Res. Plant Dis. 18(2) : 101−108 (2012) © The Korean Society of Plant Pathology http://dx.doi.org/10.5423/RPD.2012.18.2.101
자연감염된 소나무와 잣나무 내 소나무재선충 분포 및 인공접종한 소나무 묘목 내에서의 소나무재선충 이동
김재근·김병관1·이승규2·김진철3·한상섭4·차병진5*
나무종합병원, 1동강나무병원, 2국립산림과학원 산림병해충연구과, 3한국화학연구원 친환경신물질연구그룹,
4전북대학교 산림환경과학과, 5충북대학교 식물의학과
Distribution of Bursaphelenchus xylophilus in Naturally Infected Pinus densiflora and P. koraiensis and Migration of B. xylophilus in Artificially Inoculated
P. densiflora Seedlings
Jaegeun Kim, Byungkwan Kim1, Seung-Kyu Lee2, Jin-Cheol Kim3, Sangsub Han4 and Byeongjin Cha5* Kang's Tree Hospital, Seoul 138-050, Korea
1Dong-Gang Tree Hospital, Youngwol 230-800, Korea
2Division of Forest Insect Pests and Diseases, Korea Forest Research Institute, Seoul 130-712, Korea
3Ecofriendly New Materials Research Group, Daejeon 305-600, Korea
4Department of Forest Environmental Science, Chonbuk National University, Jeonju 561-756, Korea
5Department of Plant Medicine, Chungbuk National University, Cheongju 361-763, Korea (Received on May 21, 2012; Revised on June 14, 2012; Accepted on June 19, 2012)
In 2006, pinewood nematode, Bursaphelenchus xylophilus, was isolated from about 50 years old trees of Pinus densiflora and P. koraiensis showing leaf-wilt and -drying symptoms in Gwangju, Gyeonggi-do and Chuncheon, Gangwon-do. Isolation of pinewood nematodes from sapwood of infected pine trees showed no difference in population density between tree species and among the sampling heights on the main stem.
Migration of pinewood nematodes in the host tree were investigated by inoculation of red pine (P. densiflora, 3 years old) seedlings with B. xylophilus. The nematodes seemed to move in red pine seedlings prior to multiplication and it might have taken about 20 days to start multiplication and expression of symptoms including wilt and dieback. In initial time after inoculation, nematodes started migration through the cortical resin canal from inoculated site and further showed upward and downward movements. More nematodes were observed in cortical resin canal during early period of inoculation and later in resin canal of xylem and tracheid also while, the pith still remained free from nematode. The density of B. xylophilus was higher in seedlings of low-vigor with poor root growth than in seedlings of normal root growth. Seedlings showing high density of B. xylophilus exhibited stem discoloration and secondary infection by fungus at the inoculation site.
Keywords : Pinewood nematodes, Red pine, Resin canal, Wilt
서 론
소나무 시들음병 증상은 1913년 일본에서 처음으로 보 고되었다. 그러나, 당시에는 증상만이 보고되었을 뿐, 그
원인이 소나무재선충이라고 밝혀진 것은 1975년이다(Futai, 2008). 소나무 시들음병의 원인이 소나무재선충이라는 사 실은 1931년 미국에서 처음으로 보고된 이래 침엽수 27 종과 비침엽수 7종에서 발병이 확인되었다(Agrios, 2005).
이 병은 현재 일본을 비롯하여 우리나라와 중국을 포함 한 동아시아 지역의 소나무림에 막대한 피해를 주고 있 으며, 포르투갈에서도 발생이 보고되었다(Mota 등, 1999).
우리나라에서는 1988년 부산 금정산에서 처음으로 발견
*Corresponding author
Phone)+82-43-261-2557, Fax) +82-46-271-4414 Email) [email protected]
Research Article Open Access
된 이래, 18년이 지난 2006년에는 전국 8개도 54개 시군 구에서 7,871 ha의 소나무 임지에 발생하였으며, 최근에 는 충남 보령에서도 발생하였다. 2006년 12월에는 자연 상태에서 북방솔수염하늘소(Monochamus saltuarius)에 의 해 잣나무(Pinus koraiensis)에도 옮겨지는 것이 처음으로 확인되었다(http://news. naver.com).
