Candida fermentati SI의 exo-β-(1,3)-glucanase 유전자의 클로닝 및 그 특성
임유미1, 김봉기2, 김상준3, 소재현1*, 김원찬2*
1한약진흥재단
2경북대학교응용생명과학부
3해군사관학교이학과
Received: April 15, 2016 / Revised: May 19, 2016 / Accepted: May 29, 2016
서 론
Isoflavone은 lignan류와함께식물에서발견되는 phyto- estrogen으로본래 estrogen과비슷한생리활성을갖거나장 내균총에 의해 생리활성이 강화되는 phytochemical이다.
Isoflavone의종류는크게 4가지형으로분류가가능한데먼 저, aglycone 형태인 daidzien, genistein, glycitein과이들 의배당체형인 daidzin, genistin 및 glycitin이있으며, 이들 배당체에 acetyl과 malonyl이각각결합한형태등모두 12종 이존재한다[11]. 대두에존재하는 isoflavone aglycone의직 접적인작용에의한항암[9], 항산화[7], 항돌연변이작용[1]
등이보고되고있으며, 이들의화학적구조가여성 hormone 인 estrogen과유사하기때문에 phytoestrogen으로서의간 접적인효과에의한항유방암[8, 9], 항갱년기장애[12] 등 에대한효과도보고되고있다. 대두에는이러한 isoflavone 류가건조중량당약 1.5−2.5% 정도함유되어있으며, 이들 중 isoflavone 배당체의 일종인 daidzin, genistin이 전체 isoflavone의 60−70%로 가장 많은 부분을 차지하고있다 Molecular Cloning and Functional Expression of Extracellular Exo-β-(1,3)-Glucanase from Candida fermentati SI
Yu-Mi Lim1, Bong-Ki Kim2, Sang-Jun Kim3, Jai-Hyun So1*, and Won-Chan Kim2*
1National Development Institute of Korean Medicine, Gyeongbuk 38540, Republic of Korea
2School of Applied Biosciences, Kyungpook National University, Daegu 41566, Republic of Korea
3Department of Natural Science, Naval Academy, Changwon 51704, Republic of Korea
An isoflavone glucosidase that catalyzes the hydrolysis of isoflavone glucosides into glucose and corre- sponding aglycones was purified from Candida fermentati SI. The N-terminal sequence was determined to be GLNCDYCN. We designed degenerate primers on the basis of these amino acid sequences and success- fully cloned the full structural gene sequence of the isoflavone glucosidase using inverse PCR. The exo-β- (1,3)-glucanase gene consists of 1227 base-pair nucleotides, encoding a 408-amino-acid sequence that shares 41–96% amino acid homology with other yeast exo-β-(1,3)-glucanases belonging to glycoside hydrolase fam- ily 5. The recombinant exo-β-(1,3)-glucanase was expressed in Pichia pastoris X-33, using a pPICZA vector system, and further characterized. The molecular mass of the purified exo-β-(1,3)-glucanase was estimated by SDS-PAGE to be 47 kDa. The optimal pH and temperature were pH 4.5 and 40°C, respectively. The Km
values of the purified exo-β-(1,3)-glucanase for daidzin and genistin were 0.12 mM and 0.14 mM, respec- tively. The Vmax values of the purified isoflavone glucosidase were 945.03 U/mg for daidzin and 835.92 U/mg and for genistin.
Keywords: Isoflavone, Candida fermentati, exo-β-(1,3)-glucanase, isoflavone glucosidase
*Corresponding authors J.-H. S.
Tel: +82-53-810-0396, Fax: +82-53-810-0279 E-mail: [email protected]
W.-C. K.
Tel: +82-53-950-7765, Fax: +82-53-953-7233 E-mail: [email protected]
© 2016, The Korean Society for Microbiology and Biotechnology
[11]. 이에비하여배당체형태보다생리활성이다양하고높 은기능성을갖는 aglycone 형태인 daidzein, genistein 함량 은각각의배당체의약 10−30분의 1에불과한것으로알려 져있다[11]. 생체이용률에있어서는 aglycone인 genistein
을섭취한경우와배당체인 genistin을섭취한경우에이들
의체내대사속도의차이가실험동물과인체실험에서공히 genistein이더빨리흡수되는현상을보여 aglycone이생체 이용률이 우수함이 알려져 있다[2, 3, 8, 9, 14]. 따라서 isoflavone 배당체를 aglycone 형태로전환시키는능력이우 수한균주의선별이필요하며, 그러한능력이우수한균주 C. fermentati SI를된장으로부터분리보고한바있다[5]. 본 연구에서는이균주가생산하는 isoflavone 배당체가수분해 효소의생산에관여하는유전자를클로닝하여염기서열을 밝힌뒤 P. pastoris X-33에형질전환하여재조합효소의과 발현을통해 isoflavone aglycone의효율적인생산을시도하 였고, 재조합 isoflavone 가수분해효소의효소학적특성을 조사하였다.
