521 Corresponding author: [email protected]
✽장봉도 갯벌을 이용하는 대형 갑각류 군집의 계절 변화
서인수・홍재상
1✽
해양생태기술연구소,
1인하대학교 해양과학과
Temporal Variation of the Macro-crustacean Assemblages on Jangbong Tidal Flat, Incheon, Korea
In-Soo Seo and Jae-Sang Hong 1 *
Marine Eco-Technology Institute Co., Ltd., Busan 608-830, Korea
1 Department of Oceanography, Inha University, Incheon 402-751, Korea
This study examined the community structure and temporal variation of the macro-crustaceans on Jangbong tidal flat, Incheon, Korea. Macro-crustaceans on this tidal flat were collected using a modified otter trawl from November 1999 to January 2001. The macro-crustaceans comprised 32 species, 7,741 individuals, and 29,211.3 gWWt during the study period. The Jangbong tidal flat was dominated numerically by three shrimp (Palaemon carinicauda, Crangon affinis, and Metapenaeus joyneri) and two portunid crabs (Charybdis
japonica and Portunus trituberculatus). The macro-crustacean assemblages were divided into two different
season and species groups based on the q-mode and r-mode clustering methods. In terms of q-mode, they consisted of a winter species group from December through April and a summer species group from May through November. The r-mode analysis showed that the species group visiting the tidal flat in winter was dominated numerically by palaemonid and crangonid shrimps, while a species group characterized by an abundance of portunid crabs and penaeid shrimps was more frequent in spring, summer, and autumn.In conclusion, our study revealed distinct temporal variation in the tidal flat use by macro-crustaceans around Jangbong Island, Incheon, Korea.
Key words: Macro-crustaceans, Temporal variation, Community structure, Tidal flat, Jangbong Island
서 론
갯벌을 포함하는 연안역은 다양한 해양생물의 산란・성육 및 서식이 집중적으로 이루어지는 생태적으로 중요한 서식처 이다 (Potter et al., 1983; van der Veer et al., 1990; Santos and Nash, 1995; Nash and Santos, 1998; Fujita et al., 2002).
또한 다량의 유기물 유입과 높은 일차생산으로 인하여 서식 생물이 이용할 수 있는 먹이원이 풍부하다 (Levinton, 1995).
반면 환경적으로는 조석에 따른 주기적인 노출과 잠김현상 등의 변화가 크나, 조하대 지역과 비교하여 복잡하며 다양한 형태의 서식처가 있어 상대적으로 은신처로서 공간이 넓기 때문에 포식자로부터 포식 당할 위험이 적다는 장점이 있다 (McLusky, 1989). 일반적으로 해양생물의 분포는 자연적 또는 인위적인 환경요인에 의해 수시로 변화하며, 대형저서동물 가운데 이동성이 상대적으로 큰 어류 (저어류) 및 대형 갑각류 는 주야, 조석 및 계절에 따라 능동적으로 대처하는 습성을 가지게 되며, 인위적인 변화가 심하지 않은 지역에서도 시・공 간적인 변화가 큰 것으로 알려져 있다 (Lee, 1996).
한편 본 연구의 대상생물인 대형 갑각류의 분포에 관한 생물・생태학적인 연구는 연안역 환경요인의 가변성, 산란・보 육 및 성육장으로서의 중요성과 인위적인 환경변화의 정도를 파악하는데 매우 중요한 요소이다 (Szedlmayer and Able,
1996; Lazzari and Tupper, 2002). 지금까지 우리나라 연안역에 서 갑각류를 대상으로 한 연구는 공간적으로 내만과 하구역 등에 국한되어 왔다. 이 가운데 군집 연구로는 광양만 잘피군 락의 새우류와 게류 군집 (Huh and An, 1997a, 1997b)과 고리 주변해역 새우류 군집 (Huh and An, 1999)의 연구가 있다.
반면 개체군 연구로는 중하와 꽃새우의 생태 (Cha, 1997; Cha et al., 2002), 산모양깔깔새우의 성장 및 성 성숙 (Choi, 2001;
Choi et al., 2001), 민꽃게의 생태 및 종묘생산 (MOMAF, 1999) 과 꽃게의 자원생물학적 연구 (Yeon, 1997) 등이 있다. 본 연구는 섭식・생태학적 연구의 기초자료 생산을 위하여 갯벌 생태계의 중요 구성원인 대형 갑각류의 군집구조와 시간에 따른 변화양상을 파악하는데 목적이 있다.
재료 및 방법
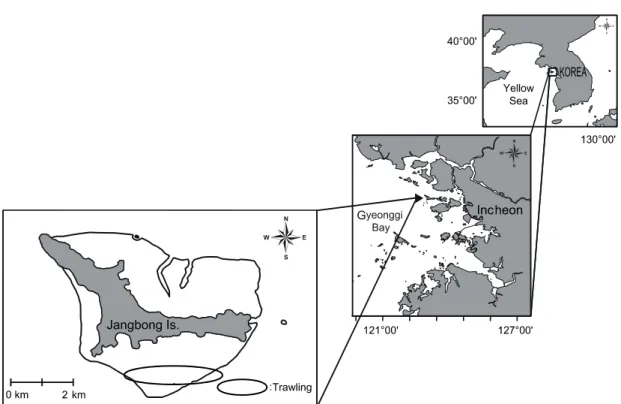
연구해역의 개황
한강 하구역에 위치한 장봉도는 섬을 기준으로 북쪽으로는
강화도와, 남쪽으로는 영종도와 접하여 있고, 인천항에서 북
서쪽으로 약 28.8 km 떨어져 있다. 해안선을 따라 남북으로
분포하는 갯벌의 총 면적은 약 19.0 km
2이다. 간조 시 노출되는
갯벌의 최대 길이는 서쪽의 한들갯벌이 약 2.0 km이었고, 동쪽
의 옹암갯벌이 1.2 km이었다. 방위상으로 서쪽에서 동쪽으로
향할수록 갯벌의 길이가 짧아지는 지형이다 (Fig. 1). 장봉도
Fig. 1. Location of the sampling sites on the tidal flat around Jangbong Island, Incheon, Korea.