지금까지 알려진 소나무 시들음병의 발병기작은 침입 한 소나무재선충이 소나무의 물관과 수지도에 기생하면 서 수분 이동을 방해하기 때문이라는 것이 가장 일반적 인 내용이다(Agrios, 2005). 하지만, 이러한 기작에 의하 여 소나무가 완전히 말라죽는다는 데 대해서는 이견이 많 아, 소나무재선충 감염에 따른 소나무의 광합성률 감소, 물 관부와 코르크유조직의 변성, traumatic 수지도 형성, 형 성층 파괴, 식물독소물질 생산, 호흡저해, 에틸렌 형성, 잎 의 증산량 변화에 따른 수분포텐셜 변화 등 다양한 발병 기작에 대한 규명이 시도되었다. 그럼에도 불구하고 아직 도 소나무 고사의 주된 원인으로 확실하게 밝혀진 발병 기작은 없다. 이 중에서도 가장 설득력 있는 가설은 잎의 증산량 및 줄기내부의 수분포텐셜과 관련된 여러 요인들 이 복합적으로 나타난다는 것이다(Fukuda, 1997). 최근에 는 소나무재선충 감염목에서 함께 분리되는 다양한 미생 물이 소나무 시들음병의 원인으로 의심되고 있으며, 이에 대한 연구가 활발하게 이루어지고 있다. 일본에서는 소나 무재선충으로부터 Bacillus 속 세균을 분리하였고, 소나무 재선충의 표면을 무균처리 하면 소나무재선충이 병원성 을 잃는다는 보고가 있다(Kawazu 등, 1997). 한편으로는 세균이 생산해 내는 물질들에 의해 시들음 증상이 유발 된다는 보고도 있다(Oku 등, 1979). 중국의 Zhao 등(2003) 은 소나무재선충으로부터 Pseudomonas속과 Pantoea속, 그리고 Enterobacter속 등의 세균을 분리하고, 이들 세균 이 생성한 물질을 소나무 callus에 접종하였을 때 갈변증 상을 일으킴을 확인함으로써 이들 물질이 식물독성이 있 음을 확인하였고, 세균을 제거한 무균선충의 경우 일반 소나무재선충에 비해 병원성이 약하거나 없다고 하였다.
또한 Zhao 등(2005)은 일본에서 분리한 세균과 중국에서 분리한 세균의 차이를 비교하면서, 지역에 따라 분리되는 세균에 차이가 있을 수 있음을 시사했다.
그동안 국내에서는 1988년에 소나무재선충이 처음으로 확인된 이래 많은 연구가 활발하게 이루어져 왔으나, 우 리보다 앞서 시작한 다른 나라의 연구에 비하면 결과가 많이 부족한 실정이다. 특히, 대부분의 연구가 전염경로 등 소나무재선충의 생태와 소나무 시들음병의 방제에 치 중되고 있으며, 발병기작이나 선충과 미생물과의 관계 등 기본적인 연구는 미미한 실정이다. 따라서 본 연구에서는
소나무재선충의 자연감염기주로 새롭게 확인된 잣나무와 소나무로부터 소나무재선충을 분리하고 배양한 뒤, 소나 무 묘목에 접종하고, 감염목내에서 선충의 분포 및 밀도 변화와 감염조직의 관찰을 통해 기주식물 내에서의 소나 무재선충의 이동 및 증식경향을 밝히고자 하였다.
재료 및 방법
감염목으로부터 소나무재선충 분리. 외견상 피해정도 에 따른 소나무재선충의 소나무 조직 내 분포양상과 밀 도를 조사하기 위해 2006년에 국립산림과학원 남부산림 연구소로부터 경기도 광주에서 발생한 잣나무 피해목과 강원도 춘천의 소나무 피해목의 부위별 시편을 분양받았 다. 피해목들은 소나무재선충에 감염되어 말라죽기 직전 인 것들이었으며, 각 피해목 마다 지표면으로부터 50 cm 간격으로 지상 3 m까지 주간의 시편(trunk disk)을 채취하 였다. 각 주간시편은 다시 수(pith), 심재(heartwood), 변재 (sapwood) 등 3 부위로 나누어 조사하였다. 조직 내의 선 충 밀도를 조사하기 위하여 시편별로 각각의 나무 조직 으로부터 일정한 부피(1 cm3)의 시료를 3 반복씩 채취하 여 베르만깔때기법(Baerman funnel method)으로 선충을 분리한 뒤(Togashi 등, 2003), 실체현미경(40×)으로 관찰 하며 선충의 수를 세었다.