재료 및 방법
균주의 선발 및 사용된 plasmid
실험에사용된 C. fermentati SI는경북일대에서수집한 재래식 된장에서 직접 분리한 균주로서, 내염성이 높고 isoflavone glucosidase를생산하는능력이우수한균주이 다. Escherichia coli TOP10F'는 plasmid의조작, 보존 및 추출을위하여사용되었으며, P. pastoris X-33은재조합단 백질의고발현을위한 host로서사용하였다. P. pastoris X- 33을이용한재조합단백질의고발현을위한 plasmid로서 pPICZA plasmid (Invitrogen, USA)를사용하였다.
DNA 염기서열 및 아미노산 서열의 분석
C. fermentati SI로부터정제된단백질은한국기초과학지 원연구원에의뢰하여 N-말단의아미노산지기의서열을결 정하였고, DNA 염기서열의결정은 Bioneer사(Korea)에의 뢰하여분석하였다. 분석된단편을이용하여미지의염기서 열을찾기위해서 inverse PCR을통한 primer walking 법 을사용하였다. 염기서열이결정된 DNA 단편의결과로부터 각말단의염기서열을기초로양방향 primer를제작하였으
며, inverse PCR에 의한 양방향의 염기서열의 분석은
Bioneer사(Korea)에의뢰하여분석하였다. 염기서열결과에 대한 open reading frames (ORFs)의 검색은 National Center for Biotechnology Information (NCBI)서운영하는 ORF finder (http://www.ncbi.nlm. nih.gov/gorf.html)를사 용하여조사하였다. DNA의염기서열의분석및단백질아 미노산서열의상동성의검색은 BLAST (http://www.ncbi.
nlm.nih.gov. BLAST)를이용하였다.
Pichia pastoris X-33의 형질전환
P. pastoris X-33의 형질전환을 위한 competent cell은 YPD 배지에서 OD600 ≒1.3−1.5가될때까지배양원심집균 한균체를 0℃로냉각된멸균 증류수로 2 차례세척한후, 0℃로냉각된 1 M sorbitol 다시한번세척한후, 원심집균 하고이를 1 M sorbitol 용액에현탁하여 P. pastoris X-33의 competent cell로사용하였다. 준비된 competent cell을 C.
fermentati SI의 exo-β-1,3-glucanase gene이삽입되어있는 plasmid와 섞은 후 electroporator (Bio-Rad, Gene pulser X-cell)를이용하여 electroporation을실시하였다.
형질전환주의 단백질 발현 유도
형질전환 된 P. pastoris X-33의 발현을 위하여 먼저 BMGY 배지에 exo-1,3-β-glucanase 유전자가 삽입된 P.
pastoris X-33을접종후 30℃에서 24시간배양한뒤원심분 리하여, 균체를취한후새로운 BMMY 배지에현탁하였다. 그후 24시간마다 methanol을 0.5%가되게첨가하여효소 의생산을유도하다. 유도된재조합단백질의정제는 Ni-NTA Agarose resin(Qiagen, Germany)을이용하여제조사의추 천방법에따라정제하였다.
단백질 정량과 전기영동
단백질의 정량은 UV spectrophotometer를 이용하여
Bradford 법에 준하여측정하였다. 단백질의순도 검정은
sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE)는 Laemmli [6]의방법에준하여수행하였다.
SDS-PAGE 상에서 분자량측정을 위한 표준 단백질로는
pagerulertm prestained protein ladder (Fermentas, USA) 를사용하였다.