해역의 조차는 대조차가 8.2 m, 소조차가 3.6 m로 평균 5.9 m의 조차를 보인다. 본 섬 남쪽해역의 장봉수로에서 밀물은 간조 시 북동으로 흐르며, 대조 시 최강유속이 104~206 cm/sec 에 이른다. 썰물은 만조 시 남서방향으로 흐르며, 만조 후 2시간 30분이 경과하면 최강유속이 90~170 cm/sec에 달한다.
한편 장봉도 인근해역은 해양생물을 먹이로 하는 각종 조류 (鳥類)가 서식하며, 1988년에는 노랑부리백로 및 괭이갈매기 의 번식지로 파악되어 천연기념물 제360호로 지정되었다. 또 한 2003년에는 국제적으로 보호가 요망되고 있는 저어새 등의 서식처로서 뿐만 아니라, 생물다양성이 뛰어나 습지보호지역 으로 지정・고시되는 등 생태학적으로 매우 가치가 높은 해역 이다 (국토해양부고시 제2008-722호). 그럼에도 불구하고, 1990년대부터 인천국제공항의 건설사업과 관련한 다양한 형 태의 개발사업이 이루어져 환경의 변화가 심한 해역으로 지적 되고 있다 (Lim et al., 1995).
Fig. 2. Sampling apparatus, small otter trawl used for the collection of fish assemblages on the tidal flat around Jangbong Island, Incheon, Korea.
현장조사
본 연구에서 대형 갑각류는 소형기선저인망 (오터트롤, small otter trawl)을 이용하여 1999년 11월부터 2001년 1월까지 총 15개월 동안 월별로 채집하였다 (Fig. 2). 대형 갑각류의 채집은 매월 사리 전후 2-3일내를 기준으로 간조 2시간 전에 실시하였다. 소형기선저인망은 목줄 (ground rope)이 20 m, 바닥줄 (bottom rope)이 16 m, 날개그물(wing net)이 좌우 8 m이었다. 당긴그물눈 (streched mesh size)은 날개그물과 몸통 그물이 공통적으로 20 mm를, 자루그물이 16 mm이었다. 그물 은 0.5 × 1 m 크기의 전개판에 의해 예인 시 그물이 벌어지게 된다. 한편 본 연구에서 대형 갑각류의 채집을 위한 소형기선 저인망의 예인시간과 채집면적은 Lee and Hwang (1995)의 결과에서 기술하고 있는 Lee (1991)의 적정채집법에 준하여 매 조사 시기 당 60분과 9,000 m
2로 통일하였다. 현장에서 채집된 대형 갑각류는 최종농도 10%인 중성포르말린으로 고 정한 후, 실험실로 운반하였다. 운반된 시료는 종 동정을 시행 한 후, 개체수를 계수하고, 생체량 (습중량, gWWt)을 측정하였 다.
환경요인의 측정은 생물 채집과 동시에 YSI 85를 이용하여 수온과 염분농도를 측정하였고, 강우량은 기상월보를 참고하 였다 (KMA, 1999-2001).
군집구조 분석
대형 갑각류의 군집구조를 파악하기 위하여, 풍부도 지수
(Margalef, 1958), 다양성 지수 (Shannon and Weaver, 1949)와
균등도 지수 (Pielou, 1975)를 구하였다. 또한 출현 종과 개체수
의 자료를 이용하여 Bray and Curtis (1957)의 유사도 지수 (similarity index)를 파악하였다. 유사도 지수 행렬로부터 각 조사 정점과 출현 종을 연결하는 방법으로는 group-average를 이용하였다. 군집분석은 PRIMER (Plymouth Routines Multivariate Ecological Research) computer package를 이용하 여 집괴분석 (cluster analysis)에 의한 수지도 (dendrogram)와 다차원배열법 (nMDS ordination)으로 표현하였다. 또한 군집 구조 분석에서 개체수는 우점종에 의한 효과를 줄이기 위하여 fourth-root로 변환하였다. 우점종 서열 분석을 위하여 LeBris index (1988)를 이용하였고, 그 식은 아래와 같다.
D'ij = Fij × Dij × 100
=(
× ) × (
) × 100
: j군집에서 i종의 출현 횟수
: j군집의 전체 정점수
: j군집에서 k번째 정점에 출현하는 i종의 개체수 또는 생체량
: k번째 정점의 전체 개체수
결 과
환경요인
조사 기간 중, 수온은 3.2℃~25.4℃의 범위에 월 평균 수온은 12.9℃이었다. 2000년 2월에 가장 낮았고, 동년 8월에 가장 높았다. 염분농도는 최소 26.2 psu (2월)에서 최대 31.0 psu (8월)의 범위에 평균 29.7 psu를 나타내었다. 강우량은 최소 1.5 mm에서 최대 591 mm의 범위에 수온도의 변화와 동일하 게 2000년 2월에 적었고, 8월에 높았다 (Fig. 3).
