292 http://dx.doi.org/10.7850/jkso.2012.17.4.292
Free Access [종 설]
북서태평양에 서식하는 살오징어(Todarodes pacificus) 계군 분석에 대한 고찰
김정연·문창호·윤문근1·강창근2·김경렬3·나태희3·최은정4·이충일5,* 부경대학교 해양학과
1한국수산자원관리공단
2포항공과대학교 해양대학원
3서울대학교 자연과학대학 지구환경과학부/해양연구소
4극지연구소
5강릉원주대학교 해양자원육성학과
Stock Identification of Todarodes pacificus in Northwest Pacific
JEONG-YUN KIM, CHANG-HO MOON, MOON-GEUN YOON1, CHANG-KEUN KANG2, KYUNG-RYUL KIM3, TAEHEE NA3, EUN JUNG CHOY4AND CHUNG IL LEE5,*
Department of Oceanography, Pukyung National University, Busan 608-737, Korea
1Yangyang Salmon Station, Korea Fisheries Resources Agency, Yangyang 215-851, Korea
2POSTECH Ocean Science & Technology Institute, Pohang University of Sciences and Technology, Gyeongbuk 790-784, Korea
3School of Earth and Environmental Sciences/Research Institute of Oceanography, Seoul National University, Seoul 151-742, Korea
4Korea Polar Research Institute, KIOST, Incheon 406-840, Korea
5Department of Marine Bioscience, Gangneung-Wonju National University, Gangneung 201-702, Korea
본 종설논문은 살오징어의 기존 및 최근에 새롭게 적용되고 있는 계군 분석방법들을 비교 분석하여 각 분석방법 의 장단점과 분석방법간의 상호보완에 대하여 고찰하였다. 살오징어는 북서태평양의 넓은 지역을 회유하는 어종으 로 생태계 및 상업적으로 중요한 자원이다. 살오징어는 해양환경변화의 생물학적 지표로서의 가능성을 평가 받고 있 으며, 장단기적인 어획량 및 분포역의 변화가 환경 변화와 함께 나타난다. 예를 들어, 1987/1988 무렵에 발생한 기 후체제전환 이후 한류성 어종으로 분류되는 명태의 어획량은 급감하여 현재까지 그 영향이 지속되고 있는 반면, 살 오징어 어획량은 크게 증가하였다. 현재까지 명태 어획량의 감소에 대하여 남획과 기후변화에 초점이 맞추어진 해 석이 있으나, 뚜렷한 원인 분석은 이루어지지 않고 있다. 그 이유 중 한 가지는 계군 분석에 근거한 생태, 환경적 측 면에 대한 정확한 원인 분석이 이루어지지 않고 있는 것과 관련이 된다. 계군은 유사한 생물학적 특징을 가진 개체 들이 제한된 영역 내에서 유성생식과정을 통하여 동일한 유전자 풀(gene pool)을 공유하는 집단으로, 동일 계군을 형성하는 개체들은 산란에서 자원으로 가입 후 다시 재생산 과정에 이르기까지 시간 및 공간적으로 각기 다른 환경 의 영향을 받을 수 있다. 따라서, 종에 대한 정확한 계군 분석은 자원의 효과적인 관리 및 급격한 변화에 대한 중요 한 대응 방안의 역할을 할 수 있다. 살오징어 계군 분석에 적용된 주요 방법은 크게 4가지로 형태학적 방법, 생태학 적 방법, 표지방류법, 유전학적 방법이 있다. 형태학적인 방법은 분석방법이 가장 간단하고 다수의 개체를 비교적 쉽 게 분석할 수 있지만 각 형질들은 성장기간 동안 환경에 의해 영향을 많이 받게 되어 개체간의 차이가 생긴다. 생 태학적 방법은 주로 개체의 생리적인 변화와 분포 및 회유상태, 기생충의 기생상태나 종류 및 기생률 등을 분석, 산 란장의 차이를 알아보는 연구이며, 현재 활발히 연구되고 있는 방법으로 유사한 환경에서 생활하는 집단을 알 수 있 지만 유전적으로 같은 집단인지는 알기 어렵다. 표지방류법은 직접적인 방법으로 계군의 회유 및 분포, 산란장의 위 치를 파악할 수 있지만 수거가 어렵고 초기 단계에는 표식을 하기 어렵다. 수산생물의 계군 분석을 위한 유전학적 방법은 자원관리학적 연구에 관한 기본적 정보를 제공해 왔다. 계군 분석을 위한 유전학적 방법은 이에 사용하는 유 전자 마커(marker)의 감도에 따라 결정되며, 유전자 마커의 다형성이 높은 것을 선택해야 한다. 계군 분석을 위한 유전자 마커로는 오랜 기간 동안 동위효소 다형이 사용되어졌으며, 최근에는 mitochondria, microsatellite와 같이 DNA 염기배열 중에서도 변이성이 높은 영역을 선택하여 마커로 이용한 연구가 증가되고 있다. 기존의 형태학적 방 Received August 10, 2012; Revised October 6, 2012; Accepted October 6, 2012
*Corresponding author: [email protected]
법, 표지방류법, 생태학적인 방법들은 살오징어의 생활사, 회유경로, 산란장의 변화 등을 밝혀내어 계군을 파악하는 데 많은 기여를 하였지만 여전히 각 해역에 분포하는 살오징어의 계군을 파악하기에는 어려움이 있다. 최근에는 기 존의 계군 분석이 지닌 장단점을 비교 분석하여 복합적인 방법의 계군 분석이 이루어지며, 이러한 정보들을 바탕으 로 유전학적 방법을 보완한다면 살오징어 자원의 변동에 대한 관리 방안을 마련하는데 도움을 줄 것이다.
This paper reviews comparison analysis of current and latest application for stock identification methods of Todarodes pacificus, and the pros and cons of each method and consideration of how to compensate for each other. Todarodes pacificus which migrates wide areas in western North Pacific is important fishery resource eco- logically and commercially. Todarodes pacificus is also considered as ‘biological indicator’ of ocean environ- mental changes. And changes in its short and long term catch and distribution area occur along with environmental changes. For example, while the catch of pollack, a cold water fish, has dramatically decreased until today after the climate regime shift in 1987/1988, the catch of Todarodes pacificus has been dramatically increased. Regarding the decrease in pollack catch, overfishing and climate changes were considered as the main causes, but there has been no definite reason until today. One of the reasons why there is no definite answer is related with no proper analysis about ecological and environmental aspects based on stock identifica- tion. Subpopulation is a group sharing the same gene pool through sexual reproduction process within limited boundaries having similar ecological characteristics. Each individual with same stock might be affected by dif- ferent environment in temporal and spatial during the process of spawning, recruitment and then reproduction.
Thereby, accurate stock analysis about the species can play an efficient alternative to comply with effective resource management and rapid changes. Four main stock analysis were applied to Todarodes pacificus: Mor- phologic Method, Ecological Method, Tagging Method, Genetic Method. Ecological method is studies for anal- ysis of differences in spawning grounds by analysing the individual ecological change, distribution, migration status, parasitic state of parasite, kinds of parasite and parasite infection rate etc. Currently the method has been studying lively can identify the group in the similar environment. However It is difficult to know to identify the same genetic group in each other. Tagging Method is direct method. It can analyse cohort’s migration, distribu- tion and location of spawning, but it is very difficult to recapture tagged squids and hard to tag juveniles.
