http://dx.doi.org/10.12925/jkocs.2017.34.3.613
대사에너지가 열 스트레스에 노출된 오리의 간, 십이지장 융모, 미생물, 유전자 조절에 미치는 영향
신종서․양부근
✝․박병성
✝ 강원대학교 동물생명과학대학(2017년 8월 13일 접수: 2017년 8월 27일 수정: 2017년 9월 5일 채택)
Influence of metabolizable energy on histology of liver and duodenal villus, microflora, heat shock protein gene in duck under heat stress
Jong-Suh Shin․Boo-Keun Yang✝․Byung-Sung Park✝
College of Animal Life Science, Kangwon National University, Chuncheonsi Gangwondo, 200-701 Republic of Korea
(Received August 13, 2017; Revised August 27, 2017; Accepted September 5, 2017)
요 약 : 본 연구는 열 스트레스 하에서 오리사료 내 대사에너지(ME) 수준이 오리의 간, 십이지장 융 모, 미생물, 유전자 조절에 미치는 영향을 조사하였다. 총 240마리의 육용 오리 채리밸리(
Anas platyrhynchos
)를 4처리구로 완전임의배치 한 후 42일 동안 사육하였다. 처리구는 ME 2900 kcal/kg, ME 3000 kcal/kg, ME 3100 kcal/kg 및 ME 3200 kcal/kg로 구분하였다. 간 조직은 처리구 사이의 차이가 없었고, 십이지장 융모 및 창자샘 길이는 ME 3000과 비교할 때 2900은 10.58% 감소하였으나 3100, 3200과의 사이에 차이는 없었다. 맹장Latobacillus
는 ME 3000과 비교할 때 2900은 9.47% 감 소하였으나 3100, 3200은 각각 2.52, 3.24% 증가하였다. Total aerobic bacteria,E. coli
, Coliform bacteria는 ME 3000과 비교할 때 2900은 증가하였으나 3100, 3200은 차이가 나타나지 않았다. 간에서 HSP (heat shock proteins)-mRNA 중 HSP 90-α는 ME 3000과 비교할 때 2900은 48.60% 감소하였 으며 3100, 3200은 차이가 없거나 증가하였다.주제어 : 오리, 열 스트레스, 대사에너지, 융모, 미생물, 유전자
Abstract : The object of this study was to determine the influence of dietary metabolic energy (ME) on ... A total of 240 meat ducks Cherry valley (
Anas platyrhynchos
) were assigned into four treatment groups with a randomized block design for 42 days. The four treatments were: ME 2900 kcal/kg, ME 3000 kcal/kg, ME 3100 kcal/kg, and ME 3200 kcal/kg. There was no difference in liver tissue among the treatments. The duodenal villi and crypt depth length decreased by 10.58% in 2900 compared with ME 3000, but there was no difference between 3100 and 3200.Counts of caecal
Latobacillus
decreased by 9.47% in 2900 compared to ME 3000, but increased by
✝
Corresponding author
(E-mail: bkyang@kangwon.ac.kr, bspark@kangwon.ac.kr)
2.52 and 3.24% in 3100 and 3200, respectively. Total aerobic bacteria,
E. coli
and Coliform bacteria were increased by 2900 when compared to ME 3000, but there was no difference between 3100 and 3200. HSP 90-α among the heat shock proteins (HSPs)-mRNA in the liver was reduced by 48.60% in 2900 compared to ME 3000, while 3100 and 3200 showed no difference or increased.Keywords : Duck, heat stress, metabolizable energy, vilus, microorganism, gene expression
