Korean J Vet Res(2017) 57(1) : 1~7 https://doi.org/10.14405/kjvr.2017.57.1.1
1
<Original Article>
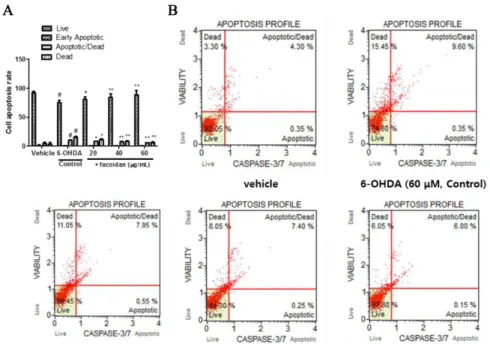
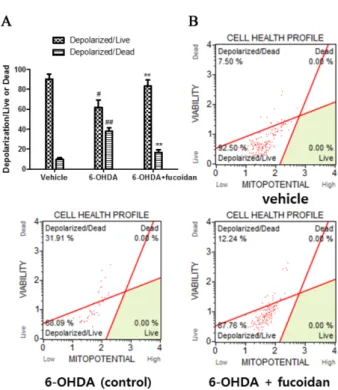
Fucoidan attenuates 6-hydroxydopamine-induced neurotoxicity by exerting anti-oxidative and anti-apoptotic actions in SH-SY5Y cells
Myung-Hwan Kim
1, Hoon Namgoong
1, Bae-Dong Jung
1, Myung-Sang Kwon
1, Yeon-Shik Choi
2, Taekyun Shin
3, Hyoung-Chun Kim
4, Myung-Bok Wie
1,*
1
College of Veterinary Medicine and Institute of Veterinary Science, Kangwon National University, Chuncheon 24341, Korea
2
Department of Laboratory Animal Science, Korea Biopolytechnic College, Nonsan 32943, Korea
3
College of Veterinary Medicine, Jeju National University, Jeju 63243, Korea
4