Genetic method, which is for useful fishery resource stock analysis has provided the basic information regard- ing resource management study. Genetic method for stock analysis is determined according to markers’ sensitiv- ity and need to select high multiform of genetic markers. For stock identification, isozyme multiform has been used for genetic markers. Recently there is increase in use of makers with high range variability among DNA sequencing like mitochondria, microsatellite. Even the current morphologic method, tagging method and eco- logical method played important rolls through finding Todarodes pacificus’ life cycle, migration route and changes in spawning grounds, it is still difficult to analyze the stock of Todarodes pacificus as those are distrib- uted in difference seas. Lately, by taking advantages of each stock analysis method, more complicated method is being applied. If based on such analysis and genetic method for improvement are played, there will be much advance in management system for the resource fluctuation of Todarodes pacificus.
Key words: Todarodes pacificus, Ecological characteristics, Stock identification method
서 론
살오징어(Todarodes pacificus)는 북서태평양의 주요 상업 어종 으로 북으로는 오호츠크해 남쪽, 쿠릴열도의 외측에서부터 동해, 서해, 동중국해, 그리고 남으로는 북서 필리핀, 남중국해의 북쪽까 지 분포한다(Araya, 1967; Okutani, 1983; Murata, 1990; Mokrin et al., 2002).
북서태평양의 넓은 지역을 회유, 분포하며 서식하는 살오징어는 연중 산란하며 주발생시기에 따라 여름에 발생하는 하계군, 가을 에 발생하는 추계군, 겨울에 발생하는 동계군으로 구분된다(Hamabe and Shimizu, 1966; Kasahara, 1982; Murata, 1990; Kim and Kang, 1995). 추계군과 동계군이 대다수를 차지하며 그 중에서도 동계군의 자원량이 가장 많은 것으로(Murata, 1989; Kawabata et al., 2006)
알려져 있다. Sakurai et al.(2002)는 기후체제전환(climate regime shift)에 의해 동계군 산란장이 한랭기에는 축소되고 온난기에는 확장되어 추계군 산란장과 동계군 산란장이 떨어져서 형성되는 한 랭기보다 추계군과 동계군 산란장이 확장되어 서로 겹쳐지는 온 난기에 자원량이 증가한다고 하였다.
Rosa et al.(2011)는 과거 한랭기에 전체 어획량은 감소하였지 만 서해의 어획비율은 증가하여 한랭기에는 해류에 의해 살오징 어 회유 경로가 변했을 것으로 추정하였다. 기후체제전환이 산란 장의 규모와 가입량 및 회유 경로의 변화에 영향을 끼치며, 그 결 과 해역별 어획량은 각기 다른 변화 경향을 나타내었다. 이는 각 해역을 회유하는 계군들의 비율이 다르고 환경변화에 따라 회유 하는 해역이 달라져서 생기는 차이로 추측된다. 이와 같이 환경변 화에 따른 자원량 관리를 효과적으로 하기 위해서는 해당 생물의
생태학적인 특성을 통한 생활사가 연구되어야 하며, 계군에 대한 이해와 적합한 계군 분석방법이 필요하다.
Zhang(2010)은 일정한 지리적인 분포구역 내에서 개체 상호간 의 임의교배를 통해서 동일한 유전자 풀(gene pool)을 공유함으로 써 일정한 유전자조성을 가지며, 동일한 생태학적 특성과 독자적 인 수량 변동의 양상을 보이는 집단이라고 계군에 대하여 정의하 였다. 현재까지 살오징어 계군 분석은 크게 형태학적 방법, 생태 학적 방법, 표지방류법, 유전학적 방법 등으로 연구되었다.
형태학적 방법은 외투장(Mantle length), 두부 길이(Head length), 체중(weight), 지느러미 길이(Fin length) 등을 측정하는 방법으로 다수의 개체들을 비교적 간편하게 연구할 수 있다. 하지만 생물은 성장환경에 의해 체장차이가 날 수 있고, 체장에 비해 성성숙도는 느릴 수 있다. 살오징어의 외투장과 체중의 상대성장은 발육단계, 계절 및 분포 해역에 영향을 받으며(Murata, 1978), Kim(1990)은 이러한 외투장과 체중의 상대성장을 비교함으로 수컷은 계군별로 차이가 없지만 암컷은 동계군이 큰 것으로 설명하였다. 그리고 Kim et al.(1997)은 외투장을 기준으로 하여 형태적인 특징들을 하나씩 비교해 본 결과 계군이 다른 경우 모든 형질이 차이난다고 하였다.
생태학적 방법은 생물의 식성과 회유 및 분포 그리고 성성숙도 를 분석하여 산란기와 산란장을 파악하고 부화에서부터 사망까지 생물과 환경의 관계를 연구하는 방법이다. 한국주변해역에서 채집 된 살오징어의 식성은 어류를 가장 많이 섭이하고, 계군에 따른 차이는 없었다(Kim and Kang, 1998). 부산주변해역에서 채집된 살오징어는 외투장의 크기에 따라 먹이 생물의 조성이 다르고, 계 절에 따라서는 산란기에 관계없이 연중 섭이하였다(Song et al., 2006). DMSP/OLS영상은 야간조업어선의 불빛을 이용하여 어선의 분포 및 어획량에 근거한 어장 중심을 추정하는 방법으로, 회유 경로의 변화를 파악하는 것이 가능하다(Kiyofuji et al., 2004; Choi et al., 2008). 그리고 성숙된 살오징어는 교미하여 알을 낳고(Murata, 1990; Sakurai et al., 2003) 그 알이 부화하는 과정에 대한 연구 (Sakurai et al., 1996; Sakurai, 2006)들이 이루어졌으며, 갓 태어난 유생들의 분포(Goto, 2002; Kim, 2008)에 대한 분석은 산란장 추 정에도 이용되었다(Sakurai et al., 2000; Rosa et al., 2011). 그 밖 에도 Takahara and Sakurai (2010)는 특정해역에 널리 분포하며 그 지역의 대표성을 띄는 기생충의 기생률을 통해 살오징어 섭이 회유경로를 파악하였다.
표지방류법은 표식을 직접 붙이거나 삽입하는 방법과 표식이 육 안으로 보이게 표시만 하는 방법이 있으며, 전기를 이용한 표식, 화학적 표식, 자연적인 표식 등 다양한 방법들이 사용되고 있다.
표지방류법은 넓은 지역의 이동 상태를 볼 수 없는 경우에 유용 하지만 수거가 어렵고 작은 개체에는 표식하기가 어렵다. 표지방 류법에 의한 계군 분석은 주로 표식을 직접 붙여서 방류하였다가 다시 잡는(recapture) 방식으로 진행되었다(Kasahara and Ito, 1968;
Osako and Murata, 1983; Mori and Nakamura, 2001; Kidokoro et al., 2010).
최근에는 생물집단의 유전자 다양성을 이용하여 수산생물의 자 원관리 및 보존을 위한 유전학적 계군 분석이 활발히 진행되고 있 다. 유전자 다양성은 환경변화에 대한 생물의 적응력과 생물진화 에 대한 정보를 제공한다. 지금까지 유전학적 계군 분석은 대부분 동위효소(isozyme)를 마커(marker)로 이용하였지만(Kang et al.,
1996), 분자생물학 기술이 발달하면서 대량의 유전정보를 처리가 능하고, 생물 내 존재하는 다량의 염기변이를 검출할 수 있게 되 었다. 이러한 유전정보는 수산생물의 계군 분석을 위한 새로운 마 커를 개발하는데 있어 많은 정보를 제공해 주고 있다. 유전학적 계군 분석은 유전자 마커의 감도가 그 성과를 좌우하고, DNA상 염기 변이성이 높은 영역을 가지는 mitochondrial DNA (mtDNA)와 microsatellite DNA (msDNA)마커 등으로 주로 연구되고 있다 (Dunham, 2004). mtDNA는 재조합 과정이 없는 모계유전을 하는 특징과 종간 또는 종내 집단 사이에 높은 돌연변이율을 보여, 종 내의 개체군 또는 종간에 진화적 유연관계를 밝히고 이해하는데 매우 유용하게 사용되고 있다(Kocher and Stepien, 1997). mtDNA 에는 자체적인 유전서열이 있고, 그 중에서도 cytochrome coxidase subunit I (COI)영역은 다양한 해양 무척추동물들의 개체군 유전 학 연구들에서 널리 사용되어져온 마커이다(Edmands et al., 1996).
