관련 문서
The porous PCL scaffolds produced from the gas foaming and salt leaching and plasma surface treatment are suitable for potential applications in bone
Here, we found that TAMR-MCF-7 cells had undergone EMT, evidenced by mesenchymal-like cell shape, down-regulation of the basal E-cadherin expression
Determining optimal surface roughness of TiO₂blasted titanium implant material for attachment, proliferation and differentiation of cells derived from
The differences found in these concepts of human resources and HRD originate from their diverse subjects and objectives. In the narrow definition of the concept, the subjects of
Conclusions: Thus, 25-HC induced odontoclast differentiation through the odontoclastogenesis factor-mediated activation of NF-κB and upregulated inflammatory
differentiation, integration (line, surface and volume differentiation, integration (line, surface and volume integrals).. del( ) operator, gradient, divergence and
We investigated the effects of BIX01294 on cellular senescence in human BM-MSCs (hBM-MSCs), and identified that an optimal treatment of BIX01294 leads to attenuated
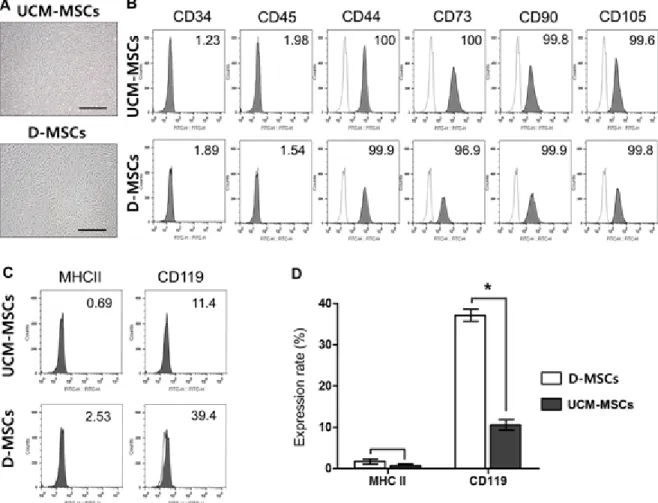
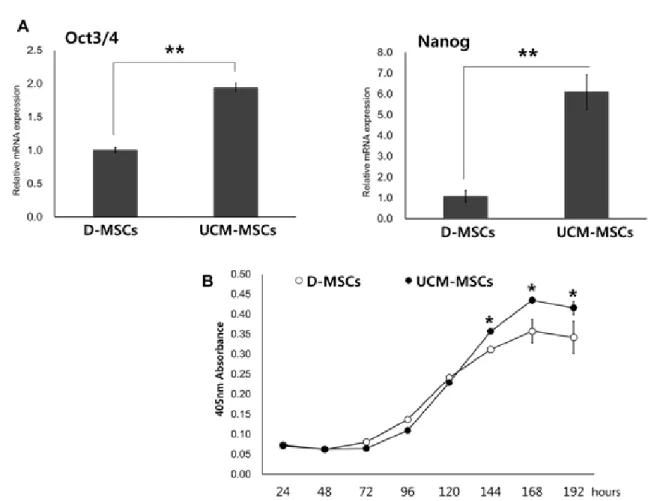
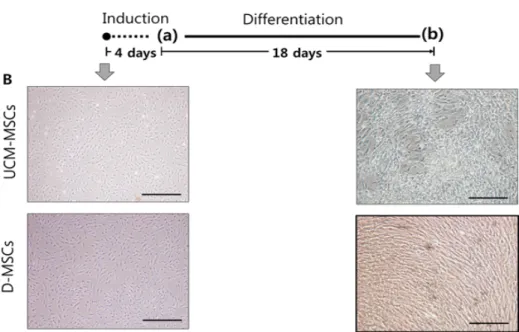
Meterials and Methods: Our study involved three stem cell sources-subacromial bursal tissue, bone marrow, and umbilical cord-derived stem cells. Under consent we