1. 서 론
기후 변화로 인한 지구온난화가 가속화되면서 육용오리의 열 스트레스 피해가 커지고 있으며 양계농가 소득감소의 원인이 된다[1]. 포유류와 달리 피부에 땀샘이 거의 없는 오리는 적정 체온 을 유지하기 어렵고 온몸이 깃털로 덮여있어 열 스트레스에 취약하다. 환경온도가 30℃ 이상 증 가하면 체온을 유지하기 위하여 헐떡거림을 통한 알칼리성 호흡증이 발생하고 환경온도가 증가할 수록 횟수가 증가하여 폐사에 이르게 된다. 오리 가 열 스트레스에 노출되면 열 충격 단백질(Heat shock protein, HSP)과 관련한 생체 유전자 조절 이 작동하여 열 스트레스에 반응한다[2]. 열 스트 레스에 취약한 닭에서 HSP 70, HSP 60, HSP 40, 및 HSP 27 이 증가되었고[3] 거위에서 HSP 70의 발현 수준이 증가하는 것으로 보고되었다 [4]. 열 스트레스는 유해 산소 생성으로 지질과산 화를 일으켜 가금류에 대사장애, 세포손상과 같은 해로운 영향을 미친다[5]. 열 스트레스로 인한 생 리적 변화로써 사료섭취, 사료효율, 성장능력, 산 란율 감소 및 폐사율이 증가하여 가금 산업에 막 대한 경제적 손실이 발생한다[6, 7]. 최근 급속히 발전한 우리나라의 오리 산업은 그 성장세에 비 해 환경 변화 대응을 위한 기술 개발이 미흡하 다. 오리의 영양소 수준 및 사양관리 프로그램이 명확히 정립되어 있지 않아 기존에 연구가 많이 된 육계 자료에 의존하고 있다. 외국의 경우 다 양한 오리 품종에서 에너지 요구량 구명 연구가 수행된 바 있지만[8, 9], 우리나라와 서로 다른 기후, 환경 차이로 인하여 농가에서 실제로 외국 의 자료를 이용하기에는 한계가 있다. 과도한 에 너지 섭취로 인한 과잉 에너지는 가금의 체내에 지방을 축적시켜 생산성과 사료효율에 해로운 영 향을 미친다[10]. 비만으로 인하여 심부전과 생식 장애, 폐사율을 증가시키며 에너지 대사에 문제가
발생한다 [11, 12]. 대사에너지(Metabolic energy, ME)는 사료의 총 에너지로부터 분뇨 및 가스로 손실되는 에너지를 뺀 에너지 값을 말하 며 가금은 방광이 없고 분과 뇨가 총배설강을 통 하여 함께 배설되므로 총 에너지에서 분 에너지 를 뺀 가소화 에너지 개념보다는 ME 값을 이용 하고 있다. 가금이 소모하는 대사에너지의 약 75%는 체열로 전환되어 주위로 발산한다[13]. 외 부 환경 온도가 증가함에 따라 에너지 섭취량은 감소하는 반면[12] 음수의 섭취량이 급격하게 증 가하여 세포 내 삼투압의 변화를 일으켜 세포활 성에 부정적인 영향을 미친다[14]. 육계에서 사료 내 에너지 수준이 낮을 때 증체량이 감소되며 에 너지 수준을 증가 시킬수록 증체량이 증가했다는 결과[15] 및 산란계에서 에너지 수준이 증가할수 록 증체량이 개선되었다는 연구 결과가 있다[16].
체온상승에 영향을 미치지 않는 양질의 고에너지 사료와 과잉 에너지로 인한 비만을 피하기 위한 적정 ME 수준 등 다양한 연구가 필요하지만 혹 서기 열 스트레스에 노출된 육용오리에 대한 ME 수준의 연구결과는 거의 알려진 바 없다. Sim 등 (2017)은 열 스트레스에 노출된 육용오리에서 ME 수준이 혈액의 생화학적 변수에 영향하는 생 리적 항상성 메카니즘을 보고하였다[17]. 본 연구 는 혹서기 열 스트레스에 노출된 육용오리의 사 료 내 조단백질을 18%로 고정하고 ME 수준을 서로 다르게 조절하였을 때 간 기능, 십이지장 융모, 미생물, 유전자 조절에 미치는 영향을 조사 하였다.
2. 실 험
2.1. 실험설계
Cherry valley (
Anas platyrhynchos
) 육용오리 240수를 부화 당일 경기도 양평 소재의 부화장으Ingredient Starter, 1 to 21d
Finisher, 22 to 42 d
ME 2900 ME 3000 ME 3100 ME 3200
Moisture, % 10.60 11.10 11.10 11.10 11.10
Crude protein, % 23.00 18.00 18.00 18.00 18.00
Crude fat, % 6.13 8.06 8.26 8.39 8.53
Crude fiber, % 2.56 2.40 2.40 2.40 2.40
Crude ash, % 5.06 4.80 4.80 4.80 4.80
Calcium, % 0.87 0.83 0.83 0.83 0.83
Available P, % 0.5 0.46 0.46 0.46 0.46
Methionine, % 0.65 0.58 0.58 0.58 0.58
Lysine, % 1.41 1.12 1.12 1.12 1.12
ME1), kcal/kg 3000 2900 3000 3100 3200
1)Metabolizable energy.