현재까지 DNA를 이용한 살오징어 계군 분석은 mtDNA COI영역 을 이용한 연구(Kim, 2012)가 시행되었으며 다양한 마커들을 이 용하여 연구할 필요가 있다. 유전학적 방법은 개체들 간의 유전적 인 차이점을 알 수 있어 유사한 환경에서 발생된 계군을 유추할 수 있지만 부화 후의 환경적 영향으로 인한 변화는 알 수 없으며 , 많은 유전자 영역 중 특별한 변화를 보이는 영역을 찾기 힘들뿐 아니라 알맞은 영역을 찾기 위해서는 앞으로 다양한 DNA분석 방 법들을 적용하여 유전학적 정보들을 축적하는 것이 바람직하다.
살오징어는 현재 우리나라에 중요한 상업어종으로 지속적인 자원 유지를 위해 계군을 구분하는 작업이 필요하며, 계군 분석시 이용 된 사례들을 통하여 각 분석방법의 장단점을 파악하고 상호보완 된 분석방법을 개발할 필요가 있다.
살오징어의 생태학적 특성 및 계군
살오징어는 광온성 생물로 서식수온이 4-27oC이며, 주서식수층 은 표층으로부터 약 200 m 정도이다. 수명은 약 1년이며 산란 후 사망한다(Roper et al., 1969; Voss, 1977). 산란은 18-24oC 수온 범위의 대륙붕이나 대륙사면(100-500m) 상층부지역에서 이루어지 고 젤라틴질의 반부유성 난괴를 방출하며 19.5-23oC에서 부화 후 생존율이 가장 높다(Sakurai, 2006)(Fig. 1). 난괴 초기발달시기의
Fig. 1. Reproductive hypothesis for the Japanese common squid, Todarodes pacificus, initially proposed by Sakurai et al. (2000) and complemented by Sakurai (2006) (from Sakutai, 2006).
수온범위는 18-24oC이고 4-9.5일 지속되다가 대부분의 유생이 25- 50 m 깊이에서 발생된다. 부화된 유생은 표층으로 떠올랐다가 시 간이 경과함에 따라 서식 수심이 조금씩 깊어져 수심 50 m에서 주로 분포한다(Yamamoto et al., 2007). 살오징어 유생은 일시성 플랑크톤으로 유영능력이 미약하여 분포와 이동이 주로 해류에 의 해 결정되는데 일본의 동쪽은 쿠로시오의 영향을 받고 일본의 서 쪽(동해)은 쓰시마 난류의 영향을 받는다. 살오징어 유생의 분포 는 해류의 영향을 크게 받는데, Bower et al.(1999)는 쿠로시오 해 류와 연안수의 전선역에서 그 분포 밀도가 높다고 하였다. 특히 두 수괴가 만나 수온전선 및 혼합수역을 이루는 지역에서는 수렴 류에 의한 부유성 생물들의 축적에 의해 유생의 밀도가 높게 나 타난다(Yamamoto et al., 2002; Kim et al., 2011). 부화 후 15일 정도의 유생기간을 거치면 어린오징어로 변태(Doi and Kawakami, 1978)하고, 부화 후 5개월이 지나면 외투장 14 cm 정도로 성장한 다(Nakamura, 1985). 유영능력을 가진 살오징어는 어군을 형성하 여 먹이를 찾아 북상 회유하고 서해, 동해, 북서태평양에서 각각 성장하는데 식성은 성장단계 및 환경에 따라 바뀌고 주로 어류, 갑각류, 두족류, 다모류 등을 섭이한다(Kim and Kang, 1998; Song et al., 2006).
살오징어 성어는 산란을 위해 남하 회유하는데 암컷의 성숙은 15oC 이상의 수온분포역을 따라 회유할 때 빠르게 나타난다 (Kidokoro et al., 2010). 수컷은 암컷보다 약 3개월 일찍 성숙되 고(Araya, 1967), 각 계군의 성숙외투장은 조금씩 차이가 있는데 동중국해에서 어획된 오징어는 암컷, 수컷 모두 외투장 15 cm부 터 성숙되었다(Shojima, 1972). 수조 실험에서 관찰된 산란을 보 면 암컷은 산란 이틀 전에 먹이를 먹지 않고 수조바닥에서 휴식
을 취하였다가 떠올라 알을 낳았다(Bower and Sakurai, 1996). 살 오징어는 산란 후 사망하는데 Hamabe and Shimizu(1966)의 연구 에 의하면 100-500 m의 대륙붕이나 대륙사면 지형의 바닥을 트롤 로 끌었을 때 산란 후의 암컷들이 종종 잡혔으며, 이를 통해 일본 주위의 대륙붕이나 대륙사면 지역이 산란장으로 유추되기 시작하 였다. 따라서 21-40oN, 121-142oE, 수심 100-500 m의 쿠로시오 평 균축이 지나가는 수온범위 19.5-23oC인 지역이다(Rosa et al., 2011).
살오징어는 연중산란 하지만 주로 발생 시기에 따라 크게 하계 군, 추계군, 동계군의 세 계군으로 구분된다(Okutani, 1983; Murata, 1990; Kim and Kang, 1995; Choi, 2005). 하계군은 회유 범위가 다른 계군에 비해 좁고, 성숙외투장이 다른 계군보다 작다. 하계 군은 태평양과 동해의 극히 한정된 해역에서 5-8월경에 소규모로 산란을 하고 쓰시마난류와 연안류의 영향을 받으며 주로 연안역 에 서식한다. 동해에서는 사토(Sato)에서 오키(Oki)까지, 북태평양 에서는 보소(Boso)에서 이즈(Izu)까지 각각 분포한다. 유생 시기 는 5-9월이고 8-10월은 어린오징어 시기이며, 11-3월에 미성숙단 계를 거쳐 4-8월에는 성숙단계에 도달하게 된다(Okutani, 1983;
Murata, 1990).
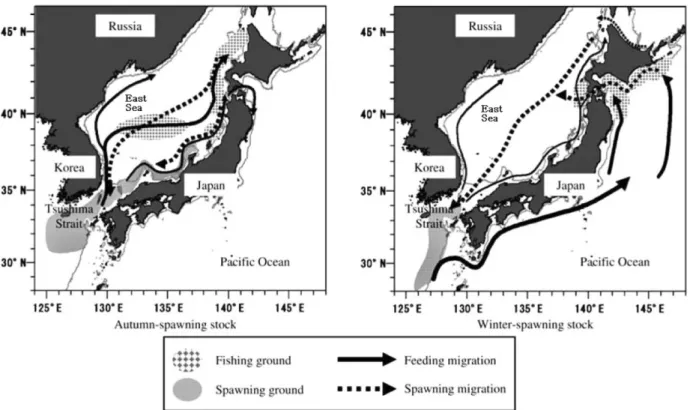
추계군은 산란장이 동해의 일본 연안과 우리나라 연안, 큐슈남 쪽까지이며, 9월에서 12월 산란하고 부화된 유생들은 쓰시마 난 류에 의해 북쪽으로 수송되면서 점차 성장하여 겨울에 훗카이도 에 도달한다. 추계군은 훗카이도 서쪽을 주 섭이장으로 하며 산란 시기가 오면 남쪽으로 산란회유한다. 유생 시기는 9-12월이고 11- 1월은 어린오징어 시기이며, 2-6월에 미성숙단계를 거쳐 7-11월에 는 성숙단계에 도달하게 된다(Okutani, 1983; Murata, 1990).