소나무재선충 배양 및 접종. 국립산림과학원으로부터 무포자 Botrytis cinerea를 분양받아 감자한천배지에 7일 이상 배양한 다음, 자연감염된 소나무로부터 분리한 소나 무재선충의 성충과 유충을 접종하고 25oC 암조건에서 배 양하였다. 소나무재선충은 Botrytis 균총의 상태에 따라 10−15일 마다 새로운 Botrytis cinerea 배지로 옮겨 계대 배양하며 증식시켰고, 필요할 때마다 멸균수와 붓으로 수 확하여 사용하였다.
소나무재선충을 접종할 소나무(Pinus densiflora)는 3년 생 묘목으로서, 지름 20 cm의 플라스틱 폿트에 재배하였 다. 접종은 묘목의 주간에 하였는데, 지제부로부터 4 cm 정도 떨어진 부위에 화염소독한 메스로 길이방향으로 1 cm 정도의 상처를 내고 멸균한 솜으로 두른 다음 파라필름 또는 알루미늄 포일로 깔때기 모양으로 약간 느슨하게 감 쌌다. 여기에 일정한 밀도(104 nematodes/ml)의 소나무재 선충 성충과 유충 혼합액 400 µl를 적셔주었다. 대조구로 는 같은 양의 멸균수를 접종하였다. 접종한 묘목은 소나 무재선충의 외부 유출을 막기 위하여 실내에 있는, 완전 히 밀폐된 식물생장실(25oC, RH 70%)에 보관하며 외관 상 변화를 관찰하였다. 시험이 끝난 소나무는 모두 소각 하였으며, 토양은 폿트와 함께 고압멸균한 다음 폐기하였다.
소나무 묘목 내 소나무재선충 분포 및 밀도변화 조사.
접종 후 소나무재선충에 감염된 조직 및 감염상태 등을 관찰하기 위하여 Ichihara 등(2000)의 방법을 변형하여 감염조직을 관찰하였다. 소나무재선충 증식 전으로 판단 되는 접종 4일 후, 접종부위를 기준으로 묘목의 주간을 1 cm 길이의 토막들로 자르고 각각의 토막들을 2.5%
glutaraldehyde에 고정하였다. 고정된 시료는 0.1 M phosphate buffer에 2−3회 세척한 후에 OCT compound(Tissuetek, New York, NY. USA)로 시료를 포매하고 냉동박편기 (Cryotome FE; Shandon, London, England)를 이용하여 두 께 60 µm의 박편을 만들었다. 각 토막으로부터 3개씩의 박편을 얻었으며, 각 박편은 광학현미경(BX50; Olympus, Tokyo, Japan) 하에서 피층수지도, 피층, 형성층, 목질부 수지도, 가도관, 수 부위 등으로 조직을 구분하여 소나무 재선충의 유무를 확인하였다.
또한, 소나무 내에서의 부위별 소나무재선충 밀도를 조 사하고 이동경로를 추적하기 위하여 소나무재선충을 접 종한 소나무 묘목을 접종 후 3일, 5일, 7일, 10일, 20일, 30일에 각각 3그루씩 무작위로 선발하여 선충 분리를 시 도하였다. 지상부의 주간은 접종부위를 기준으로 1 cm 씩 나누어 선충을 조사하였고, 가지는 위치에 따라 크게 상, 중, 하 세 부위로 나누어 각각 조사하였다. 지하부는 주 근의 경우 주간과 마찬가지로 1 cm 간격으로 나누어 조 사하였으며, 이차근 이상부터는 굵은 뿌리 위주로 묘목 1개체 당 3 뿌리씩을 조사하였다. 접종한 묘목에서 채취 한 모든 시편들은 변형한 베르만깔때기법을 이용하여 소 나무재선충을 분리하였고(Togashi 등, 2003), 실체현미경 (40×)으로 관찰하며 시편 당 소나무재선충의 수를 세었다.
선충을 분리하기 위하여 시편을 채취하는 소나무 묘목 은 모두 잎의 변색과 시들음 등 외부병징을 우선적으로 관찰하였으며, 줄기 단면의 변색여부를 확인하여 변색이 있을 경우에는 절편을 만들어 일반광학현미경 하에서 균 사의 존재여부를 확인하고 균분리를 시도하였다.