Table 1. Chemical composition of the experimental diets 로부터 공급받아서 4처리×3반복(반복 당 20수)
으로 완전임의배치 한 후 42일 간 사육하였다.
처리군은 사료 내 조단백질 수준을 18%로 고정 하고 ME (kcal/ kg)의 수준을 각각 2900, 3000, 3100, 3200으로 조절하였다. 한국가금사양표준 [18]에 의하면 오리의 영양소 요구량이 조단백질 18%, ME 3000 kcal/kg이며 본 연구는 이를 바 탕으로 ME 수준을 조절하였다. 시험사료는 옥수 수, 대두박 위주의 일반 상업용 원료를 이용하였 으며 영양성분표는 Table 1과 같다. 왕겨를 깔집 으로 사용하였으며 사육실의 온도는 입추 당일로 부터 3일까지는 33℃로 유지하였고 그 다음 기간 부터 주당 2-3℃씩 낮췄다. 실험기간 동안 처리 군은 연속조명과 일반음수를 무제한 급여하였다.
전기(1-21일)에는 일반사료를 무제한 급여하였으 며 후기(22-42일)에는 하루 5시간(11:00-16:00) 씩 33-40℃의 열 부여와 함께 상대습도 70%로 조절하여 열 스트레스를 유도하였다. 동물실험은 유럽의 실험동물취급면허 교재에 제시된 규정을 준수하였으며[19] 강원대학교 동물생명과학대학 의 관행 사육방법에 따라서 진행하였다.
2.2. 간, 소장 융모의 임상병리학적 소견 간을 채취한 다음에 10% 중성 포르말린에 고 정하여 자동침투기를 이용하여 에탄올 70, 90, 95, 100%까지 단계별로 탈수시켰다. Xylene으로 투명과정을 거친 후 파라핀으로 포맷하였다. 조직 절편을 3-5 μm로 제작하였으며 Hematoxylin and Eosin (H&E)을 실시하여 슬라이드를 제작 후 광학현미경(Olympus, BX41, Japan)을 이용하 여 조직병리학적 검사를 진행하였다. 소장을 채취
한 다음에 10% 중성 포르말린에 고정하여 자동 침투기를 이용하여 에탄올 70, 90, 95, 100%까 지 단계별로 탈수시켰다. Xylene으로 투명과정을 거친 후 파라핀으로 포맷하였다. 조직 절편을 3-5 μm로 제작하였으며 Hematoxylin and Eosin (H&E) 염색을 실시하였다. 융모(Villus) 길이와 창자샘 길이(Crypt depth)를 측정하기 위 해 조직 슬라이드를 광학현미경(Olympus, BX41, Japan)으로 검경하였다. 하나의 시료 당 임의의 5개 융모 길이와 창자샘 두께를 측정한 후 그 평 균 값을 이용하여 각 시료의 융모 및 창자샘의 길이를 계산하였다. 조직 제작과정 중 발생 할 수 있는 손상된 형태의 융모들은 측정대상에 포 함시키지 않았다. 디지털 이미지 분석 프로그램인 IMT iSolution lite version 8.0 (IMT Inc., Vancouver, Canada)로 이용하여 40배율의 사진 을 촬영하였다.