Fig. 3. Seasonal variation of water temperature, salinity and precipitation on the tidal flat around Jangbong Island, Incheon, Korea.
출현 종수, 개체수 및 생체량의 변화
조사 기간 중 총 32종, 7,741개체와 29,211.3 gWWt의 갑각류 가 채집되었다 (Table 1, 2). 분류군에 따른 출현 종수를 보면, 새우류 (Macrura)가 18종이 출현하여 56.2%를 점유하였고, 다음으로 게류 (Brachyura)가 11종으로 34.5%를 차지하였다.
이 외에 등각류 (Isopoda), 곤쟁이류 (Mysida)와 구각류 (Stomatopoda)가 공통적으로 1종이 출현하여 3.1%의 낮은 점 유율을 보였다. 조사 시기에 따른 출현 종수는 1~14종의 범위 에 매월 평균 8종 (±3종)이 출현하였다. 2000년 11월에 가장 다양성이 낮았고, 2000년 4월에 가장 높았다. 이 밖에 수온이 비교적 높은 시기에 해당하는 2000년 6월, 8월 및 9월에 10종 이상이 채집되어 상대적으로 다양하였다. 한편 생물다양성이 높았던 새우류는 1~12종의 범위에 매월 평균 5종 (±3종)이 출현하였다. 2000년 11월에 가장 적었고, 동년 4월에 가장 많았다. 이 외에 2000년 4월에 12종이 채집된 것을 제외하면, 기타 시기에는 상대적으로 적었다. 반면 게류는 2000년 7월부 터 9월까지 6~7종의 범위로 출현한 것을 제외하면, 이 밖의 시기에는 5종 이하로 출현하여 다양성이 낮았다 (Fig. 4).
개체수는 총 7,741개체가 채집된 가운데 새우류가 7,199개 체, 93.0%를 차지하여 가장 우점하였고, 다음으로 게류와 구각 류가 각각 363개체, 4.7%와 175개체, 2.3%를 점유하였다. 반면 곤쟁이류와 등각류는 불과 3개체와 1개체만이 채집되어 밀도 가 매우 적었다. 조사 시기별로는 5~2,814개체의 범위에 매월 평균 516개체 (±783개체)가 출현하였다. 생물다양성이 가장 낮았던 2000년 11월에 출현 개체수가 가장 적었던 반면, 동년 12월에 가장 많았다. 이 외에 가장 다양한 생물이 출현하였던 4월에 1,871개체가 채집되어 상대적으로 밀도가 높았다. 한편 개체수 점유율이 가장 높았던 새우류는 매월 평균 480개체 (±798개체)가 채집되었고, 2000년 11월에 적었으며, 동년 12 월에 많았다. 전반적으로 개체수의 변화 양상은 새우류의 밀 도 변화와 동일한 패턴을 나타내었다 (Fig. 4).
생체량은 단일 개체의 중량이 상대적으로 높은 게류가
15,290.8 gWWt을 나타내 전체의 52.3%를 점유하였고, 새우류
는 10,118.4 gWWt으로 34.6%를 차지하였다. 또한 구각류는
3,802.0 gWWt, 13.0%를, 등각류와 곤쟁이류는 공통적으로
0.1 gWWt 미만의 낮은 생체량을 보였다. 조사 시기에 따라서
는 10.2~5,280.5 gWWt의 범위에 매월 평균 1,947.4 gWWt
(±1,755.4 gWWt)을 나타내었고, 2000년 11월에 가장 낮았으
며, 동년 7월에 가장 높았다. 이 밖에도 2000년 8월, 9월 및
12월에 3,000.0 gWWt 이상의 생체량을 나타내 상대적으로
높았다. 주요 분류군별 생체량의 변화양상을 보면, 게류는
2000년 8월에 최대 4,164.6 gWWt을 기록하였고, 매월 평균
1,019.4 gWWt (±1,491.7)에 수온이 높은 6월부터 9월 사이에
생체량이 높았다. 새우류는 최소 10.2 gWWt (2000년 11월)에
서 최대 3,249.4 gWWt (2000년 12월)에 매월 평균 674.6 gWWt
(±899.4 gWWt)을 나타내었다. 이 외에 1999년 12월과 2000년
4월에 각각 1,245.2 gWWt과 2,194.3 gWWt의 높은 생체량을
보였다 (Fig. 4).
Table 1. Seasonal variation in abundance of macro-crustaceans collected by modified otter trawl on the tidal flat around Jangbong Island, Incheon, Korea (inds./9,000 m 2 )
Species name / Time 1999 2000 2001
Total Nov. Dec. Jan. Feb. Mar. Apr. May Jun. Jul. Aug. Sept. Oct. Nov. Dec. Jan.