결 과
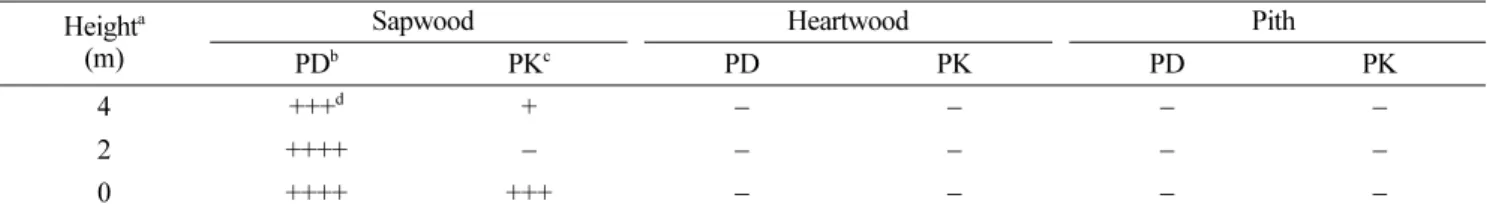
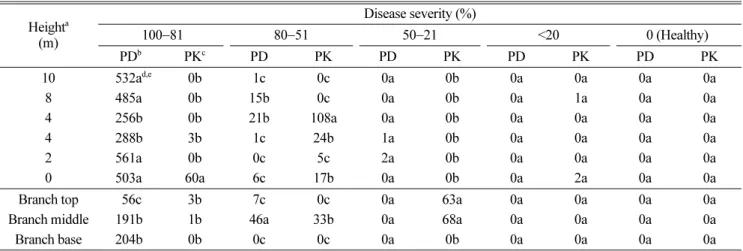
소나무재선충의 자연감염목 내 분포 및 발병정도에 따 른 분리율. 소나무재선충에 감염된 병징을 보이고 있는 경기도 광주의 잣나무들과 강원도 춘천의 소나무들을 피 해정도에 따라 구분하고, 부위별로 선충을 분리한 결과 수간의 여러 조직들 중 오직 변재부위에서만 소나무재선 충이 분리되었으며(Table 1), 피해정도가 클수록 선충 분 리율도 높아지는 경향을 보였다(Table 2). 우선 소나무와 잣나무 모두 변재를 제외한 피층과 심재 및 수에서는 소 나무재선충이 전혀 분리되지 않았다(Table 1). 발병정도에 따라서는, 수관이 완전히 말라죽은 100% 감염목의 경우 경기도 광주지역의 잣나무 시료에선 주로 지제부 쪽과 윗 부분의 가지에서 소나무재선충의 분리율이 높았으나 통 계적으로는 위치에 따른 유의차는 없었다. 반면, 강원도 춘천지역의 소나무 시료에서는 비교적 많은 선충이 분리 되었는데, 주간의 위쪽보다는 아래쪽에서 더 많이 분리되 었다. 전반적으로는 잣나무 고사목보다 소나무 고사목 시 료에서의 분리율이 더 높은 것으로 나타났다. 70% 고사 목 시료나 30−50% 고사목 시료의 경우 경기도 잣나무 시료가 강원도 소나무 시료에 비해 더 높은 선충 분리율 을 보이고 있어(Table 2), 전반적으로 지역 간 또는 수종 간 차이를 인정하기는 힘들었다. 또한, 초기 병징을 보이 는 시료의 경우에는 두 지역(수종) 모두 소나무재선충 분 리율이 매우 낮아 감염목 내 소나무재선충 밀도와 고사 율간에 밀접한 관련을 보였다.
소나무재선충의 감염초기 소나무 묘목 내 분포. 접종 4일 후 접종부위와 그 위쪽으로 1 cm 떨어진 주간시료에 서는 피층수지도와 피층조직, 형성층 부근까지 소나무재 선충의 침입이 확인되었으나(Fig. 1A, C, D), 목질부의 수 지도나 가도관 및 수에서는 전혀 관찰되지 않았다(Table 3). 피층수지도에서는 그 보다도 더 위쪽에서도 소나무재 선충이 관찰되었는데, 접종부위로부터 위쪽으로 최고 9 cm
Table 1. Isolation density of Bursaphelenchus xylophilus from different trunk tissues of naturally infected Pinus densiflora and P.
koraiensis Heighta
(m)
Sapwood Heartwood Pith
PDb PKc PD PK PD PK
4 +++d + − − − −
2 ++++ − − − − −
0 ++++ +++ − − − −
aFrom the ground level.
bP. densiflora of Chuncheon, Gangwon-do.
cP. koraiensis of Kwangju, Kyeonggi-do.
dDensity of B. xylophilus in 1 cm3 of the tissue (−: none, +: 1−10, +++: 50−100, ++++: over 100 nematodes).