2.3. 맹장 미생물
42일 사육이 종료된 오리를 경추탈골에 의하 여 안락사 시킨 후 얼음 위에서 맹장의 양쪽 끝 을 실로 묶어 혐기적 조건으로 채취하였다. 맹장 은 AnaeroGen sachets (Oxoid, Hampshire, UK) 가 갖춰진 Sealed anaerobic jars (Oxoid, Basingstoke, UK)에서 혐기적 상태로 유지하였
다. 맹장 내용물과 멸균된 생리식염수
(phosphorus buffered saline, PBS 0.1 M, pH 7.0)를 혼합하여 희석(1:9, wt/vol)한 후 단계별 희석을 진행하였다. 모든 절차는 anaerobic chamber (5% hydrogen, 5% CO2, balanced nitrogen)에서 혐기적 상태로 이루어졌다. 희석된
HSP 90-α
f
orward 5’-TATTACACCTCCGCATCTGG-3’
reverse 5’-CGGAAGCTCCAAACCCTCT-3’
HSP 70
forward 5’-CCCCCAGATCGAGGTTACTTT-3’
reverse 5’-CTCCCACCCGATCTCTGTTG-3’
HSP 60
forward 5’-CCTAAAGGGGAAGGGTGAA-3’
reverse 5’-ACATCACTCGTCCCTGCTACGCCAT-3’
HSP 40
forward 5’-CCTGTGAAACGACGAGCAT-3’
reverse 5’-CGACCACCTGGCATTTCT-3’
HSP 10
forward 5’-AGTTCCTTCCCCTGTTTGAT-3’
reverse 5’-GCTTGTAGCACTTTCCCTTGA-3’
β-actin
forward 5’-ATGTCGCCCTGGATTTCG-3’
reverse 5’-CACAGGACTCCATACCCAAGAA-3’
10-2-10-7에서 각각 100 uL를 분주하여 멸균 된 평판배지에 각각 분주하였다.
Lactobacillus
(MRS agar, Oxoid, Basingstoke, UK), Total aerobic bacteria (Nutrient agar, Difco),E.coli
(MaConkey purple agar, Difco), Coliform bacteria (Violet red bile agar, Difco)를 각각의 해당 배지를 이용하여 배양하였다.Lactobacillus
는 Sealed anaerobic jars를 이용한 혐기적 상태 에서 37℃에서 48시간 배양하였고 기타 미생물은 37℃에서 24시간 호기 배양하였다. 미생물 군락 의 수는 맹장내용물 g당 균수 (CFU, colony forming unit/g of fresh cecal content)로써 상용 로그를 취하여 제시하였다[20].2.4. 생체 유전자 발현
간 1-2 g을 채취하여 30 cm의 aluminum cane을 이용하여 액체질소에 급속 동결 후 -8 0℃ 에서 보관 후 분석용 시료로 이용하였다. 간 조직시료30 mg을 사용하여 Xprep Tissue RNA Mini Kit (Phile Korea Technology, PKT)로써 total RNA를 추출하였다. RNA 농도는 ND-1000 Spectrophotometer (NanoDrop, Wilmington, DE, USA)를 사용하여 260 nm 흡 광도에서 측정하였으며 300-500 ng/uL의 total RNA를 얻었다. 추출된 RNA 1 uL를 이용하여 cDNA synthesis kit (PKT)로서 1st strand cDNA를 합성하였다. cDNA (complementary DNA)는 70℃ 5분, 42℃ 30분, 85℃ 5분 배양하 여 증폭하여 RT-PCR에 의한 주형으로써 사용하 였다. RT PCR (real time-polymerase chain reaction, Illumina Inc. USA)의 프로토콜에 따라 서 heat shock protein의 유전자발현을 측정하였 다. RT-PCR은 QuantiMix SYBR Kit (PKT)를 이용하였으며 cDNA를 Rnase free water를 이용 하여 1/5로 희석한 후 각각의 목표 유전자 단편 을 혼합하여 수행하였다. target gene의 적절한 primers는 미국 국립생명공학정보센터(National center for biotechnology information, NCBI)의 gene bank data base 자료를 기초로 하여 오리 의 HSP 90-α, HSP 70, HSP 60, HSP 40, HSP 10에 대하여 (주)바이오니아에 제작, 의뢰하 였다. Target gene에 대한 mRNA의 상대적 발현 측정을 위한 house keeping gene으로서 β-actin 을 사용하였다.
2.5. 통계분석
모든 data는 IBM SPSS statistics[21]를 이용하 여 평균 값과 표준오차를 산출하였으며 처리군 사이의 통계적 유의차 검증을 위하여 분산분석을 실시하였다. Duncan's multiple range test를 이용 하여 95% 신뢰수준에서 유의차를 검정하였다 (p<0.05).