정제효소의 특성
정제된효소의특성을알아보기위하여 100 mM citrate phosphate buffer (pH 4.5) 2.7 ml에 효소액 100 μl와 10 mM p-nitrophenyl-β-D-glycosides 종류의 각 기질을 200μl 첨가하여 37℃에서 30분간반응시킨후 1 M sodium carbonate를 1 ml 첨가하여효소반응을정지시켰다. 이때생 성된 p-nitrophenol을 A420에서흡광도를측정후미리작성 된검량식을이용하여효소활성을측정하였다. 본논문에서 는기질로부터 1분간 1 μmol의 p-nitrophenol을생산하는데 사용된효소의양을 1 U (μmol/min)로정의하였다. 또한천 연기질(laminarin, amygdalin, esculin, cellobiose, salicin)을 이용한효소활성측정은 glucose assay kit (Sigma, USA)를 이용하였다. 각 10 mM 기질 100 μl와정제된효소 100 μl를
800 μl의 100 mM citrate phosphate buffer (pH 4.5)와섞 은후 37℃에서 30분간반응시킨뒤, 100℃에서 2분간끓임 으로써효소반응을정지하였다. 각반응액을 1/10로희석하 여 200 μl를 만든 후 glucose assay kit의 assay reagent (glucose oxidase 500 U, peroxidase 100 U) 400 μl와 37℃ 에서 30분간반응시킨뒤 12 N H2SO4 400 μl를가하여반 응정지후 A540nm에서흡광도를측정하여 glucose standard
solution으로미리작성한검량식을이용하여효소의활성을
측정하였다. 본 논문에서는 기질로부터 1분간 1 μmol의 glucose를생산하는데사용된효소의양을 1 U (μmol/min) 로정의하였다.
HPLC 분석
HPLC를 이용하여 genistin, daidzin, genistein 및 daidzein을정량하였다. 이때 사용된 column은 Nova-Pak (Waters, 3.9 × 150 mm)이고, mobile phase는 1% acetic acid를함유한 H2O와 1% acetic acid를함유한 methanol (이 때 methanol은 40분동안 10%에서 60%까지증가하도록농 도구배를주었다)이다. Flow rate는 0.8 ml/min이며검출을 UV detector로 UV254 nm에서검출하였다.
결과 및 고찰
Isoflavone 배당체 가수분해효소의 유전자 클로닝
C. fermentati SI 유래의단백질로부터결정된 8개의 N 말 단아미노산잔기(GLNCDYCN)의서열을 NCBI의 BLAST 프로그램 (http://www.ncbi.nlm.nih.gov /BLAST/)을이용하 여 alignment를실시한결과 Meyerozyma guilliermondii의 exo-1,3-beta glucanase의 N-말단의배열순서(GLNWDYDN)와 가장유사함을확인할수있었다. 분석을통하여 N 말단아 미노산잔기의서열을확인하였으나그정확성이확보되지
않은상태여서 NCBI 검색을통하여얻은정보와종합하여
degenerate primer를제작하고 PCR을수행하였다. 정방 향 primer의제작을위하여 8개의 N 말단 아미노산서열 을 이용하여 degenerated primer GF1를 제작하였다(5'- GGIYTIAAYTGGGAYTAYGAYAA-3', I: inosine, Y: c or t).
역방향 primer의제작을위해서 glycosyl hydrolase family 5에속하는유전자들이가지는보전적인부위의아미노산서
열을다른 family와비교한결과가장보존적인아미노산서
열은 IGIEKKNEPL로확인되었으며, 이부위의아미노산서 열을기준으로 degeneration을고려하여역방향 primer GR2 을제작하였다(5'-TCTGTCAAAGCAGCAGACCATTCACC-3').
합성한 GF1 및 GR2 primer로 degenerated PCR을이용하 여얻은 841 bp의 DNA 단편의 정보(Fig. S1)를이용하여 inverse PCR을실시하였다. DNA 단편의내부를절단하지
않는제한효소(EcoR I)를이용하여 chromosomal DNA를처 리한후 T4 ligase를이용하여 ligation을시키고동일한말 단으로인해 self ligation된 DNA를미리제작해둔 primer를 이용하여 PCR을통해증폭을실시한결과 3,160 bp의 DNA 단편을얻었다(Fig. S2). 이 DNA 단편의염기서열을분석한 결과 1227 bp의 exo-β-1,3-glucanase 구조유전자를가지고 있음을확인하였다(Fig. 1).
염기서열이 결정된 DNA 단편의 정보 분석
염기서열이결정된 DNA 단편내에존재하는 ORF를확 인하기위하여 NCBI에서운영하는 ORF finder program을 이용하여최종아미노산서열을분석하였다. Expasy에서운 영하는 Compute pI/Mw (http://web.expasy.org/compute_pi/) 프로그램으로추정한결과분자량은약 50.4 kDa으로추정 되었다. 또한 glycoside hydrolase family 5에포함된다른 종의 yeast 유래 exo-1,3-β-glucanase와의 homology 비교를 하였을 때 각 41–96%의 homology를 나타내었고, 그 중 Meyerozyma guilliermondii ATCC 6260의 exo-1,3-β-glucanase 와 96%로가장높은 homology를나타내었다. 또한 N 말단 아미노산분석과비교한결과 19개의아미노산잔기로된 signal peptide가존재함을확인하였다.