Phylum Arthropoda Crustacea Brachyura
Atergatis reticulatus 1 1 2
Charybdis japonica 2 1 10 38 46 94 69 13 273
Matuta lunaris 3 1 4
Matuta planipes 1 1
Neodorippe japonica 3 2 5
Parthenope validus 2 9 2 1 14
Philyra pisum 1 1 1 3
Pugettia quadridens 1 2 1 4
Typhlocarcinops canaliculata 1 1
Xenophthalmus pinnotheres 1 1 2
Portunus trituberculatus 2 2 29 14 7 54
Macrura
Acetes japonicus 1 1
Alpheus japonicus 3 146 1 2 6 158
Alpheus rapax 1 3 1 5
Crangon affinis 22 522 36 8 57 997 24 10 3 436 46 2,161
Heptacarpus sp.1 1 2 71 1 75
Heptacarpus sp.2 1 1
Latreutes mucronatus 1 1 2 2 55 23 5 89
Leptochela gracilis 1 1
Metapenaeopsis dalei 2 12 40 54
Metapenaeus joyneri 3 8 62 53 20 141 18 305
Palaemon carinicauda 237 224 124 125 280 476 1 5 2,363 301 4,136
Palaemon gravieri 4 1 5 95 3 108
Palaemon macrodactylus 2 1 3 4 10
Palaemon modestus 1 1
Palaemon paucidens 2 2
Pandalus sp. 6 1 7
Penaeus orientalis 1 1 13 4 19
Trachysalambria curvirostris 1 2 6 52 5 66
Isopoda
Idotea sp. 1 1
Mysidacea
Neomysis orientalis 3 3
Stomatopoda
Oratosquilla oratoria 11 1 13 9 18 68 47 7 1 175
Total 276 755 165 138 350 1,871 68 180 213 184 320 43 5 2,814 359 7,741
Table 2. Seasonal variation in biomass of macro-crustaceans collected by modified otter trawl on the tidal flat around Jangbong Island, Incheon, Korea (gWWt/9,000 m 2 )
Species name / Time 1999 2000 2001
Total Nov. Dec. Jan. Feb. Mar. Apr. May Jun. Jul. Aug. Sept. Oct. Nov. Dec. Jan.
Phylum Arthropoda Crustacea Brachyura
Atergatis reticulatus 2.1 1.1 3.2
Charybdis japonica 87.1 74.8 388.4 1,644.1 1,803.2 2,549.4 2,171.1 1,058.5 9,776.6
Matuta lunaris 50.3 27.6 77.9
Matuta planipes 30.0 30.0
Neodorippe japonica 12.2 16.1 28.3
Parthenope validus 32.2 156.4 19.7 1.6 209.9
Philyra pisum 5.4 5.5 2.7 13.6
Pugettia quadridens 45.7 26.3 27.7 99.8
Typhlocarcinops canaliculata 0.4 0.4
Xenophthalmus pinnotheres 0.0
Portunus trituberculatus 597.3 382.7 1,548.2 1,516.5 1,006.4 5,051.1
Macrura
Acetes japonicus 0.2 0.2
Alpheus japonicus 2.3 177.3 0.7 3.1 6.2 189.6
Alpheus rapax 2.5 8.6 0.3 11.4
Crangon affinis 17.9 817.4 32.3 6.7 55.7 1,001.5 37.7 16.2 4.2 643.3 33.5 2,666.3
Heptacarpus sp.1 1.2 1.3 39.8 0.3 42.6
Heptacarpus sp.2 0.7 0.7
Latreutes mucronatus 0.4 0.1 0.3 0.2 6.8 8.0 0.7 16.5
Leptochela gracilis 0.2 0.2
Metapenaeopsis dalei 1.3 6.8 29.7 37.8
Metapenaeus joyneri 8.0 39.9 324.6 380.7 181.8 210.3 26.0 1,171.3
Palaemon carinicauda 416.6 421.2 184.6 191.8 358.9 831.6 2.5 10.2 2,596.3 419.2 5,433.0
Palaemon gravieri 3.9 0.4 3.3 107.7 1.5 117.0
Palaemon macrodactylus 2.2 0.7 1.2 2.2 6.2
Palaemon modestus 0.2 0.2
Palaemon paucidens 1.6 1.6
Pandalus sp. 16.9 0.4 17.3
Penaeus orientalis 28.5 28.5 145.7 148.4 351.1
Trachysalambria curvirostris 0.3 3.0 9.8 34.9 7.5 55.5
Isopoda
Idotea sp. <0.01 <0.01
Mysidacea
Neomysis orientalis 0.1 0.1
Stomatopoda
Oratosquilla oratoria 306.7 39.3 395.1 328.9 665.5 1,340.1 578.2 103.4 44.8 3,802.0
Total 836.7 1,405.1 218.8 202.5 453.4 2,589.6 1,430.4 3,134.1 5,280.5 4,925.5 3,729.6 1,289.3 10.2 3,249.4 456.6 29,211.3
Fig. 4. Seasonal variation of species number, abundance and biomass on the tidal flat around Jangbong Island, Incheon, Korea.
생태학적 제지수
생태학적 제 지수의 분석에서 단 1종이 출현한 2000년 11월 은 제외하였다. 종 풍부도는 0.8~2.1의 범위에 매월 평균 값은 1.2 (±0.5)이었다. 2000년 1월과 2월에 가장 낮았고, 동년 9월에 가장 높았다. 이 밖에 1999년과 2000년의 12월 (0.9)을 제외하 면 , 기타 시기에는 1.0 이상의 값을 보였다. 종 다양도는 최소 0.4 (2000년 2월)에서 최대 1.7 (2000년 5월과 6월)의 범위에 매월 평균 값은 1.0 (±0.6)이었다. 2000년 4월부터 11월까지는 1.0 이상의 수치를, 이 밖의 시기에는 1.0 이하로 낮았다. 균등 도는 0.2~0.8 (평균 0.5±0.2)의 범위에 2000년 5월과 10월에 가장 높았다. 전반적으로 수온이 낮은 시기에는 0.5 이하의 낮은 값을 나타내었다. 이와 같은 결과는 수온이 낮은 시기에 출현하는 자주새우 (Crangon affinis)와 밀새우 (Palaemon
carinicauda)의 대량 출현에 그 원인이 있었다 (Fig. 5).Fig. 5. Species richness, diversity and evenness of macro- crustacean assemblages on the tidal flat around Jangbong Island, Incheon, Korea.