까지 확인되었다(Fig. 1B). 아래쪽으로는 3 cm 떨어진 절 편의 피층수지도에서 소나무재선충이 확인되었다. 접종부 위와 그 위쪽으로 1 cm 떨어진 부위를 제외한 다른 부위 에서는 피층수지도에서만 선충의 존재가 확인되었다. 접 종 30일 후에 접종묘목을 잘라 조직 내 선충을 다시 관 찰한 결과, 피층수지도나 피층조직뿐만 아니라 목질부의 수지도와 가도관에서도 소나무재선충의 존재를 확인 할
수 있었다(Table 4). 그러나 접종 30일 후에도 수에서는 소나무재선충이 여전히 발견되지 않았다.
감염목내 소나무재선충 밀도변화. 접종 3일 후에 소 나무재선충을 분리한 결과, 묘목 내로 침입한 선충들은 대부분 접종부위에 존재하여 접종부위의 선충 밀도가 가 장 높았다. 일부는 다른 부분으로 이동하였음이 확인되었 는데, 접종부위로부터 위쪽으로는 최대 8 cm, 아래쪽으로 Table 2. Population of Bursaphelenchus xylophilus in trunks and branches of naturally infected Pinus densiflora and P. koraiensis
Heighta (m)
Disease severity (%)
100−81 80−51 50−21 <20 0 (Healthy)
PDb PKc PD PK PD PK PD PK PD PK
10 532ad,e 0b 1c 0c 0a 0b 0a 0a 0a 0a
8 485a 0b 15b 0c 0a 0b 0a 1a 0a 0a
4 256b 0b 21b 108a 0a 0b 0a 0a 0a 0a
4 288b 3b 1c 24b 1a 0b 0a 0a 0a 0a
2 561a 0b 0c 5c 2a 0b 0a 0a 0a 0a
0 503a 60a 6c 17b 0a 0b 0a 2a 0a 0a
Branch top 56c 3b 7c 0c 0a 63a 0a 0a 0a 0a
Branch middle 191b 1b 46a 33b 0a 68a 0a 0a 0a 0a
Branch base 204b 0b 0c 0c 0a 0b 0a 0a 0a 0a
aFrom the ground level.
bP. densiflora of Chuncheon, Gangwon-do.
cP. koraiensis of Kwangju, Kyeonggi-do.
dNumbers of B. xylophilus isolated from 1 cm3 of the tissue.
eNumbers followed by the same letters in each column are not significantly different in Duncan’s multiple range test (P < 0.05).
Fig. 1. Bursaphelenchus xylophilus (arrows) in cortex (A), cortical resin canal (B), xylem resin canal (C), and tracheid (D) of main stems of Pinus densiflora seedlings 4 days after artificial inoculation. (A, C, and D = 1 cm above the inoculation point, B = 9 cm above the inoculation point and stained with acid fuchsin, Scale bar = 200 µm).
Table 3. Distribution of Bursaphelenchus xylophilus in tissues of Pinus densiflora seedlings four days after inoculation
Heighta (cm) Cortical resin canal Cortical tissue Cambial region Resin canal of xylem Trachied Pith
+10 −b − − − − −
+9 + − − − − −
+8 − − − − − −
+7 − − − − − −
+6 + − − − − −
+5 + − − − − −
+4 − − − − − −
+3 + − − − − −
+2 + − − − − −
+1 + + + − − −
0* + + + − − −
−1 + − − − − −
−2 − − − − − −
−3 + − − − − −
−4 − − − − − −
a‘0’, ‘+’, and ‘−’ represent the inoculation site, above the site, and below the site, respectively.
b‘+’ and ‘−’ represent ‘detected’ and ‘not detected’, respectively.