Exo-β-1,3-glucanase의 고발현
P. pastoris X-33을통해 exo-β-1,3-glucanase의과발현을 실시하였다. 제한효소 EcoR I과 KpnI 부착자리가첨가된정 방향 및 역방향 primer를 제작하여 C. fermentati SI의 chlomosomal DNA를주형으로하여 PCR을수행한후 exo- β-1,3-glucanase 구조유전자를증폭하였다. 이증폭된 PCR 산물을정제후제한효소 EcoR I과 KpnI로처리하고미리 EcoR I과 KpnI로처리된 pPICZA에삽입하였다. Saccharomyces cerivisiae와같이선형화된 DNA는전체 genome내에있는 homologous한 영역과 재조합 DNA사이에서 homogolous
recombination이일어남으로써안전한형질전환체를얻을
수있다고알려져있다. 마찬가지의방법으로재조합 plasmid 상에있는 alcohol oxidase 영역과 genome내에있는 alcohol oxidase 영역사이에서 homologous recombination이일으켜 서 exo-β-1,3-glucanase 유전자를 P. pastoris X-33 genome 내로삽입시켰다. Alcohol oxidase의 promoter 하류영역에 삽입된 exo-β-1,3-glucanase 유전자의발현을위해 24시간마 다 methanol 0.5%가되도록첨가하면서 exo-β-1,3-glucanase의 발현을유도하였다. 이렇게발현시킨 exo-β-1,3-glucanase를 Ni-NTA Agarose resin (Qiagen, Germany)을이용하여정제 한후전기영동을실시한결과약 47 kDa 부위에서단일밴 드를얻을수있었다(Fig. 2). 추정치와실제전기영동상의분 자량의차이는 signal peptide에기인하는것으로생각된다.
exo-1,3-β-glucanase의 활성과 안전성에 미치는 pH의 영향 정제된 exo-1,3-β-glucanase의활성에미치는 pH의영향 을 조사하기 위해서 pH 3.0−pH 7.0은 100 mM citrate phosphate buffer, pH 6.5−pH 8.0은 100 mM sodium
phosphate buffer, pH 7.5−9.0은 100 mM Tris-HCl buffer 를이용하여 pH 3.0에서 pH 9.0 사이의 buffer를만든후, 37℃ 에서 30분간반응시킨뒤 p-nitrophenyl-β-D-glucopyranoside를 기질로사용하여효소의활성을측정하였다. 그결과 Fig. 3A Fig. 1. Nucleotide sequence and deduced amino acid sequence of exo-β-(1,3)-glucanase gene in Candida fermentati SI. The deduced amino acid sequences are shown below the nucleotide sequences and start or stop codon for exo-β-(1,3)-glucanase are indicated in bold.
와같이정제된 exo-β-1,3-glucanase의최적 pH는 100 mM citrate phosphate buffer가사용된 pH 4.5 부근이었다. 또한
효소의 pH 안정성을조사하기위해서정제된효소액에 pH 3.0−9.0 사이에서위와동일완충액을첨가하여각 pH별로 효소를 4℃에서 24시간방치한후효소의최적 pH에서 37℃ 로 30분반응후효소의잔존활성을측정하였다. 그결과 Fig.
3B와같이정제된 exo-β-1,3-glucanase는비교적넓은영역 의 pH 4.5−8.0 사이에서 80% 이상의활성을유지하는안정 한효소였다.
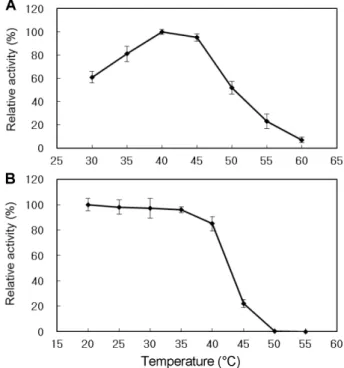
exo-1,3-β-glucanase의 활성과 안전성에 미치는 온도의 영향 효소활성에미치는온도의영향을조사하기위해서효소 의최적 pH 4.5 및 30−60℃의온도범위에서 5℃간격으로 각온도에서 30분간처리시킨효소의활성을측정하였다. 그 결과, Fig. 4A와같이정제된 exo-1,3-β-glucanase의최적온 도는 40℃였으며, 45℃이상에서는효소활성이급격히떨어 졌다. 효소의열안정성을조사하기위해서정제된효소를 최적 pH 4.5 및 30−60℃온도범위에서 5℃간격으로각온 도변화를주면서 30분간방치한후최적온도인 40℃에서 30분간반응시켜효소의잔존활성을측정하였다. 그결과, Fig.