Table 3. Dominant species ranking in abundance by LeBris index (1988) on the tidal flat around Jangbong Island, Incheon, Korea
Taxa Species name Total abundance Mean density
(inds.) % of total individuals Frequency of
occurrence LeBris index CDM Palaemon carinicauda 4,136 276±596 53.4 10 291,934.8 CDM Crangon affinis 2,161 144±287 27.9 11 119,711.1 CDB Charybdis japonica 273 18±30 3.5 8 57,298.6 CDM Metapenaeus joyneri 305 20±39 3.9 7 52,568.8 CST Oratosquilla oratoria 175 12±20 2.3 9 36,011.8 CDM Trachysalambria curvirostris 66 4±13 0.9 5 7,601.3 CDB Portunus trituberculatus 54 4±8 0.7 5 6,103.0 CDM Metapenaeopsis dalei 54 4±11 0.7 3 5,330.2 CDM Latreutes mucronatus 89 6±15 1.1 7 4,292.8 CDM Penaeus orientalis 19 1±3 0.2 4 2,736.2
Index; CDB, Arthropoda Crustacea Decapoda Brachyura; CDM, Arthropoda Crustacea Decapoda Macrura; CST, Arthropoda Crustacea Stomatopoda.
Table 4. Dominant species ranking in biomass by LeBris index (1988) on the tidal flat around Jangbong Island, Incheon, Korea
Taxa Species name Total biomass Mean biomass
(gWWt) % of total biomass Frequency of
occurrence LeBris index CST Oratosquilla oratoria 3,802.0 253.5±376.3 13.0 9 1,520,810.0 CDM Palaemon carinicauda 5,433.0 362.2±665.0 18.6 10 285,380.3 CDB Charybdis japonica 9,776.6 651.8±928.9 33.5 8 114,333.9 CDM Crangon affinis 2,666.3 177.8±340.0 9.1 11 78,074.1 CDB Portunus trituberculatus 5,051.1 336.7±567.7 17.3 5 31,346.6 CDM Metapenaeus joyneri 1,171.3 78.1±130.5 4.0 7 10,159.9 CDM Penaeus orientalis 351.1 23.4±51.2 1.2 4 3,256.6 CDM Alpheus japonicus 189.6 12.6±45.6 0.6 5 1,838.7 CDB Pugettia quadridens 99.8 6.7±14.4 0.3 3 1,283.5 CDM Palaemon gravieri 117.0 7.8±27.7 0.4 5 1,209.3
Index; CDB, Arthropoda Crustacea Decapoda Brachyura; CDM, Arthropoda Crustacea Decapoda Macrura; CST, Arthropoda
Crustacea Stomatopoda.
개체수 및 생체량에 따른 우점종
개체수와 생체량 및 그 출현빈도를 고려하여 우점종을 선정 하는 LeBris index (1988)를 이용하여 상위 10위까지의 우점종 을 선별하였다. 개체수에 있어서는 새우류가 7종으로 가장 많았고, 게류와 구각류는 각각 2종과 1종이 상위에 위치하였 다 (Table 3). 이들을 최우선 순위별로 나열해보면, 밀새우, 자주새우, 민꽃게 (Charybdis japonica), 중하 (Metapenaeus
joyneri), 갯가재 (Oratosquilla oratoria), 꽃게 (Portunus trituberculatus), 산모양깔깔새우 (Metapenaeopsis dalei), 남방넓적뿔꼬마새우 (Latreutes mucronatus) 및 대하 (Penaeus
orientalis) 등이었다. 이들 상위 10위종들이 차지하는 밀도는7,332개체로 전체의 94.7%를 점유하였다. 생체량에 있어서도 새우류가 6종으로 가장 많았고, 게류와 구각류는 각각 3종과 1종이 서열되었다. 그러나 최우선 순위에서는 갯가재가 1위에 위치하였고, 다음으로 밀새우, 민꽃게, 자주새우, 꽃게, 중하, 대하, 긴발딱총새우 (Alpheus japonicus), 뿔물맞이게 (Pugettia
quadridens) 및 그라비새우 (Palaemon gravieri) 순이었다(Table 4). 이들이 나타내는 생체량은 총 28,657.7 gWWt으로 전체의 98.1%를 차지하여 생체량의 변화 양상을 주도하는 것으로 파악되었다.
중요 수산자원의 계절변화
게류는 총 8과 11종이 출현하였고, 362개체가 채집되었다.
이 가운데, 꽃게과가 327개체의 밀도를 나타내 게류 출현 개체 수의 90.6%를 점유하였고, 기타 게류는 35개체, 9.4%를 차지 하였다. 특히 민꽃게와 꽃게는 각각 273개체와 54개체의 밀도 를 나타내 민꽃게의 출현비율이 상대적으로 높았다. 한편 이 들 2종의 계절변화 양상을 보면, 조사 전 기간에 걸쳐 공통적으 로 수온이 상승하는 2000년 4월부터 10월까지 갯벌에서 출현 하였고, 가장 높은 밀도를 보인 시기는 9월로 각각 141개체와 52개체가 채집되었다 (Fig. 6).