는 4 cm 정도에서까지만 소나무재선충이 검출되었다. 접 종 5일 후에는 접종부위로부터 20 cm 이상 떨어진 신초 부위에서도 소나무재선충이 분리되었고, 지하부에서는 주 근의 말단부위까지도 분포함을 확인하였으나 2차근에서 는 전혀 분리되지 않았다. 하지만, 접종부위로부터 멀리 떨어진 조직에서 분리되는 선충은 1−2마리 정도로 밀도 가 매우 낮아 아직까지 활발하게 증식하지는 않는 것으 로 보였다. 반면에 접종 7일 후부터는 각각의 절편으로부 터 분리되는 소나무재선충의 밀도가 서서히 증가하는 것 을 관찰하였고, 비록 밀도는 낮았지만 접종한 묘목의 전 부위에서 소나무재선충을 분리할 수 있었다. 접종 10일 후에는 2차근에서도 선충이 분리되었으나(Fig. 2), 이 때 까지도 조직 내 소나무재선충의 밀도는 그리 높지 않았 으며, 외견상으로도 소나무재선충에 의한 침엽의 갈변증 상이나 처짐 증상은 나타나지 않았다. 그러나 접종 후 20 일이 경과하자 접종 묘목의 모든 부위에서 소나무재선충 의 밀도가 급격히 증가하였으며, 2년생 침엽의 갈변증상 과 함께 잎의 처짐이 관찰되기 시작하였다(Fig. 3). 그 이
후로 접종 묘목은 빠른 속도로 시들었으며, 30일이 경과 하자 소나무재선충의 밀도는 더욱 높게 증가하였고, 접종 한 소나무 묘목 모두가 빠르게 고사하였다.
한편, 접종 20일 후 묘목으로부터 소나무재선충을 분리 하였을 때, 소나무재선충의 밀도는 반복 간에 큰 차이를 나타내었다(Fig. 4). 접종한 묘목들의 지하부를 조사한 결 과, 총 세 그루 중 한 그루는 잔뿌리가 거의 발달하지 않 았고 뿌리의 생육상태도 좋지 못하였으며, 또 다른 한그 루는 뿌리의 생육상태는 양호한 편이었으나 목질부 변색 증상이 있었으며 가도관 내에서 짙은 갈색의 균사가 관 찰되었다. 그리고, 나머지 한 그루는 목질부 변색도 없었 으며 뿌리도 잘 발달되는 등 정상적인 모습을 하고 있었 다. 이들 중 뿌리가 부실한 묘목과 목질부가 변색된 묘목 에서는 소나무재선충이 높은 밀도로 분리되었던 반면, 건 전해 보이는 나머지 한 그루에서는 상대적으로 매우 적 은 수의 소나무재선충만 분리되었다. 한편, 목질부 변색 의 원인은 가도관에서 발견된 곰팡이인 Alternaria sp.였 음을 확인하였고, 그로부터 분리한 곰팡이를 기내에서 감 Table 4. Migration of inoculated Bursaphelenchus xylophilus in different tissues of Pinus densiflora seedlings
Days after inoculation
Tissue type
Cortical resin canal Cortex Cambial region Resin canal of xylem Tracheid Pith
4 +a + + − − −
30 + + + + + −
a+: detected, −: not detected.
Fig. 2. Distribution of Bursaphelenchus xylophilus in Pinus densiflora seedlings inoculated with 104 nematodes/ml/wound. Each column represents P. densiflora seedling (0: inoculation point, +: above, -: below, TW: this year's twig, MS: main stem, MR: main root, SR:
secondary roots).
Fig. 3. Wilt symptom development on Pinus densiflora seedlings inoculated with Bursaphelenchus xylophilus (104 nematodes/seedling).
자한천배지에 배양하며 균총에 소나무재선충을 접종한 결 과 소나무재선충의 증식이 확인되었다(자료 미제시).
고 찰
1988년 우리나라에서 소나무재선충에 의한 피해가 처 음 확인된 이래 해를 거듭할수록 피해규모가 증가하여 국 내 소나무림의 보호를 위해 많은 방제연구가 이루어지고 있다. 본 연구에서는 소나무재선충 접종실험을 통해 감염 목내 소나무재선충의 분포와 밀도변화를 조사하여 시들 음병의 발병진전과 어떤 관계가 있는지 알아보고자 하였다.
감염된 기주 내에서의 소나무재선충 분포 조사를 위해 소나무재선충에 자연감염되어 고사하였거나 또는 고사하 고 있는 소나무와 잣나무로부터 높이와 조직에 따라 소 나무재선충을 분리한 결과 수종이나 높이에 따른 분리율 에는 큰 차이가 없어, 소나무재선충은 감염목의 모든 부 위에 존재하고 있음을 재확인하였다. 그러나 고사목을 심 재와 변재부위로 나누어 소나무재선충을 분리 하였을 때 는 변재부위에서만 선충이 분리되었다(Table 1). 이는 양 분과 수분이동이 활발하고 유세포가 있는 변재부위가 먹 이공급원으로서 소나무재선충의 서식에 적합하기 때문으 로 보인다. 또한, 피해정도가 심할수록 소나무재선충의 분 리 밀도가 높은 것은 감염목 내 소나무재선충 밀도와 고 사율이 비례함을 보여주는 것이었다.