4B와같이 exo-1,3-β-glucanase는 40℃까지는 80% 이상의 효소활성을유지하였고, 그이후에는효소활성이급격하게 Fig. 2. SDS-PAGE analysis of the purified recombinant exo-
1,3-β-glucanase from Pichia pastoris X-33. Molecular weight of recombinant exo-1,3-β-glucanase was determined to be 47 kDa. M, molecular size marker; 1, purified recombinant exo- 1,3-β-glucanase.
Fig. 3. Effect of pH on exo-β-(1,3)-glucanase activity (A) and stability (B). ◆-◆, 100 mM citrate phosphate buffer; ■-■, 100 mM sodium phosphate buffer; ▲-▲, 100 mM Tris-HCl buffer.
Fig. 4. Effect of temperature on exo-β-(1,3)-glucanase activity (A) and stability (B). For the optimal temperature, the enzyme activity was measured in the standard reaction mixture at the indicated temperature for 30 min at pH 4.5. For the thermal stability, the enzyme was incubated at optimal pH for 30 min at the indicated temperature. The remaining activity was measured in the standard reaction mixture for 30 min at 45℃.
떨어졌다.
효소의 기질특이성
정제된효소의기질특이성을확인하기위하여합성기질및 천연기질에대하여효소의기질특이성을조사하였다. 그결 과합성기질의 경우ρ-Nitrophenyl-β-D-glucopyranoside에 대해서 1.12 U (100%)로가장 높은활성을나타내었으며, 천연기질의경우 laminarin에대하여 5.40 U (100%)로서 가장 높은 활성을 나타내었고 amygdalin, esculin 및 cellobiose에대해서각각 3.40 U (63%), 0.16 U (3%), 0.11 U
(2%)의활성을나타내었다. 하지만 salicin에대해서는거의 활성을나타내지않았다(Table 1).
효소반응속도
정제된재조합 exo-β-1,3-glucanase의 isoflavone 배당체에 대한 효소반응속도를 알아보기 위해 여러 가지 농도의 daidzin과 genistin을 각 10 μl와 정제된 효소 50 μl를 440μl의 100 mM citrate phosphate buffer (pH 4.5)에첨 가하여 37℃에서 30분간반응후 500 μl의 ethyl acetate를 첨가하여 isoflavone을추출한뒤 ethyl acetate층을회수하 여 methanol을적당량섞은후 10 μl를 HPLC 분석에이용 하였다. 그결과정제된재조합 exo-β-1,3-glucanase Daidzin 에대한 Km값은 0.12 mM, Vmax는 945.03 U/mg protein이 었으며, genistin에 대한 Km 값은 0.14 mM, Vmax는 835.92 U/mg protein으로나타났다(Fig. 5). 이는기존에보 고된 Thermotoga maritima [13], soybean [10], almod emulsion [4] 유래의β-glucosidase 등에비하여낮은 Km값 과높은 Vmax를나타낸것으로, 재조합 exo-β-1,3-glucanase는 발효공학을통한 isoflavone 배당체의분해에이용할수있 는우수한효소라고생각된다.
요 약
본연구에서는 C. fermentati SI가생산하는 isoflavone 배 당체가수분해효소를클로닝하여염기서열을밝힌뒤 P.
pastoris X-33에형질전환하여재조합효소의과발현을시켰 고, 또한재조합 isoflavone 가수분해효소의효소학적특성 을조사하였다. 재조합 isoflavone 가수분해효소의분자량 은약 50.4 kDa이었으며, Meyerozyma guilliermondii ATCC 6260의 exo-1,3-β-glucanase와 96%로가장높은 homology 를나타내었다. exo-1,3-β-glucanase의 ORF는 pPICZA 벡터 로클로닝 후 P. pastoris X-33으로형질전환을 하였으며, His6-tag을 이용하여 효소를 정제하였다. 정제된 효소는 citrate phosphate buffer pH 4.5에서최적활성을나타내었 으며, 효소의최적활성온도는 40℃로나타났다. 40℃이상 Table 1. Substrate specificity for the purified recombinant exo-β-1,3-glucanase.