3. 결과 및 고찰
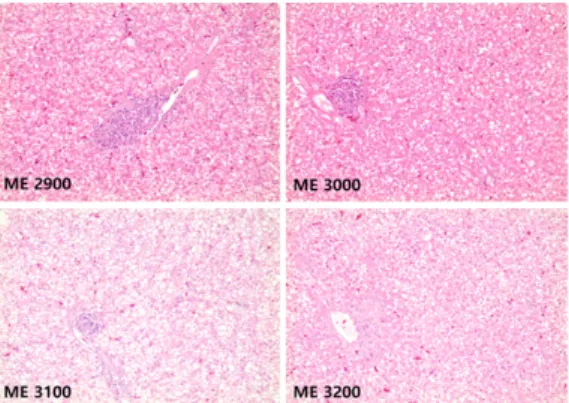
3.1. 간, 소장 융모의 임상병리학적 소견 열 스트레스에 노출된 육용오리에게 조단백질 을 18%로 고정하고 서로 다른 ME 수준의 사료 를 급여한 이후 측정한 소장 융모 및 간의 임상 병리학적 결과는 Table 2, Fig. 1, Fig. 2와 같다.
전체적으로 검사한 간장의 대부분에서 관찰된 소 견으로 중심정맥을 중심으로 방사상으로 배열되 어 있던 세포판이 붕괴됨으로써 동양모세혈관의 전형적인 모습을 찾아볼 수 없었다. 간세포 내 글리코겐 축적에 의한 공포화가 다양한 정도로 관찰되었지만, 이러한 간의 손상 정도는 ME 처 리군 사이에 큰 차이가 없었다. 십이지장 융모 및 창자샘 길이 비율은 ME 3000과 비교할 때 ME 2900은 10.58% 감소하였으나(p<0.05) ME3100, 3200과의 사이에 차이는 없었다. 혹서
ME (kcal/kg)
2900 3000 3100 3200
Villus height, ㎛ 980.4±8.91c1) 1210.7±37.91a 1176.3±18.21b 1186.7±11.34ab Crypt depth, ㎛ 236.8±9.13b 261.8±10.93a 256.9±10.62a 259.5±8.80a DVH/CD ratio 4.14±0.13b 4.63±0.21a 4.58±0.17a 4.58±0.12a
1)Mean values±standard errors (n=9). a,b,c Different letters represent significant differences between groups (p<0.05).
Table 2. Duodenal villus height (DVH), crypt depth (CD) and ratios of DVH to CD in meat duck under heat stress (22-42 days)
Fig. 1. Histology of the liver according to the levels of metabolizable energy (ME) in meat ducks under heat stress (22-42 days).
기 열 스트레스에 노출된 육계에서 융모와 창자 샘 길이의 비율은 소장에서 영양소의 분해와 흡 수에 대한 능력을 평가하는 기준이 될 수 있으며 이러한 비율이 증가할 때 영양소의 분해와 흡수 가 극대화 된다[22]. 그러나 혹서기 열 스트레스 하에서 오리 사료 내 ME 수준의 증가에 따른 보 고는 알려 진 바 거의 없다.
3.2. 맹장 미생물
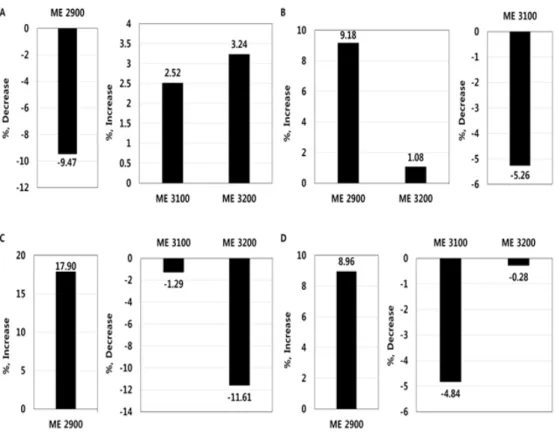
열 스트레스에 노출된 육용오리에게 조단백질 을 18%로 고정하고 서로 다른 ME 수준의 사료 를 급여한 이후 측정한 맹장 미생물은 Table 3 및 Fig. 3과 같다. 맹장 유익한 미생물
Latobacillus
는 ME 3000과 비교할 때 ME 2900 은 9.47% 감소하였으나 ME 3100, 3200은 각각 2.52, 3.24% 증가하였다(p<0.05). 장내 유해한 미생물 총호기성균은 ME 3000과 비교할 때 ME 2900은 9.18% 증가하였으나(p<0.05) ME 3100,3200 사이는 차이가 없었다.