Fig. 2. Schematic diagram of the distribution range and migration route of the autumn-and winter-spawning stocks of Japanese common squid, Todarodes pacificus (modified from Kidokoro et al., 2010).
동계군의 산란장은 큐슈에서 타이완까지의 동중국해이고, 1월 에서 4월에 산란 후 쿠로시오를 따라 북상회유하여 오야시오역을 주 섭이장으로 하며 늦여름과 초가을사이 산란을 위해 다시 회유 한다. 동계군은 북서태평양에 대규모 분포하며, 서해로도 유입되 어 계군 중 가장 넓은 지역을 회유한다. 유생 시기는 1-4월이고 3- 5월은 어린오징어 시기이며, 6-10월에 미성숙단계를 거쳐 11-3월 에는 성숙단계에 도달하게 된다(Okutani, 1983; Murata, 1990).
살오징어는 크게 세 계군으로 구분되지만 대부분이 추계군과 동 계군으로 구성되며 현재까지의 연구결과들을 종합해보면 동계군의 자원량이 가장 많다(Araya, 1967; Kasahara, 1978; Okutani, 1983, Murata, 1989). 추계군과 동계군의 회유경로를 살펴보면, 추계군 은 쓰시마 난류를 타고 북쪽으로 이동하여 동해의 극전선역 및 훗 카이도 주변역이 주 섭이장이 된다. 그리고 동계군은 일부가 쓰시 마 난류를 타고 이동하며, 대부분은 일본의 동쪽연안으로 쿠로시 오를 타고 이동하면서 성장하고 오야시오와 쿠로시오가 만나는 경 계지역이 주 섭이장이 된다. 두 계군 모두 산란시기가 되면 남쪽 으로 산란회유를 하는데 그 해의 환경에 따라 산란장의 규모가 달 라지며(Fig. 2), 이는 어획량의 장단기변동과도 밀접한 관련이 있 다(Choi et al., 2008). 살오징어 어획량은 어구 및 어법의 발전 등 과 같은 인위적인 요소(Zhang and Lee, 2004; Kim et al., 2010)
뿐만 아니라 기후체제전환에 따른 해양환경변화에 의해 영향을 받 기도 한다(Sakurai et al., 2000; Rosa et al., 2011). 온난기에는 산 란장이 확장되어 유생의 양이 증가함에 따라 자원량도 증가하였 지만 한랭기에는 산란장의 규모가 축소되고 유생의 양이 감소하 면서 자원량 또한 줄어들었다(Sakurai et al., 2000). 지금까지 추 정된 추계군의 산란장은 한국의 동해연안, 동해의 일본연안과 큐 슈남부까지이고, 동계군의 산란장은 큐슈부터 타이완까지이다. 한 랭기와 온난기에 추계군의 산란장은 거의 변화가 없었지만 동계 군의 산란장은 크기와 위치에 큰 변화를 보였으며 온난기에 추계 군과 동계군의 산란장이 겹쳤을 때는 자원가입 성공률이 높았지 만 한랭기에 동계군의 산란장이 떨어져있을 때는 자원가입 성공 률이 매우 낮게 나타났다(Sakurai et al., 2002)(Fig. 3).
계군 분석 형태 및 생태학적 계군 분석
서론에서 언급하였듯이 지금까지 적용되어온 살오징어 계군 분 석법은 형태학적 방법, 생태학적 방법, 표지방류법, 유전학적 방법 등이다. 형태학적 분석방법은 계군 분석이 용이하여 다수의 개체 를 쉽게 분석할 수 있고, 기존의 분석된 계군을 잘 반영하는 반면, 성 Fig. 3. Possible scenario on stock fluctuation of Todarodes pacificus related to climatic regime shifts (Right: warm regime. Left: cool regime). Arrows represent migration routes (from Sakurai et al., 2002).
장은 섭이장에서 이미 성체에 이르지만 성숙은 산란회유 중 급속 히 이루어지므로 외투장만으로 계군을 구분하기에는 오차가 크다 는 한계가 있다. 이러한 형태학적 분석방법이 갖는 단점을 보완하 기 위해 생태학적 방법이 함께 연구되었으며, 생태학적 분석방법 은 생물 상호간의 관계나 생물과 환경과의 관계를 통해 계군을 분 석하는 방법으로 다양한 연구가 이루어지고 있다.
외투장 조성과 생식소 발달 상태로 비교한 연구(Kim and Kang, 1995; Choi, 2005)는 연중 계절별로 외투장이 유체로 가입되는 시 기가 크게 세 모드로 나타나 계절별 발생 시기에 따라 세 계군이 있다는 가설을 뒷받침한다. Choi(2005)는 동해와 서해에서 외투장 과 생식소 발달에 따라 계군을 분석하였다. 그 결과 동해에서는 3 개의 계군이 존재하는 것으로 나타났다. 9월에는 하나의 계군(추 계군)이 그리고 5, 7, 12월은 3개의 군이 모두 존재하였으며, 나머 지 달에는 차이가 나는 2개의 군이 나타났다. 서해에서는 1987년 에 6-12월에 어획한 살오징어의 발생군을 분석해 본 결과 추계군 과 동계군으로 구성되었다. 동해에서는 3개의 계군이 모두 출현하 였지만 서해에서는 하계군을 제외한 추계군과 동계군만이 어획되 었으며 동해보다 서해에서 어획된 살오징어가 더 빨리 성숙되었 다고 보고했다.
Kim and Kang(1995)은 1991년 2월부터 1992년 7월까지 남해, 동 해에서 살오징어를 채집하여 외투장과 생식소 발달상태에 의해 계 군을 분리하였다. 그 결과 외투장이 6-12 cm인 유체의 주 가입 시 기는 5-6월, 8-9월, 1-3월로 세 개의 계군으로 분류되었다. 1991년 5월, 10월에는 각각 동계군, 추계군 판단되는 계군이 하나만 보였 으며, 1991년 4, 8월, 1992년 6, 7월에 세 개의 계군으로 나타났 고 나머지는 모두 차이가 나는 2개의 계군이 존재하였다. 지금까 지의 연구를 종합해보면 과거의 형태 및 생태학적 분석결과에서 는 연도가 다르지만 연중 세 번의 가입이 있었고 각 계군이 중복 해서 존재하는 시기가 많은 것이 확인되었다. 따라서 살오징어는 계절적으로 크게 세 계군이 존재하며, 계군이 중복되는 시기가 많 으므로 계군 분석시 각 군별 오차가 클 것이라고 판단된다.
Murayama et al.(1993)은 동해에서 월별 어획량변화, 외투장, 살 오징어 분포와 먹이생물 사이의 관계를 살펴봄으로써, 계절적으로 변하는 먹이분포가 산란양을 달라지게 한다고 하였다. 이를 증명 하기 위해 훗카이도 태평양쪽에 분포하는 살오징어의 월별 외투 장 변화(Araya and Ishii, 1972)와 동해의 월별 외투장 변화를 비 교하였고, 그 결과 태평양에서 어획된 살오징어 외투장의 최빈값 은 동해에서 두 달 전에 어획된 것과 거의 같았다. 이런 평균은 주 로 동해에 서식하는 계군이 태평양에 서식하는 계군보다 두 달 먼 저 태어난 것으로 추측되며, 이를 통해 동해에는 추계군, 북서태 평양에는 동계군이 주로 서식하는 것으로 추정하였다.
