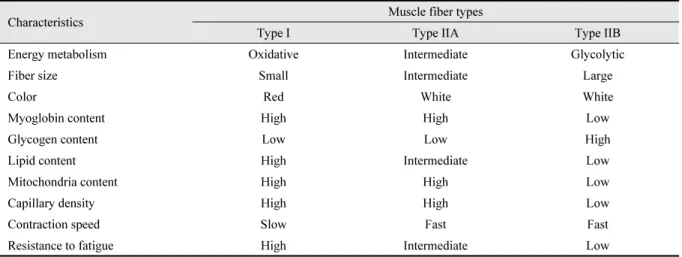
Overview of muscle metabolism, muscle fiber characteristics, and meat quality
8
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
(7)
(8)
수치
관련 문서
• Junction between motor neuron and muscle fiber. – Motor
1 John Owen, Justification by Faith Alone, in The Works of John Owen, ed. John Bolt, trans. Scott Clark, "Do This and Live: Christ's Active Obedience as the
Effects of Nitroglycerin(Ni) on Spontaneous Activity(S.A) and Oxytocin (OT) Induced Contractions in the Uterine Smooth Muscle. Data are expressed as mean
In the change of physical strength, The exercise group showed statistically significant differences in muscle strength(right), muscle strength(left),
The third was combination of levator palpebrae, superior rectus, inferior oblique muscle weakness(n=5) and the fourth was the third group plus medial rectus
In the change of functional fitness, muscle strength and endurance, cardiopulmonary endurance, wits, and flexibility showed significantly increased results,
The improvement factors of the performance showed significant increase in muscle strength, muscle endurance, cardiovascular endurance, power, agility and
In the health fitness category of the adult women who underwent instrument pilates exercise, they were statistically significant in muscle strength,