Chromogenic substrates Natural substrates
Substrate name Relative activity (%) Substrate name Relative activity (%)
ρ-Nitrophenyl-β-D-glucopyranoside 100 Laminarin 100
ρ-Nitrophenyl-β-D-xylopyranoside 13 Amygdalin 63
ρ-Nitrophenyl-β-D-galactoopyranoside 9 Esculin 3
ρ-Nitrophenyl-β-D-cellobiopyranoside 3 Cellobiose 2
o-Nitrophenyl-β-D-glucopyranoside 18 Salicin 0
o-Nitrophenyl-β-D-galactoopyranoside 1
Fig. 5. Lineweaver-Burk plot of the purified exo-β-1,3-gluca- nase for daidzin (A) and genistin (B).
에서는효소의활성이급격하게감소함을확인하였으며, pH 안정성을조사한결과비교적넓은범위인 4−8 사이에서 80%
이상의활성을유지하였다. 따라서, 재조합효소의과발현을 통해 isoflavone aglycone의효율적인생산에이용할수있 을것으로사료된다.
References
1. Akiyama T, Ishida J, Nakagawa S, Ogawara H, Watanabe S, Itoh N, et al. 1989. Genistein, a specific inhibitor of tyrosine protein kinases. J. Biol. Chem. 262: 5592-5595.
2. Brandi ML. 1997. Natural and synthetic isoflavones in the pre- vention and treatment of chronic diseases. Calcif. Tissue. Int. 61:
S5-8.
3. Hutchins AM, Slavin JL, Lampe JW. 1995. Urinary isoflavonoid phytoestrogen and lignan excretion after consumption of fer- mented and unfermented soy products. J. Am. Dietetic Assoc. 95:
545-551.
4. James R, Ketudat C, Asim E. 2010. β-Glucosidases. Cell. Mol. Life Sci. 67: 3389-3405.
5. Kim WC, So JH, Kim SI, Shin JH, Song KS, Yu CB, et al. 2009. Isola- tion, identification, and characterization of Pichia guilliermondii K123-1 and Candida fermentati SI, producing isoflavone β-glyco- sidase Korean traditional soybean paste. J. Appl. Biol. Chem. 52:
163-169.
6. Laemmli UK. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685.
7. Lee JH, Kim MH, Im SS. 1991. Antioxidative Materials in Domestic
Meju and Doenjang. 1. Lipid Oxidation and Browning during Fermentation of Meju and Doenjang. J. Korean Soc. Food Nutr.
20: 148-155.
8. Pagliacci MC, Amacchia M, Migliorati G, Grignani F, Riccardi C, Nicoletti I. 1994. Growth-inhibitory effects of the natural phy- toestrogen genistein in MCF-7 human breast cancer cells. Eur. J.
Cancer 30: 1675-1682.
9. Peterson G, Barnes S. 1991. genistein inhibition of the growth of human breast cancer cells: Independence from estrogen recep- tors and the multi-drug resistance gene. Biochem. Biophysic. Res.
Comm. 179: 661-667.
10. Suzuki H, Takakashi S, Watanabe R, Fukushima Y, Fujita N, Nogu- chi A, et al. 2006. An isoflavone conjugate-hydrolyzing β-gluco- sidase from the roots of soybean (Glycine max) seedlings. J. biol.
chem. 281: 30251-30259.
11. Wang HJ, Murphy PA. 1994. Isoflavone composition of American and Japanese soybean in Iowa: effects of variety, crop year, and location. J. Agric. Food. Chem. 42: 1647-1677.
12. Won HJ, Lee BS, Lee SK, Cjoi Y, Yoon S, Park KH, et al. 2001. The effect of isoflavone on postmenopausal symptoms and hor- monal changes in postmenopausal women. J. Korean Soc. Meno- pause 7: 54-63.
13. Xue Y, Yu J, Song X. 2009. Hydrolysis of soy isoflavone glycosids by recombinant β-glucosidase from hyperthrmophile Thermo- toga maritima. J. Ind. Microbiol. Biotechnol. 36: 1401-1408.
14. Yamaguchi M, Gao YH. 1998. Anabolic effect of genistein and genistin on bone metabolism in the femoral-metaphyseal tis- sues of elderly rats: the genistein effect is enhanced by zinc. Mol.
Cell. Biochem. 178: 377-382.