E. coli
는 ME 3000 과 비교할 때 ME 2900은 17.90% 증가하였으나 ME 3100은 차이가 없었고 ME 3200은 11.61%감소하였다(p<0.05). 콜리폼균은 ME 3000과 비 교할 때 ME 2900은 8.96% 증가하였으나 ME 3100은 4.84% 감소하였고(p<0.05) ME 3200은
차이가 없었다. 장내 유익한 미생물인
Latobacillus
및 Bifidobacteria의 증식과 함께 유 해한E. coli
의 성장억제는 영양소 이용율을 증가 하여 줌으로써 사료효율과 성장율을 높이고 가축 의 생산성을 향상시키는 것으로 알려져 있다[23].Fig. 2. Photomicrograph of the duodenal villus height and crypt depth according to the levels of metabolizable energy (ME) in meat ducks under heat stress (22-42 days). The villi (blue arrow) are seen as finger-like projections.
Crypt (red arrow). (Haematoxylin–
eosin staining, ×40).
ME (kcal/kg)
2900 3000 3100 3200
Lactobacillus
7.55±0.20c1) 8.34±0.18b 8.55±0.27ab 8.61±0.24a Total aerobicbacteria 8.09±0.44a 7.41±0.43b 7.02±0.43b 7.49±0.44b
E. coli
7.31±0.38a 6.20±0.28b 6.12±0.27b 5.48±0.34cColiform bacteria 7.66±0.21a 7.03±0.17b 6.69±0.22c 7.01±0.18b
1)Mean values±standard errors (n=9). a,b,cDifferent letters represent significant differences between groups (p<0.05).
Table 3. Effects of different levels of metabolizable energy (ME) on cecal microflora in meat ducks under heat stress (22-42 days) (CFU log10/ fresh cecal content g)
Fig. 3. Changes in cecal microflora according to the levels of metabolizable energy (ME) in meat ducks under heat stress (22-42 days). Comparison of ME 3000 versus ME 2900, 3100 and 3200. A:
Lactobacillus
, B: Total aerobic bacteria, C:E. coli
, D:Coliform bacteria.
ME (kcal/kg)
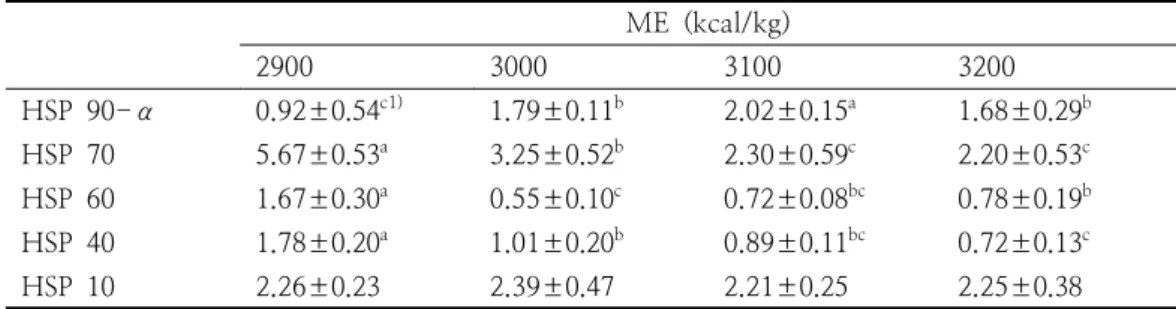
2900 3000 3100 3200
HSP 90-α 0.92±0.54c1) 1.79±0.11b 2.02±0.15a 1.68±0.29b HSP 70 5.67±0.53a 3.25±0.52b 2.30±0.59c 2.20±0.53c HSP 60 1.67±0.30a 0.55±0.10c 0.72±0.08bc 0.78±0.19b HSP 40 1.78±0.20a 1.01±0.20b 0.89±0.11bc 0.72±0.13c
HSP 10 2.26±0.23 2.39±0.47 2.21±0.25 2.25±0.38
1)Mean values±standard errors (n=9). a,b,cDifferent letters represent significant differences between groups (p<0.05).