Fig. 6. Seasonal variation of Charybdis japonica and Portunus trituberculatus on the tidal flat around Jangbong Island, Incheon, Korea.
새우류는 8과 17종이 출현하였고, 7,199개체가 채집되었다.
주요 새우류 군집은 징거미새우과 (Palaemonidae, 5종), 보리 새우과 (Penaeidae, 4종) 및 자주새우과 (Crangonidae, 1종)등 이었다. 이 가운데 밀새우가 포함된 징거미새우과가 4,257개
체로 전체의 59.1%를 차지하였고, 자주새우과와 보리새우과 가 각각 2,161개체, 30.0%와 444개체, 6.2%의 점유율을 보였 다. 특히 단일 종으로 밀새우와 자주새우는 각각 4,136개체와 2,161개체가 채집되어 가장 밀도가 높았다. 한편 이들 주요 3과의 계절 변화 양상을 보면, 징거미새우과는 1~2,370개체의 범위에 매월 평균 287개체 (±599개체)가 출현하였다. 2000년 8월과 10월에 가장 적었고, 동년 12월에 가장 많았다. 전반적 으로 11월부터 이듬해 4월까지는 출현 밀도가 높았고, 그 밖의 시기에는 출현하지 않거나 또는 개체수가 매우 적었다. 반면 단일종으로 구성된 자주새우과는 3~997개체의 범위에 매월 평균 196개체 (±322개체)가 채집되었다. 1999년 12월, 2000년 4월과 12월에 400개체 이상의 높은 밀도를 나타내었고, 그 밖의 시기에는 50개체 이하로 적었다. 특히 하계인 8월부터 11월까지는 단 1개체도 출현하지 않았다. 반면 보리새우과는 최소 1개체 (2000년 4월)에서 최대 69개체 (2000년 6월)의 범위에 매월 평균 26개체 (±54개체)가 출현하였다. 수온이 낮은 시기에는 채집되지 않았고, 비교적 수온이 상승하는 2000년 4월부터 출현하기 시작하여 10월까지 채집되었다 (Fig. 7).
Fig. 7. Seasonal variation of Palaemonidae, Penaeidae and Crangonidae shrimps on the tidal flat around Jangbong Island, Incheon, Korea (upper, relative percent).
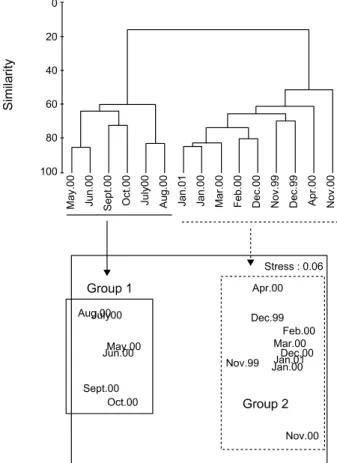
군집구조 분석
군집분석은 전체 출현 개체수의 0.1% 이상 (10개체 이상)을
점유하는 15종을 대상으로 실시하였다. 분석 결과, 정점 간
(q-mode) 및 종 간 (r-mode)에 따라 크게 2개의 그룹으로 대별
되었다. 정점 간 분석 결과, 그룹 1은 2000년 5월부터 10월까지
로, 그룹 2는 이를 제외한 시기로 구성되었다. 한편 R-mode
분석결과에서도 이를 잘 반영하여 그룹 1은 중하, 꽃새우,
대하, 산모양깔깔새우와 꽃게, 민꽃게, 자게 (Parthenope
validus) 및 갯가재가 포함되었다. 반면 그룹 2는 밀새우, 그라
비새우, 붉은줄참새우 (Palaemon macrodactylus), 자주새우, 남방넓적뿔꼬마새우, 긴발딱총새우 및 좁은뿔꼬마새우류 (Heptacarpus sp.1) 등이었다. 이와 같은 결과는 다차원배열법 (nMDS)에서도 동일하였다 (Fig. 8).
Fig. 8. Dendrogram for hierarchical clustering and 2-dimensional MDS configuration, using group-average linking of Bray- Curtis similarities calculated on fourth-root transformed abundance data.
고 찰
갯벌을 이용하는 대형 갑각류 군집의 생물량 (출현 종수, 개체수 및 생체량) 및 계절 변화
갯벌의 다양한 생태학적 기능들이 제시되었음에도 불구하 고, 이를 뒷받침할만한 연구는 매우 미약한 수준이라고 할 수 있다. 특히 지금까지 내만 및 연안역을 제외한 갯벌 및 그 인근해역 생태계에 서식하는 대형 갑각류 군집의 구조와 계절변화 양상을 동시에 밝힌 연구는 없었다. 갯벌을 대상으 로 대형 갑각류에 대한 연구는 거의 전무하다. 우리나라에서 수행된 대형 갑각류에 대한 연구는 광양만 잘피군락에서 수행 된 게류와 새우류 군집 및 고리 주변해역에서 출현한 새우류 군집의 계절변동에 관한 연구가 있을 뿐이다 (Huh and An, 1997a, 1997b, 1999).
