diet- induced metabolic syndrome in rats. J Nutr 142: 1026-1032.
15. Goldwasser J, Cohen PY, Yang E, Balaguer P, Yarmush ML, Nahmias Y. 2010. Transcriptional regulation of human and rat hepatic lipid metabolism by the grapefruit flavonoid nar- ingenin: role of PPARα, PPARγ and LXRα. PLoS One 5:
e12399.
16. Kim CS, Kwon Y, Choe SY, Hong SM, Yoo H, Goto T, Kawada T, Choi HS, Joe Y, Chung HT, Yu R. 2015. Quer- cetin reduces obesity-induced hepatosteatosis by enhancing mitochondrial oxidative metabolism via heme oxygenase-1.
Nutr Metab (Lond) 12: 33.
17. Zhang Y, Gu M, Cai W, Yu L, Feng L, Zhang L, Zang Q, Wang Y, Wang D, Chen H, Tong Q, Ji G, Huang C. 2016.
Dietary component isorhamnetin is a PPARγ antagonist and ameliorates metabolic disorders induced by diet or leptin deficiency. Sci Rep 6: 19288.
18. Igarashi K, Ohmuma M. 1995. Effects of isorhamnetin, rham- netin, and quercetin on the concentrations of cholesterol and lipoperoxide in the serum and liver and on the blood and liver antioxidative enzyme activities of rats. Biosci Biotech-
nol Biochem 59: 595-601.
19. Itoh N, Ornitz DM. 2011. Fibroblast growth factors: from molecular evolution to roles in development, metabolism and disease. J Biochem 149: 121-130.
20. Kharitonenkov A, Shiyanova TL, Koester A, Ford AM, Mi- canovic R, Galbreath EJ, Sandusky GE, Hammond LJ, Moyers JS, Owens RA, Gromada J, Brozinick JT, Hawkins ED, Wroblewski VJ, Li DS, Mehrbod F, Jaskunas SR, Shanafelt AB. 2005. FGF-21 as a novel metabolic regulator.
J Clin Invest 115: 1627-1635.
21. Gomez-Samano MA, Grajales-Gomez M, Zuarth-Vazquez JM, Navarro-Flores MF, Martinez-Saavedra M, Juarez-Leon OA, Morales-Garcia MG, Enriquez-Estrada VM, Gomez- Perez FJ, Cuevas-Ramos D. 2017. Fibroblast growth factor 21 and its novel association with oxidative stress. Redox
Biol 11: 335-341.
22. Lundasen T, Hunt MC, Nilsson LM, Sanyal S, Angelin B, Alexson SE, Rudling M. 2007. PPARα is a key regulator of hepatic FGF21. Biochem Biophys Res Commun 360: 437- 440.
23. Koppe SW. 2014. Obesity and the liver: nonalcoholic fatty liver disease. Transl Res 164: 312-322.
24. Ronis MJ, Chen Y, Badeaux J, Badger TM. 2009. Dietary soy protein isolate attenuates metabolic syndrome in rats via effects on PPAR, LXR, and SREBP signaling. J Nutr 139:
1431-1438.
25. Ali F, Ismail A, Kersten S. 2014. Molecular mechanisms underlying the potential antiobesity-related diseases effect of cocoa polyphenols. Mol Nutr Food Res 58: 33-48.
26. Kobori M, Masumoto S, Akimoto Y, Oike H. 2011. Chronic dietary intake of quercetin alleviates hepatic fat accumu- lation associated with consumption of a Western-style diet in C57/BL6J mice. Mol Nutr Food Res 55: 530-540.
27. Ding L, Liu JL, Hassan W, Wang LL, Yan FR, Shang J.
2014. Lipid modulatory activities of Cichorium glandulosum
Boiss et Huet are mediated by multiple components within
hepatocytes. Sci Rep 4: 4715.28. Ma L, Li K, Wei D, Xiao H, Niu H, Huang W. 2015. High anti-oxidative and lipid-lowering activities of flavonoid gly- cosides-rich extract from the leaves of Zanthoxylum bungea-
num in multi-system. J Food Nutr Res 3: 62-68.
29. Lu Y, Liu X, Jiao Y, Xiong X, Wang E, Wang X, Zhang Z, Zhang H, Pan L, Guan Y, Cai D, Ning G, Li X. 2014.
Periostin promotes liver steatosis and hypertriglyceridemia through downregulation of PPARα. J Clin Invest 124: 3501- 3513.
30. Bechmann LP, Vetter D, Ishida J, Hannivoort RA, Lang UE, Kocabayoglu P, Fiel MI, Muñoz U, Patman GL, Ge F, Yakar S, Li X, Agius L, Lee YM, Zhang W, Hui KY, Televantou D, Schwartz GJ, LeRoith D, Berk PD, Nagai R, Suzuki T, Reeves HL, Friedman SL. 2013. Post-transcriptional activa- tion of PPAR alpha by KLF6 in hepatic steatosis. J Hepatol 58: 1000-1006.
31. Abdelmegeed MA, Yoo SH, Henderson LE, Gonzalez FJ, Woodcroft KJ, Song BJ. 2011. PPARα expression protects male mice from high fat-induced nonalcoholic fatty liver.
J Nutr 141: 603-610.
32. Montagner A, Polizzi A, Fouche E, Ducheix S, Lippi Y, Lasserre F, Barquissau V, Regnier M, Lukowicz C, Ben- hamed F, Iroz A, Bertrand-Michel J, Al Saati T, Cano P, Mselli-Lakhal L, Mithieux G, Rajas F, Lagarrigue S, Pineau T, Loiseau N, Postic C, Langin D, Wahli W, Guillou H.
2016. Liver PPARα is crucial for whole-body fatty acid ho- meostasis and is protective against NAFLD. Gut 65: 1202- 1214.
33. Seo YS, Kim JH, Jo NY, Choi KM, Baik SH, Park JJ, Kim JS, Byun KS, Bak YT, Lee CH, Kim A, Yeon JE. 2008.
PPAR agonists treatment is effective in a nonalcoholic fatty liver disease animal model by modulating fatty-acid meta- bolic enzymes. J Gastroenterol Hepatol 23: 102-109.
34. Oishi K, Uchida D, Ishida N. 2008. Circadian expression of FGF21 is induced by PPARα activation in the mouse liver. FEBS Lett 582: 3639-3642.
35. Zeng K, Tian L, Patel R, Shao W, Song Z, Liu L, Manuel J, Ma X, McGilvray I, Cummins CL, Weng J, Jin T. 2017.
Diet polyphenol curcumin stimulates hepatic Fgf21 produc- tion and restores its sensitivity in high-fat-diet-fed male mice.
Endocrinology 158: 277-292.
36. Xu J, Lloyd DJ, Hale C, Stanislaus S, Chen M, Sivits G, Vonderfecht S, Hecht R, Li YS, Lindberg RA, Chen JL, Jung DY, Zhang Z, Ko HJ, Kim JK, Véniant MM. 2009.
Fibroblast growth factor 21 reverses hepatic steatosis, in- creases energy expenditure, and improves insulin sensitivity in diet-induced obese mice. Diabetes 58: 250-259.
37. Zhang Y, Lei T, Huang JF, Wang SB, Zhou LL, Yang ZQ, Chen XD. 2011. The link between fibroblast growth factor 21 and sterol regulatory element binding protein 1c during lipogenesis in hepatocytes. Mol Cell Endocrinol 342: 41-47.