Table 4. Effects of different levels of metabolizable energy (ME) on heat shock protein (HSP)-mRNA expression in meat ducks under heat stress (22-42 days)
Fig. 4. Changes in heat shock protein (HSP)-mRNA expression according to the levels of metabolizable energy (ME) in meat ducks under heat stress (22-42 days).
Comparison of ME 3000 versus ME 2900, 3100 and 3200. A: HSP 90-α, B:
HSP 70, C: HSP 60, D: HSP 40.
3.3. 생체 유전자 발현
열 스트레스에 노출된 육용오리에게 조단백질 을 18%로 고정하고 서로 다른 ME 수준의 사료 를 급여한 이후 측정한 간 스트레스 관련 유전자 발현은 Table 4 및 Fig. 4와 같다. 간에서 열 스 트레스와 관련된 HSP-mRNA 중 HSP 90-α는 ME 3000과 비교할 때 ME 2900은 48.60% 감
소하였으나 ME 3100은 12.85% 증가하였고 (p<0.05) ME 3200은 차이가 없었다. HSP 70은 ME 3000과 비교할 때 ME 2900은 74.46%가 증가하였으나 ME 3100, 3200은 각각 29.23, 32.31% 감소하였다(p<0.05). HSP 60은 ME 3000과 비교할 때 ME 2900, 3100, 3200은 각 각 203.6, 30.91, 41.82%가 증가하였다(p<0.05).
HSP 40은 ME 3000과 비교할 때 ME 2900은 76.24% 증가하였으나 ME 3100, 3200은 각각 11.88, 28.71% 감소하였다(p<0.05). HSP는 환경 적 스트레스, 특히 열 스트레스에 반응하여 합성 되는 특이 단백질이며 열 스트레스가 높아지면 HSP의 발현률이 증가하기 때문에 상대적으로 양 계의 스트레스 정도를 가늠할 수 있는 척도로 알 려졌다[24]. 열 스트레스에 노출된 오리에 대한 HSP 90-ɑ-mRNA 유전자 발현은 감소하지만 기타 HSP family 유전자의 발현은 증가하는 것 으로 알려졌다[25].
4. 결 론
본 연구결과는 여름철 열 스트레스에 노출된 오리의 후기(22-42일) 사료 내 단백질 수준을 18%로 고정하고 ME (kcal/kg) 수준을 3000 이 상으로 증가해주면 간 세포의 정상적인 발육을 유지함과 동시에 소장 융모와 창자샘의 길이를 개선시키는 것으로 나타났다. 맹장 내 유익한 미 생물과 유해 세균의 균총을 적절하게 유지해주며 생체 유전자 조절에 의해 열스트레스를 완화시키 는 것으로 나타났다.
1. 간장의 손상 정도는 ME 처리군 사이에 큰 차이가 없었다.
2. 십이지장 융모 및 창자샘 길이 비율은 ME 3000과 비교할 때 ME 2900은 감소하였으 나 3100, 3200은 증가 또는 차이가 없었다.
3. 맹장
Latobacillus
는 ME 3000과 비교할 때 ME 2900은 감소하였으나 3100, 3200은 증 가 또는 차이가 없었다.4. 총호기성균,
E. coli
, 콜리폼균은 ME 3100 과 비교할 때 ME 2900은 증가하였으나 3100, 3200은 감소 또는 차이가 없었다.5. 간에서 열 스트레스 단백질 HSP-mRNA 중 HSP 90-α는 ME 3000과 비교할 때 ME 2900은 감소하였으나 3100, 3200은 증가하였으며, 기타 유전자 발현은 서로 다 른 발현 양상을 나타냈다.
결론적으로 이러한 작용 메카니즘에 의해 오리 의 열 스트레스 저감효과를 갖는다고 판단된다.
감사의 글
본 연구는 농촌진흥청 2016년 아젠다사업 (C1010613-01-03) 지원으로 연구하였으며 이에 감사드립니다.
References
1. L. Tomanek, Variation in the heat shock response and its implication for predicting the effect of global climate change on species' biogeographical distribution ranges and metabolic costs,
J. Exp. Biol
, 213, 971-979 (2010)2. F. Di Domenico, R. Sultana, C. F. Tiu, N.