접종실험을 통해 감염목내 밀도조사를 실시한 결과, 접 종 후 3일까지는 대부분의 선충이 접종부위에 남아있고 일부만이 다른 부분으로 이동하는 것으로 나타났는데, 이 는 접종된 소나무재선충은 접종부에 머무는 것과 다른 부
위로 빠르게 이동하는 두 군으로 나뉜다는 Togashi 등 (2003)의 보고와 부합하는 것이었다. 또한, 접종 후 20일 까지는 소나무재선충의 분리밀도도 그리 높지 않았으며, 소나무에서 뚜렷한 증상도 없었으나, 20일 이후 소나무에 시들음병징이 나타나며 소나무재선충의 분리 밀도가 급 격히 증가하였는데, Fukuda(1997) 및 Ichihara 등(2000)을 포함한 많은 연구자들은 소나무재선충의 빠른 증식이 시 들음 병징과 고사를 일으키는 직접적인 원인을 제시한 바 있다.
소나무재선충의 밀도가 높았던 일부 묘목들은 건전한 묘목과 비교하여 잔뿌리가 부실하였거나, 목질부의 변색 부로부터 Alternaria sp.를 분리하였다(자료미제시). 따라 서, 기주식물이 외부환경요인이나 다른 식물병해충의 피 해를 입어 수세가 쇠약해진 상태에서 소나무재선충에 감 염된다면 고사가 더욱 빠르게 진행될 수도 있을 것으로 보인다. 실제로 Uzunovic 등(1999)은 변재변색균의 일종 으로서 Alternaria sp.를 보고한 바 있으나, 본 연구에서 확인된 목재변색이 소나무재선충 감염에 따른 것인지 또 는 목재변색이 소나무재선충의 증식을 촉진한 것인지는 앞으로 더 연구가 필요할 것이다. 한편, Maehara 등(2006) 과 Wang 등(2005)은 곰팡이 접종이 소나무 묘목 내에서 의 소나무재선충의 분포와 증식 등을 향상시킨다고 보고 한 바 있다.
접종 30일 후 감염목내에서 증식된 크기가 작은 소나 무재선충 유충은 가도관에서도 관찰이 되었고 목질부의 방사조직을 통과하는 소나무재선충의 유충도 확인되어 감 염후기에는 전부위에 퍼져나감을 확인하였다. 즉, 상처를 통해 묘목 내로 침입한 소나무재선충은 피층에 존재하는 수지도를 통해 접종부위로부터 빠르게 이동을 하고, 시간 이 경과함에 따라 조직 내에서 증식한 소나무재선충이 피 층수지도 뿐만 아니라 목질부수지도나 가도관까지 이동 하는 등 접종된 묘목의 모든 부위에 고루 분포하는 것으 로 나타났다.
요 약
소나무재선충에 자연 감염된 소나무와 잣나무로부터 소 나무재선충을 분리하여 수종 간 분리율과 수종 내 부위 별 밀도 등을 비교하고, 분리한 선충을 소나무 묘목에 접 종하여 묘목 내에서의 이동을 알고자 하였다. 2006년, 소 나무재선충에 감염되어 말라죽었거나 죽어가고 있는 소 나무와 잣나무로부터 소나무재선충을 분리한 결과, 소나 무재선충은 두 수종 모두에서 높은 밀도로 분리되어 수 종 간에 차이를 보이지 않았다. 또한, 각 수종의 변재부 Fig. 4. Re-isolation of Bursaphelenchus xylophilus from red pine
seedlings that were inoculated artificially 20 days before and showing different symptoms.
위에서만 분리되었으며, 분리한 조직을 채취한 줄기의 높 이에 따른 분리율에는 차이가 없었다. 즉, 말라죽은 나무 의 주간은 모든 조직이 소나무재선충에 감염되어 있었다.