유생의 분포(Goto, 2002; Kim, 2008; Kim et al., 2011)를 통해 계군을 분석하는 방법은 각 채집정점에서 유생을 채집하여 외투 장을 측정하면 부화 후 일령이 추정되고, 이를 통해 발생 시기를 판단한다. 부화한 유생의 외투장은 0.95 mm이고, 일주일 후에는 약 1.25 mm로 자라며(Watanabe et al., 1996), 부화 후 23일이 지 나면 외투장은 26 mm, 30일이 지나면 35 mm로 성장한다(Tanaka, 1988). 이로부터 유생시기 외투장 길이는 부화 후 일령을 알 수 있 는 기준으로 제시되었고 산란시기와 산란장을 추정하여 계군을 판 단한다. 하지만 유생 시기는 유영능력이 미약하여 주로 해류에 의
한 영향을 많이 받으며, 해류의 강약과 방향에 따라 유생의 분포 가 달라진다. 그리고 해류를 파악한다 해도 유생이 성장해감에 따 라 수평이동 뿐만 아니라 수직이동까지 하기 때문에 유생의 분포 와 이동을 알기위해서는 더 많은 연구가 필요하다. Sakurai(2006) 는 실험 및 현장조사를 바탕으로 살오징어 산란환경은 수심 100- 500 m의 경사진 지역, 수온은 18-24oC에서 산란이 활발히 일어나 고, 부화 후 생존율이 가장 높은 수온을 19.5-23oC라고 가정하였 으며 GIS (Geographic Information System)자료를 이용하여 이런 지역들을 산란장으로 추정했다. 북태평양에 위치한 각 계군의 살 오징어 산란장은 기후체제전환에 의해 규모와 위치가 변화되었는 데, 한랭기에 각 계군의 산란장은 비교적 규모가 작고 추계군과 동계군의 위치는 구분되었지만 온난기에는 산란장이 팽창되고 겹 쳐서 나타나는 경향을 보였다. 산란장 추정을 통한 살오징어의 계 군 분석은 온난기동안 유생의 분포가 증가하였을 때 자원량이 증 가했고, 산란장의 규모가 커졌을 때 추계군, 동계군의 자원량이 증 가했다(Sakurai et al., 2002; Rosa et al., 2011). 따라서 계절별로 크게 가을과 겨울에 산란이 일어나는 계군이 존재한다는 것은 추 정할 수 있지만 산란장이 겹쳐서 생성될 때는 두 계군의 구별은 어렵다고 판단된다.
따라서 형태 및 생태학적 계군 분석에서는 연중 계절별로 크게 3개의 계군으로 구분되지만, 두드러지게 나타난 부분을 제외하고 나머지부분은 빨리 태어난 개체와 늦게 태어난 개체가 뚜렷이 구 별되지 않았다. 그리고 추계군과 동계군의 주서식해역이 서로 다 르며, 기후체제전환에 의해 산란장 규모가 결정되고 이는 각 계군 의 자원량이나 계군식별에 영향을 미친다고 파악된다. 해양생물은 수온과 먹이 등의 환경에 따라 성장과 성숙에 차이가 나타나며, 환경이 변했을 때의 자원의 변화는 예측하기 어렵다.
표지방류를 이용한 계군 분석
살오징어의 표지방류법은 주로 산란회유하는 개체들을 잡아 표 지를 부착한 후 놓아 주었다가 다시 잡는 방식으로 시행되었으며, 넓 은 지역의 자원량 추정이 불가능할 때나 집단의 회유경로를 알고 싶을 때 유용하다. 어획량이 증가했던 시기에 표지방류 회수율은 그렇지 못했던 해의 회수율보다 감소했고(Nagasawa et al., 1993), 1988/1989 기후체제전환 이후에 혼슈중앙부로 내려오던 추계군이 한국동해연안으로 이동하는 등 산란회유경로의 변화가 나타났다 (Kidokoro et al., 2010). 이러한 표지방류법은 환경변화에 따른 회 유경로의 변화를 설명해준다. 하지만 이 방법은 대규모로 조사해 야하므로 비용이 많이 들며, 국가간에 공조가 이루어지지 않을 경 우 수거에 어려움이 있다. 표지방류법은 표지를 부착하는 방식 외 에도 전기를 이용한 표식, 화학적 표식, 자연적인 표식 등의 다양 한 방법들이 있으며 그 중에 기생충을 생물학적 표식으로 사용한 연구가 최근 활발하게 이루어지고 있는데 이를 위해서는 목표로 하는 기생충의 생활사에 대한 이해가 우선적으로 필요하다. 기생 충은 특정 생물의 채내 혹은 체표에서 생활하며 그 생물로부터 영 양분을 취하여 살아가는데 이런 생활사를 이용하여 숙주인 생물 의 이동경로, 식성, 서식 환경의 변화 등을 알아볼 수 있다. 이렇 듯 기생충은 생물학적 표식으로 이용하기 적합하며 인공표식처럼 비용이 많이 들지 않고 생물에 직접 표식작업을 할 필요가 없으 므로 경제적이고 유용한 표식으로 사용가능하다. 하지만 모든 기
생충을 생물학적 표식으로 사용할 수는 없는데 생물학적 표식으 로 사용될 수 있는 기생충의 조건으로는 비교 지역 간의 기생률 차이를 나타내고, 숙주로부터 쉽게 떨어지거나 소실되지 않아야 하며, 채집방법이 간단하고 특정부위에만 기생하면서, 유사한 다 른 종이 없어 동정하기 용이해야한다. 그리고 연구기간 동안 사멸 하지 않을 정도의 생존이 가능하고 쉽게 숙주를 옮기지 않아야 하 며, 숙주의 생사나 활동에 영향을 주지 않고 생활사가 복잡하지 않아야 한다(Mackenzie, 1983; Kim, 2007). Pascual and Hochberg (1996)는 두족류 중 대표성을 띄는 지역별, 어종별의 기생충을 조 사하여 정리하였고, 생물학적 표식으로 계군 분석에 사용하였으며, 이를 통해 Takahara and Sakurai (2010)는 섭이장 차이에 따른 기 생충의 기생률로 추계군과 동계군을 구분하였다. 기생충은 해양생 물들의 계군을 분석하는 연구들에 많이 이용되고 있으며(MacKenzie et al., 2008), Takahara and Sakurai (2010)는 2008년 동해에서 채 집한 살오징어에 감염되는 anisakis를 조사하였다. 그 중에서 Anisakis simplex와 Lappetascaris sp. 2종에 의해 감염이 되었는데 Anisakis simplex에 의한 감염은 동계군이 추계군보다 더 심했다.
그 이유는 오호츠크해와 오야시오 지역에서 Anisakis simplex에 의 해 감염된 명태를 먹이로 하기 때문이다. 하지만 추계군은 이 지 역을 회유하지 않으며 훗카이도 근처가 섭이장인 두 계군 모두에 서 Lappetascaris sp.가 많이 발생하여 감염된 기생충의 비율차이 로 섭이장의 위치를 추측하였다(Fig. 4). 감염된 기생충의 기생률 은 섭이장이 다른 추계군과 동계군을 구분하는 기준이 될 수 있 다. 하지만 같은 섭이장을 이용하는 추계군과 동계군의 차이는 알 수 없으며, 환경변화로 인해 먹이로 이용하는 특정어종의 자원량 에 문제가 생기거나 주된 먹이가 변하는 경우에는 기생충의 기생 률이 달라져 계군의 구분이 어렵다. 기생충을 이용한 방법은 수거
가 어렵고 비용이 많이 드는 표지방류법의 단점을 보완하는 대신 환경에 따라 변할 수 있는 생태학적 분석방법의 한계를 포함한다.