본 연구 결과, 총 32종, 7,741개체와 29,211.3 gWWt의 대형 갑각류가 채집되었다. 출현 종수와 개체수에 있어서 새우류와 게류의 2개 생물군은 각각 90.6%와 97.7%를 차지하였고, 생체 량에 있어서는 이들 2개의 분류군과 함께 구각류가 99.9%를 점유하였다. 본 연구에서 출현한 대형 갑각류 군집의 생물량 (출현 종수, 개체수 및 생체량)은 비교적 생물다양성이나 생물 량이 높다고 알려진 해초지 (잘피) 군락의 37종과 비교하여 다소 적은 수준이었다 (Huh and An, 1997a, 1997b). 반면 해초 지 군락에서 출현한 개체수와 생체량은 각각 14,608개체와 27,537.4 gWWt을 나타내 본 연구와 비교하여 개체수는 약 2배 적었던 반면, 생체량은 다소 높거나 유사한 수준으로 파악 되었다. 그러나 해초지 군락에서의 채집면적이 본 연구와 비 교하여 상대적으로 컸음을 감안한다면, 생물다양성과 생체량 은 거의 유사하거나 많았고, 개체수는 적은 것으로 추정할 수 있다. 한편 새우류 군집만을 대상으로 고리 주변해역과 비교하면 (Huh and An, 1999), 생물다양성은 본 연구에서 17종 이 출현하여 고리 주변해역의 11종과 비교하여 많았다. 그러 나 개체수는 본 연구와 고리 주변해역에서 각각 7,199개체와 161,573개체가 채집되어 매우 큰 차이를 보였다. 이러한 출현 밀도는 채집면적의 차이에서 기인한 원인도 있겠지만, 일부 종의 대량 출현이 보다 크게 작용하였다. 즉 고리 주변해역의 경우, 자주새우류가 총 154,528개체가 출현하여 전체 출현 밀도의 95.6%를 점유하고 있기 때문이다.
일반적으로 해양생물 군집의 계절변화를 발현시키는 직접 적인 원인은 수온으로 알려져 있고, 개별 종에 따라서는 생활 사의 일부 혹은 전부에서 분포패턴에 차이를 발생시킨다 (Demestre et al., 2000). 이러한 이유로 수온이 상승하는 봄철에 외해에서 월동한 해양생물이 내만, 하구역 및 갯벌 등의 천해 연안역으로 이동하고, 그 결과 생물다양성 및 생물량 (개체수 및 생체량)이 증가하는 것이 일반적인 현상이다 (Lee and Seok, 1984; Kwak, 1997; Seo, 2003). 본 연구에서도 출현 종수 및 생물량은 수온이 낮은 겨울철에 가장 낮았고, 늦은 봄부터 늦가을에 상대적으로 높은 수치를 나타내었다. 이와 같은 결 과는 어류, 새우류 및 게류 군집의 계절변동을 밝힌 연구 결과 와 동일하였고, 결국 계절변동은 수온의 변화양상과 밀접한 연관이 있다는 것을 입증해 주고 있다 (Huh and An, 1997a, 1997b, 1999).
한편 우점종에 있어서도, 밀새우 및 자주새우를 포함한 상 위 10위 종들이 차지하는 밀도가 총 7,332개체로 전체 출현 개체수의 94.7%를 점유하고 있었다. 이렇듯 일부 종들의 우점 현상은 남해안 및 동해안과 비교하여 본 연구지역이 속한 서해안의 독특한 해양학적 특성의 반영으로 추정할 수 있다.
즉 서해안은 남해안 및 동해안과 비교하여 조석간만의 차가
상대적으로 클 뿐만 아니라, 해역의 물리적인 환경변화 요인
도 매우 다양하고 많다. 결국 환경요인의 변화가 심한 해역에
서는 환경에 적응한 일부 소수종이 우점하는 경향이 높다는
결과와 부합된다고 볼 수 있고, 이러한 현상은 갑각류를 포함
한 다른 해양생물에게서도 서해안의 만이나 연안에서 흔히
관찰되는 결과로 본 연구와 유사하였다 (Lee, 1989; Lee and
Kim, 1992; Lee and Hwang, 1995).
갯벌을 이용하는 대형 갑각류 군집의 계절 분할 전략 갯벌 생태계와 직・간접적으로 관련을 맺는 생물들은 일생 동안 일시적으로 갯벌을 이용하거나 혹은 생활사의 전 부분을 보내는 2가지의 행동양태를 보인다. 즉 여기에서는 생물이 가지는 고유의 특성인 유영능력과 물리적인 환경 특성을 바탕 으로 한 먹이원 (food sources)의 접근에서 그 원인이 비롯된다 고 하겠다. 이와 관련하여 Reise (1985)는 갯벌에서 먹이가 되는 생물 개체군 (prey populations)에 미치는 포식자 영향 (predator effects)을 갯벌의 상・중부 해역에서는 새들 (birds)에 의한 하향포식자(top-down predator)로, 중・하부에서는 어류 나 대형 갑각류들에 의한 상향포식자 (bottom-up pradtor)로의 강도(强度)가 더욱 중요하다는 가설을 제시하기도 하였다.