소나무 묘목에의 접종실험을 통하여 감염목 내 이동을 조 사한 결과, 감염초기에 소나무 조직 내로 침입한 소나무 재선충은 증식에 앞서 감염목 내에서 빠르게 이동하여 퍼 지기 시작하였고, 그 이후에 왕성한 증식활동을 하였으며, 이 시기가 바로 소나무 시들음병의 외부병징이 나타나는 시기가 일치함을 알 수 있었다. 소나무재선충의 이동경로 를 조사하기 위해 접종한 소나무 줄기횡단면을 현미경으 로 관찰하고, 또 높이별로 선충을 분리하여 조직 내 선충 의 분포를 관찰한 결과, 줄기의 피층에 있는 수지도를 통 해 접종부위로부터 다른 부위로 빠르게 이동을 하고, 시 간이 경과하면서 조직 내에서 증식한 소나무재선충은 피 층의 수지도뿐만 아니라 목질부수지도나 가도관까지 분 포하여 감염목내 전 부위에 걸쳐 고루 분포하였다. 그러 나 수조직에서는 분리할 수 없었다. 소나무재선충 분리밀 도가 높았던 묘목은 뿌리의 발달이 불량하거나, 목질부 변색균에 감염된 것들이었으며, 비교적 튼튼한 외양을 한 묘목에서는 선충의 분리밀도가 낮았다.
Acknowledgements
This work was supported by the research grant of the Chungbuk National University in 2010.
References
Agrios, G. 2005. Plant Pathology, 5th ed., Elsevier Academic Press, Burlington, USA. 922 pp.
Fukuda, K. 1997. Physiological process of the symptom development and resistance mechanism in pine wilt disease. J.
For. Res. 2: 171−181.
Futai, K. 2008. Pine wilt in Japan: from first incidence to the present. In: Pine Wilt Disease. ed. by Zhao, B. G., Futai, K., Sutherland, J. R. and Takeuchi, Y., pp. 5−12. Springer, Tokyo, Japan.
Http://news.naver.com/main/read.nhn?mode=LSD&mid=
sec&sid1=102&oid=001&aid=0005551717
Ichihara, Y., Fukuda, K. and Suzuki, K. 2000. Early symptom development and histological changes associated with migration of Bursaphelenchus xylophilus in seedling tissues of Pinus thunbergii. Plant Dis. 84: 675−680.
Kawazu, K. and Kaneko, N. 1997. Asepsis of the pine wood nematode isolate OKD-3 causes it to lose its pathogenicity.
Jpn. J. Nematol, 27: 76−80.
Maehara, N., Tsuda, K., Yamasaki, M., Shirakikawa, S. and Futai, K. 2006. Effect of fungus inoculation on the number of Bursaphelenchus xylophilus (Nematoda:Aphelenchoididae) carried by Monochamus alternatus (Coleoptera: Cerambycidae).
Nematology 8: 59−67.
Mota, M., Braasch, H., Bravo, M. A., Penas, A. C., Burgermeister, W., Metge, K. and Sousa, E. 1999. First report of Bursaphelenchus xylophilus in Portugal and in Europe.
Nematology 1: 727−734.
Steiner, G. and Buhrer, E. H. 1934. Aphelenchoides xylophilus n.
sp., a nematode associated with blue-stain and other fungi in timber. J. Agric. Res. 48: 949−151.
Togashi, K., Matsunaga, K., Arakawa, Y. and Miyamoto, N. 2003.
The random dispersal of Bursaphelenchus xylophilus in pine twigs. Trans. Jpn. For. Soc. 114: 753.
Uzunovic, A., Yang, D. Q., Gagn, P., Breuil, C., Bernier, L., Byrne, A., Gignac, M. and Kim, S. H. 1999. Fungi that cause sapstain in Canadian softwoods. Can. J. Microbiol. 45: 914−
922.
Wang, Y., Yamada, T., Sakaue, D. and Suzuk, K. 2005. Influence of fungi on multiplication and distribution of the pine wood nematode, Bursaphelenchus xylophilus in axenized Pinus thunbergii cuttings. Nematology 7: 809−817.
Zhao, B. G., Wang, H. L. and Han, S. F. 2003. Distribution and pathogenicity of bacteria species carried by Bursaphelenchus xylophilus in China. Nematology 5: 899−906.
Zhao, B. G. and Lin, F. 2005. Mutualistic symbiosis between Bursaphelenchus xylophilus and bacteria of the genus Pseudomonas. Forest Pathol. 35: 339−345.
Zhao, B. G., Liu, Y. and Lin, F. 2007. Effect of bacteria associated with pine wood nematode (Bursaphelenchus xylophilus) on development and egg production of the nematode. J.
Phytopathol. 155: 26−30.