유전학적 분석방법
집단유전학을 이용한 연구는 생물의 몸 속에 있는 유전정보를 통하여 종의 진화를 알아보는 방법이다. 생물집단은 유전적으로 완전히 분리되어 있지 않고 번식을 통해 유전자 교환이 일어나 유 전자 흐름(gene flow)양상에 따라 생기는 유전적 차이점을 비교해 봄으로써 계군을 결정하고 이를 통해 살오징어의 유전학적 특징 을 알 수 있다. 집단유전학을 통한 계군 분석에는 효소, 단백질, mtDNA, 핵DNA를 이용할 수 있으며, 이러한 분석방법은 태어날 때부터 가지고 있는 고유의 유전자를 비교하는 방법이기 때문에, 언제, 어디서 어획되든 고유의 자료를 가진다. 따라서 유전학적 계 군 분석은 각국의 한정된 해역에서만 어획해야하는 현 상황에 기 존 분석방법이 가지는 한계점을 보완해 줄 계군 분석 방법으로 판 단된다. 그 동안 연구된 살오징어의 집단유전학을 통한 계군 분석 은 특정효소를 이용한 isozyme분석법으로 시행한 연구(Kang et al., 1996)와 전기영동법에 의한 근육 단백질과 먹물 단백질의 polypeptide 수준으로 계군을 분석한 연구가 이루어졌다(Kim, 1990). 하지만 isozyme다형은 DNA상의 구조영역에 존재하여 생 물적 기능을 담당하는 부분이며 짧은 시간 내에 많은 시료를 처 리 할 수 있지만 조직 샘플시 조작이 어려우며, 단형적 유전자좌 (gene loci)가 많고 다형적 유전자좌일지라도 변이성이 떨어져 생 식격리 후의 시간이 짧은 지역집단간의 차이를 검출하는데는 한 계가 있다(Brown et al., 1970; Park et al., 1993). 따라서 최근에 분자적 기술이 발전함에 따라 집단의 유전자 다양성을 평가하는 데 있어서 광범위하게 이용되고 있는 mtDNA와 msDNA 마커 등 Fig. 4. (a) Prevalence of nematodes in squid at each sampling site, (b) Map of northeast Asia showing nine areas where Todarodes pacificus were collected (modified from Takahara and Sakurai., 2010).
을 이용하여 재확인 해볼 필요가 있다고 판단된다. 계군 분석 연 구에 사용된 유전자 마커는 다형의 원인이 염기치환에 의한 것과 단위 배열이 반복한 수의 차이에 의한 것으로 구분된다. 특히, 염 기치환으로 유전적 다형을 검출하는 방법으로는 분화정도가 비교 적 빠른 mtDNA 및 단일염기변이 다형(SNP: Single Nucleotide Polymorphism)이 있으며, 단위 배열의 반복 수에 의한 방법으로 는 개체 간 다형이 높게 나타나는 msDNA가 있다. mtDNA는 DNA재조합 과정이 없는 모계유전을 하고 높은 돌연변이율을 보 여, 동물에 있어 종 내의 개체군 또는 종간에 진화적 유연관계를 밝히고 이해하는데 매우 유용하게 이용되고 있으며(Taberlet and Bouvet, 1994; Kocher and Stepien, 1997), 특히 어류의 집단 유 전학 연구에 있어서 유용한 marker로 이용되고 있다(Saito et al., 2008; Hamasaki et al., 2010). 그리고 msDNA마커는 핵DNA 염기 서열 상에서 2-6개의 염기가 반복적으로 나타나는 DNA로서 공우 성(codominant)으로 mtDNA에 비해 변이 정도가 크기 때문에 집 단유전학에서 강력한 유전자 마커로서 이용되어지고 있다(Avise et al., 2000). 이상에서 언급했듯이 현재 이용되고 있는 유전자 마 커는 매우 다양해서 실용성이 높은 검출방법을 선택할 필요성이 있다. 특히, 염기치환에 의한 다형과 단위배열 반복 수 다형 검출 법의 대표적인 마커는 초기비용이 많이 요구되지만 대량의 시료 를 처리해야 하는 점에선 타 마커보다 뛰어난 장점이 있다(Table 1). 따라서 마커 선택시 일반적으로 수산생물의 계군 분석에는 대 량의 샘플을 처리해야 하기 때문에 경제성 역시 고려해야 할 필 요성이 있다.
지금까지 오징어에 대한 유전학적 연구는 주로 분류학적 연구 가 주를 이루었고 계군 분석에 대한 연구는 극히 제한적으로 이 루어졌다. 이러한 원인으로는 대부분의 오징어에 대한 생활사 및 이동경로에 대한 연구가 미흡해 계군 분석에 필요한 생태학적 정 보를 얻기가 힘들기 때문으로 판단된다. 오징어의 유전학적 계군 분석을 통해 보고된 사례를 보면, Kassahn et al.(2003)은 mtDNA 및 msDNA 마커를 이용하여 호주연안에서 서식하는 오징어(Sepia apama)에 대한 계군 분석을 통해 이들이 크게 지역에 따라 서로
다른 계군이 존재하고 있으며, 형태학적으로 분석한 결과 이들 계 군이 서로 독립된 환경에 따른 형태변이가 함께 존재하고 있음을 밝혀냈다. 또한 Shaw et al.(2004)은 msDNA 마커를 이용하여 Patagonian squid (Loligo gahi)가 포클랜드 제도 주위에서는 계절 적, 지리적으로 구분된 계군이 유전적으로 종내 집단이었지만 남 동 태평양(페루)의 집단과는 유전적으로 아종 간으로 보고하였다.
살오징어의 유전학적 계군 분석으로 Kang et al.(1996)은 동위효 소(isozyme)를 이용한 연구를 통해 기존의 형태 및 생태학적으로 구분한 하계군, 추계군, 동계군은 해류, 번식생태 등 여러 요인 등 에 의해 유전자를 교환하는 집단으로 확인하였고, 그 중에서 하계 군과 추계군내에는 유전적 격리현상으로 인한 독립된 지역개체군 이 존재한다고 보고하였다. 또한 Kim(2012)은 mtDNA COI영역 을 마커로 사용하여 기존의 형태학적 분석방법으로 구분한 하계 군과 추계군은 유전적으로 같은 집단이고, 동해 내에 분포하는 추 계군내에는 유전적으로 분리된 집단이 존재함을 밝혔다. 따라서 살오징어를 기존의 형태 및 생태학적으로 분류한 계군은 3개의 계 군으로 구분되지만 유전적으로는 하나의 집단이었으며, 기존의 방 식으로 분류된 집단 내에서 유전적 격리현상으로 인해 구분되는 집단이 존재했다. 이와 유사한 사례로 Patagonian squid와 살오징 어를 비교해보면, 유전학적 계군 분석을 통해 계절적으로 구분된 계군은 유전적으로 종내 집단이었다. 하지만 Patagonian squid는 태평양 집단과 대서양 집단이 아종 간으로 나타났으므로 살오징 어 또한 북서태평양과 동해 집단 간의 유전학적 계군 분석을 할 필요가 있다. 이처럼 유전학적 계군 분석은 기존의 형태 및 생태 학적 계군 분석과는 다른 결과를 나타냈다. 살오징어는 단년생 생 물로 연중산란하며 분포 범위가 한정적인 회유성 어종이고, 산란 기에는 계군의 구분없이 성숙된 개체들 간의 교미가 이루어지는 것으로 추정된다. 또한 기후체제전환에 의해 산란장의 규모가 변하 였으며, 온난기에는 추계군과 동계군의 산란장이 겹쳐 계군이 서 로 혼합되었다고 판단된다.