따라서 포식자 및 피식자 간의 섭식상호관계는 해역의 생물다 양성 및 생물량의 차이를 유발할 가능성이 높다. 다시 부연하 면, 계절에 따른 대형 갑각류의 출현 양태는 먹이자원이 풍부 한 해역에서 머물 수 있는 능력과 생활사의 어느 일부분에서 최적의 서식처를 선택할 수 있는 능력에 지배된다는 것이다 (Lasiak, 1984, 1986). 특히 우리나라와 같은 온대기후대에서는 수온의 변화가 계절에 따라 크게 다르기 때문에, 갯벌을 이용 하는 서식 생물들도 산란, 보육 및 성장을 위한 월동의 행동양 태를 명확하게 나타낸다. 따라서 갯벌에서 산란을 하는 종들 은 산란 이후, 여름과 가을에 먹이를 찾기 위한 회유를 하고, 수온이 하강하는 겨울에 월동장으로 향하게 되며 이러한 원인 으로 계절에 따른 차이가 발생하게 된다 (Jin and Tang, 1996).
본 연구에서도 새우류의 경우에 자주새우와 밀새우는 수온이 낮은 동계에 우점하였던 반면, 보리새우과의 새우류는 하계에 주로 높은 밀도를 나타내어 차이가 있었다. 또한 꽃게과의 게류에서도 이러한 현상은 동일하였다. 따라서 본 연구에서 대형 갑각류 군집은 크게 동계와 하계 특징종군으로 구분되고 있었다. 이 가운데 1999년 11월, 12월, 2000년 4월, 12월 및 2001년 1월에 높은 밀도를 보이는 밀새우와 자주새우는 냉수 기 특징종 (cold-water specialist)으로 나타낼 수 있었던 반면, 민꽃게, 꽃게와 중하 등은 2000년 5월부터 10월 사이의 시기에 우점하는 특성을 나타내 난수기 특징종 (warm-water specialist)으로 구분할 수 있었다.
한편 본 연구에서 특이한 사항은 자주새우의 포란 개체 출현의 시기와 2개과 새우류의 생태학적 지위에 관련된 것이 다. 첫째, 자주새우는 2000년 4월 및 12월에 포란한 개체가 동시에 출현하는 것으로 보아 갯벌을 명확하게 산란장으로 이용하고 있음을 추정할 수 있었다. 다만 그 포란 시기가 동계 와 춘계로 뚜렷하게 구분되는 것이 특징적이었다. 낙동강 하 구역에 서식하는 자주새우의 개체군 연구를 보면, 본 종은 거의 연중 포란하고 있었고, 추계인 9월과 춘계인 4월에는 포란율이 60.0% 이상으로 높아 본 연구결과와 유사한 경향을 보였다 (Hong and Oh, 1989). 따라서 생물이 서식하는 해역의 물리적인 조건은 상이하나, 이들의 생태적인 특성상 갯벌에서 의 연중 포란 개체의 출현은 일반적인 현상으로 추정할 수
있다. 둘째, 새우류 자원을 다양한 형태로 잘 이용해 왔던 우리나라의 경우, 징거미새우과와 보리새우과에 대한 생태학 적 지위에 관한 연구가 향후 보다 필요할 것으로 생각된다.
Everett and Ruiz (1993)는 동일 해역에서 Palaemonetes pugio라 는 종이 보다 대형 크기의 새우류인 Penaeus setifer가 상대적 으로 적었을 때 출현 밀도가 높았고, 특히 2종이 동일해역에서 공통적 또는 동시적으로 우점하는 경우에는 서로 중복되는 서식처를 가지지 않으려는 행동양태를 보인다고 제시한 바 있다. 즉 P. pugio는 보다 큰 포식자에 의해 점령된 해역에서 상대적으로 보다 떨어진 해역으로 그들의 생활장을 옮긴다는 점이다. 결국 중・소형 크기의 P. pugio가 다른 종간의 상호작용 과 요인의 조합에 의해 생물・생태학적인 반응을 보이고, 특별 히 자신보다 큰 생물 (P. setifer)이 분포하는 해역을 의도적으 로 피하는 경향이 있음을 제안하였다. 위의 결과로 볼 때, 본 연구해역에서도 징거미새우과와 보리새우과는 이러한 생 물 상호간의 작용에 의해 출현 빈도와 밀도를 달리한다는 가설을 제기할 수 있다. 특히 본 연구에서 이들 2과는 한개 과의 군집이 출현하면 다른 과는 전혀 채집되지 않는 특징으 로 매우 뚜렷하게 시기를 달리하였다. 그러나 동일해역에서 수행된 건간망 자료를 분석한 결과에 의하면 (Seo and Hong, 2006), 밀새우는 연중 갯벌 해역을 활발하게 이용하는 종으로 이미 확인된 바 있다. 따라서 밀새우는 보리새우과가 출현하 는 하계에 갯벌의 상・중부로 서식장을 옮겨서 분포하였음을 입증한다고 볼 수 있다. 결국 2개 과의 새우류가 나타내는 공간분포의 차이는 먹이원과 그에 따른 서식공간에 따른 경쟁 의 결과로 추정할 수 있다. Seo (2003)에 의하면, 본 연구해역에 서 출현한 보리새우과의 중하와 징거미새우과의 밀새우는 공통적으로 연체동물류, 갑각류 및 갯지렁이류 등 다양한 대 형저서동물을 먹이원하고 있었고, 다른 많은 연구들에서도 잡식성의 먹이습성을 나타내는 동일한 결과를 제시하고 있기 때문이다 (Cartes, 1995; Dittel et al., 1997; Collins and Paggi, 1998).
사 사
본 연구는 인하대학교의 지원에 의하여 연구되었으며, 이 에 감사드립니다.
참고문헌