살오징어의 유전학적 계군 분석은 형태 및 생태학적으로는 구분 할 수 없는 차이를 유전자의 차이를 통해 나누는 방법이다. 특히 살 Table 1. Relative costs of RFLP, AFLP, microsatellites and SNP at variable numbers of individuals sampled, markers tested and technicians required (from Dunham, 2004)
Fish/markers/technicians Costs (US$)
RFLP AFLP Microsatellites SNP
Start-up costs 6,358 92,320 98,104 96,327
10/1/1 31,889 40,402 41,040 40,670
10/3/1 32,056 40,402 41,278 40,839
10/10/1 32,752 40,402 41,995 41,125
10/50/1 36,205 40,402 75,693 42,581
100/1/1 32,094 40,976 41,212 40,774
100/3/1 32,487 40,976 41,457 40,978
100/10/1 33,207 40,976 42,380 41,473
100/50/1 37,480 40,976 47,691 43,269
1,000/1/2 64,605 74,964 73,753 72,343
1,000/3/2 65,318 74,964 73,917 72,667
1,000/10/2 69,685 74,964 77,571 73,442
10,000/1/4 150,494 177,848 139,349 146,536
10,000/50/4 370,000 177,848 349,221 214,866
오징어는 회유성 어종이므로 넓은 해역에 분포하는데 언제, 어디서 잡히던지 유전정보는 일정하므로 신뢰할 수 있는 자료가 된다. 하지 만 유전적인 정보만으로는 살오징어의 생활사나 자원량의 변화 등 은 판단하기 어려우며, 유전적으로 차이가 나는 경우도 집단 간의 유전적 차이의 존재여부만 알 수 있어, 그에 따른 원인을 찾기 위해 서는 기존에 시행된 계군 분석이외에도 해양과 대기의 폭넓은 환경 조사와 고해양학 등의 다양한 연구가 함께 이루어져야 할 것이다.
결 론
살오징어는 현재 생태학적 위치뿐만 아니라 상업어종으로 그 가 치가 중요하고 많이 연구되는 생물이며 각 해역별로 계절에 따라 산란장이나 해류가 변경되어 어획량에 차이가 나타난다. 따라서 보다 객관적인 어획량예측을 위해서는 계군 분석이 필요하다. 앞 서 설명하였듯이 계군을 분석하는 방법에는 크게 형태학적 방법, 생태학적 방법, 표지방류법, 유전학적 방법 등이 있다. 형태학적 방법은 분석방법이 용이한 대신 각 계군별 구분기준의 오차가 크 다는 단점이 있어 생식소 발달로부터 성숙도를 알아보는 생태학 적 방법과 함께 사용되어 발생 시기를 추정하고 계군을 분리하였 다. 그 밖에도 생태학적 방법은 식성, 회유 및 분포, 유생분포, 산 란장 추정 등의 다양한 방법으로 연구 가능하다. 하지만 형태 및 생태학적 방법은 환경에 따라 영향을 많이 받으므로 환경에 영향 을 받지 않는 기준과 함께 연구되어야한다. 표지방류법은 표지를 부착하여 넓은 지역의 회유경로를 볼 때 유용한 방법으로 환경 변 화시 살오징어의 회유가 어떻게 변하는지 판단 할 수 있지만, 국 가 간의 공조가 이루어지지 않아 수거가 어렵다는 단점이 있다.
유전학적 방법은 유전정보를 통해 알아보는 방법이기 때문에, 언 제, 어디서 어획되든 고유의 자료를 가지므로, 각국의 한정된 해 역에서만 어획해야하는 현 상황에 필요한 방법이다. 그러나 유전 정보만으로는 생활사나 자원량의 변화 등은 판단하기 어려우며, 유전적 차이가 나는 원인을 밝힐 수 있는 연구를 병행해야한다.
앞서 보았듯이 형태학적 분석에서는 다른 계군으로 구분되었지만 유전적으로 같은 집단으로 판단되며, 형태학적 분석으로는 같은 계군이었지만 동일한 해역 내에서 유전적으로 차이가 나는 집단 이 존재했다. 그 동안 각 해역별, 시기별로 유전학적 계군 분석은 극히 제한적으로 이루어졌으며, 유전학적인 정보가 부족하여 같은 계군 내에서 유전적으로 분리된 집단이 존재하지만, 몇 개의 집단 으로 나눠지고, 각 해역에 따라서는 어떠한 차이가 생기는지에 대 해서는 밝힐 수 없었다. 각각의 계군 분석방법에는 장단점이 있으 며, 그 동안 한 방법만 사용하기보다는 주로 형태학적 계군 분석 을 토대로 다른 분석방법들을 비교하여 분류하였고, 주로 성체 위 주의 국지적인 조사만이 이루어졌다.
앞으로는 계군 분석은 성장시기별로 장기간의 광역적인 조사를 시행하고 그 동안 미흡했던 표지방류법을 포함한 복합적인 방법 의 접근이 필요하다. 또한 기존의 분석결과와는 다른 유전학적으 로 계군이 어떻게 구분되는지 알아보고, 다양한 마커의 개발을 통 해 분자생물학적 정보들을 축적하여야 한다. 그리고 해양과 대기 의 폭넓은 환경조사와 고해양학 등의 연구를 이용하여 유전학적 계군 분석에 대한 해석을 찾는다면 차후 각 계군사이의 관계 및 각 해역에 주로 서식하는 계군을 밝히는데 도움이 될 것이다.
사 사
본 연구는 2012년 국토해양부의 재원으로 한국해양과학기술진 흥원의 지원을 받아 수행된 연구임(장기해양생태계 연구: 환경변 화와 생태계 반응, 동해 시계열관측 및 생태환경진단(East-1))을 밝힙니다.
참고문헌(References)
Araya, H., 1967. Resources of common squid, Todarodes pacificus, Steenstrup in the Japanese waters. J. Jan. Fish. Res., 16: 60. (in Japanese)
Araya, H. and T. Ishii, 1972. Population structure of common squid in the waters around Hokkaido. Res. Rep. Tech. Couns. Agr.
For. Fish., 57: 192−205. (in Japanese)
Avise, J.C., 1994. Molecular markers, Natural History and Evolu- tion. Chapman and Hall, New York.
Bower, J.R. and Y. Sakurai, 1996. Laboratory observations on Todarodes pacificus (Cephalopoda: Ommastrephidae) egg masses. Amer. Mal. Bull., 13: 65−71.
Bower, J.R, Y. Nakamura, K. Mori, J. Yamamoto, T. Isoda and Y.
Sakurai, 1999. Distribution of Todarodes pacificus (Cepha- lopoda: Ommastrephidae) paralarvae near the Kuroshio off southern Kyushu, Japan. Mar. Biol., 135: 99−106.
Brown, W.M., M. George, Jr and A.C. Wilson, 1979. Rapid evo- lution of animal mitochondrial DNA. Proc. Nat'l. Acad. Sci.
USA, 76: 1967−1971.
Choi, K.H., 2005. Fishing conditions of common squid, Todarodes pacificus (Steenstrup) in relation to oceanic conditions in Korean Waters. Ph.D.Thesis. Pukyong National University, Busan, 81 pp.
Choi, K.H., C.I. Lee, K.S. Hwang, S.W. Kim, J.H. Park and Y.
Gong, 2008. Distribution and migration of Japanese common squid, Todarodes pacificus, in southwest part of the East (Japan)sea. Fish. Res., 91: 281−290.
Doi, T. and T. Kawakami, 1978. Biomass of Japanese common squid Todarodes pacificus Steenstrup and the management of its Fishery. Bull. Tokai. Reg. Lab., 99: 65−83. (in Japanese) Dunham, R.A., 2004. Aquaculture and fisheries biotechnology.
CABI Publishing press, Wallingford, pp. 104−109.
Edmands, S., P. E. Moberg and R. S. Burton, 1996. Allozyme and mitochondrial DNAevidence of population subdivision in the purple sea urchin Stronglocentrotus purpuratus. Mar. Biol., 126: 443−450.
Goto, T., 2002. Paralarval distribution of Ommastrephid squid Todarodes pacificus uring fall in the southern sea of Japan, and its implication for locating spawning rounds. Bull. Mar. Sci., 71: 299−312.
Hamabe, M. and T. Shimizu, 1966. Ecological studies on the com- mon squid, Todarodes pacificus Steenstrup, mainly in the south- western waters of Japan Sea. ull. Jap. Reg. Fish. Res. Lab., 16:
13−55.
Hamasaki, K., S. Kitada, S. Toriya., H. Shishidou and T. Sugaya, 2010. Genetic effects of hatchery fish on wild population in red
sea bream Pagrus major (Perciformes, Sparidae) inferred from a partial sequence of mitochondrial DNA. J.Fish. Biol., 77:
2123−2136.
Kang, Y.J., Y.H. Kim, Y.K. Hong, J.Y. Park and K.Y. Park, 1996.
A population genetic analysis of the common Squid, Todarodes pacificus Steenstrup in the Korean waters. J. Kor. Fish. Soc., 29: 320−331.
Kasahara, S. and S. Ito, 1968. Studies on the migration of common squids in the Japan Sea. 2. Migrations and some biological aspects of common squids having occurred in the offshore region of the Japan Sea during the autumn season of 1966 and 1967. Bull. Reg. Fish. Res. Lab., 20: 49−70.
Kasahara, S., 1978. Descriptions of offshore Squid Angling in the Sea of Japan, with Special Reference to the Distribution of Common Squid and on the Techniques or Forecasting Fishing Conditions. Bull. Jan. Reg. Fish. Res. Lab., 29: 179−199.
Kasahara, S., 1982. Population structure of Japanese common squid, Todarodes pacificus Steenstrup, in the Tsushima Warm Current area. In S.56 nendo ikarui shigen gyokaikyo kentokaigi gijiroku, Hokkaido Reg. Fish. Res. Lab. Kushiro. Japan, 47 p.
(in Japanese)
Kassahn, K.S., S.C. Donnellan, A.J. Fowler, K.C. Hall, M. Adams and P.W. Shaw, 2003. Molecular and morphological analyses of the cuttlefish Sepia apama indicate a complex population struc- ture. Mar. Ecol., 143: 947−962.
Kawabata, A., A. Yatsu, Y. Ueno, S. Suyama and Y. Kurita, 2006.
Spatial distribution of the Japanese common squid, Todarodes pacificus, during its migration in the western North Pacific Ocean. Fish. Oceanogr., 15: 113−124.
Kidokoro, H., T. Goto, T. Nagasawa, H. Nishida, T. Akamine and Y. Sakurai, 2010. Impact of a climate regime shift on the migra- tion of Japanese common squid (Todarodes pacificus) in the Sea of Japan. ICES J. Mar. Sci., 67: 1314−1322.
Kim, J.H., 2007. Use of parasites for stock analysis of salmonid fishes. J. Kor. Soc. Oce., 12(2): 112−120.
Kim, J.J., 2008. Summer Occurrence and Transport Process of Common Squid (Todarodes pacificus) Paralarvae in the East China Sea. M.S.Thesis. Pukyong National University, Busan, Korea. 61 pp.
Kim, J.J., H.H. Lee., S. Kim and C. Park, 2011. Distribution of lar- vae of the common squid Todarodes pacificus in the northern east china sea. Kor. J. Fish. Aquat. Sci., 44(3): 267−275.
Kim, J.Y., 2012. Stock Analysis of Todarodes pacificus in Korean waters. M.S.Thesis. Pukyong National University, Busan, 51 pp.
Kim, Y.H. and Y.J. Kang, 1995. Population analysis of the common squid, Todarodes pacificus Steenstrup in Korean waters 1. Sep- eration of population J. Kor. Fish. Soc., 28(2): 163−173.
Kim, Y.H., Y.J. Kang and C.I. Baik, 1997. Population analysis of the common squid, Todarodes pacificus Steenstrup in Korean waters 2. Morphological analysis J. Kor. Fish. Soc., 30(5): 903 pp.
Kim, Y.H. and Y.J. Kang, 1998. Stomach contents analysis of the common squid, Todarodes pacificus Steenstrup in Korean Waters. J. Kor. Fish. Soc., 31: 26−30.
Kim, Y.H., C. H. Moon, K. H. Choi and C. I. Lee, 2010. Rela- tionship between squid (Todarodes pacificus) catch by sea block and marine environment in the East Sea during 1980s and 1990s. J. Kor. Soc. Mar. Env. Saf., 3: 259−268.
Kim, Y.S., 1990. Population Analysis with Electrophoresis of Todarodes pacificus (STEENSTRUP) Collected from the East Sea and West Sea. M.S.Thesis, Inha University, Incheon, 46 pp.
Kocher, T.D. and C.A. Stephen, 1997. Molecular systematics of fish. Academic. press. New York, 99: 1−11.
Kiyofuji, H. and S.I. Saitoh, 2004. Use of nighttime visible images to detect Japanese common squid Todarodes pacificus fishing areas and potential migration routes in the Sea of Japan. Mar.
Ecol. Prog. Ser., 276: 173−186.
MacKenzie, K., 1983. Parasites as biological tags in fish population studies. Adv. Appl. Biol., 7: 251−331.
MacKenzie, K., N. Campbell, S. Mattiucci, P. Ramos, A.L. Pinto and P. Abaunza, 2008. Parasites as biological tags for stock identification of Atlantic horse mackerel Trachurus trachurus L. Fish. Res., 89: 136−145.
Mori, K. and Y. Nakamura, 2001. Migration patterns of the Pacific subpopulation of Japanese common squid Todarodes pacificus, estiamated from tagging experiments. Bull. Hokkaido Natl.
Fish. Res. Inst., 65: 21−43.
Mokrin, N.M., Y. Novikov and Y. Zuenko, 2002. Seasonal migra- tions and oceanographic conditions for concentration of the Jap- anese flying squid (Todarodes pacificus Steenstrup, 1880) in the northwestern Japan Sea. Bull. Mar. Sci., 71: 487−499.
Murata, M., 1978. The relation between mantle length and body weight of the Squid, Todarodes pacificus Steenstrup. Bull. Hok- kaido Reg. Fish. Res. Lab., 43: 3−51.
Murata, M., 1989. Population assessment, management and fishery forecasting for the Japanese common squid, Todarodes pacifi- cus. In: Marine Invertebrate Fisheries, edited by J.F. Caddy, Their Assessment and Management, Wiley, New York, pp.
613−636.
Murata, M., 1990. Ocean resources of squids. Mar. Behav. Physiol., 18: 19−71.
Murayama, T., Y. Hiyama and S. Kasahara, 1993. Why is autumn the main spawning season of the Common squid in the Japan Sea? Bull. Jap. Reg. Fish. Res. Lab., 43: 93−103.
Nagasawa, K., S. Takayanagi and T. Takami, 1993. Cephalopod tagging and marking in Japan: a review. In: Recent advace in Cephalopod fisheries biology, edited by Okutani, T., O'Dor, R.K. and T. Kubodera, Tokai University Press, pp. 331−333.
Nakamura, Y., 1985. Observation on periodical growth rings in the statoliths of common squid (Todarodes pacificus). Report of 1984 Annual Meeting on Resources and Fisheries of squids.
Hokkaido Reg. Fish. Res. Lab., 70−73. (in Japanese) Okutani, T., 1983. Todarodes pacificus. In: Cephalopod life cycles,
edited by Boyle, P.R., Vol 1. Academic Press, London, pp. 201− 216.
Osako, M. and M. Murata, 1983. Stock assessment of cephalopod resources in the Northwestern Pacific. In: Advances in Assess- ment of World Cephalopod Resources, edited by J.F. Caddy, FAO Fish. Tech. Paper., 231: pp. 55−144.