N. Scheff, M. Perluigi, C. Cini, D. A.
Butterfield, Protein levels of heat shock proteins 27, 32, 60, 70, 90 and thioredoxin-1 in amnestic mild cognitive impairment: an investigation on the role of cellular stress response in the progression of alzheimer disease,
Brain.
Res
, 1333, 72-81 (2010)3. J. Yu, E. Bao, J. Yan, L. Lei, Expression and localization of Hsps in the heart and blood vessel of heat-stressed broilers,
Cell Stress and Chaperones
, 13, 327 –335 (2008)4. B. W. Wang, X. P. Wu, X. H. Zhang, X.
H. Jia, M. A. Zhang, F. Y. Long, Z. G.
Yang, L. Wang, Expression and purification of goose HSP70 and compound formation with virus polypeptide,
Agr. Sci. China
, 7, 239-247 (2008)5. K. Sahin, O. Kucuk, Heat stress and dietary vitamin supplementation of poultry diets, Nutrition Abstracts and Reviews.
Series B:
Livestock Feeds and Feeding
, 1, 41–50 (2001)6. M. M. Mashaly, G. L. Hendricks, M. A.
Kalama, A. E. Gehad, A. O. Abbas, P. H.
Patterson, Effect of heat stress on production parameters and immune responses of commercial laying hens,
Poult.
Sci
, 83, 889-894 (2004)7. M. U. Sohail, M. E. Hume, J. A. Byrd, D.
J. Nisbet, A. Ijaz, A. Sohail, M. Z.
Shabbir, H. Rehman, Effect of supplementation of prebiotic mannan oligosaccharides and probiotic mixture on growth performance of broilers subjected to chronic heat stress,
Poult. Sci
, 91, 2235-2240 (2012)8. H. P. Fan, M. Xie, W. W. Wang, S. S.
Hou, W. Huang, Effects of dietary energy on growth performance and carcass quality of white growing Pekin ducks from two to six weeks of age,
Poult. Sci,
87, 1162-1164 (2008)9. M. Xie, Y. Guo, T. Zhang, S. Hou, W.
Huang, Lysine requirement of male white pekin ducklings from seven to twenty-one days of age,
Asian-Aust. J. Anim. Sci
, 22, 1386-1390 (2009)10. P. M. Hocking, R. Bernard, G. W.
Robertson, Effects of low dietary protein and different allocations of food during rearing and restricted feeding after peak rate of lay on egg production, fertility and hatchability in female broiler breeders,
Br.
Poult. Sci,
43, 94-103 (2002)11. R. Nukreaw, C. Bunchasak, Effect of supplementing synthetic amino acids in low-protein diet and subsequent re-feeding on growth performance, serum lipid profile and chemical body composition of broiler chickens,
Japan Poult. Sci. Assoc
, 52, 127-136 (2015) 12. M. S. Ragab, Impact of betainesupplementation to triticale diets varying in their metabolizable energy content on broiler performance during summer season,
Egyptian J. Nutr. Feeds,
16, 113-129 (2013)13. J. H. Kim, Nutritional method for high temperature stress relaxation,
Kor. Poult.
Assoc
, 45, 140-142 (2013)14. K. Sahin, O. Kucuk, A. Hayiril, S. Prasad, Role of dietary zinc in heat stressed poultry,
Poult. Sci
, 88, 2176-2183 (2009)15. M. Ghaffari, M. Shivazad, M. Zaghari, R.
Taherkhani, Effects of different levels of metabolizable energy and formulation of diet based on digestible and total amino acid requirements on performance of male broiler,
Int. J. Poult. Sci
, 6, 276-279 (2007)16. S. Nahashon, N. Adefope, A. Amenyenu, D. Wright, Effects of dietary metabolizable energy and crude protein concentrations on growth performance and carcass characteristics of French guinea broilers,
Poult. Sci
, 84, 337-344 (2005)17. J.B. Sim, D.H. Choi, C.R. Kim, B.S. Park.
Effect of metabolizable energy on homeostasis of duck in high ambient temperature. Korean J. Oil Chemists’ Soc., 34: 132-141 (2017)
18. Korean feeding standard for poultry.
Nutrient requirement of poultry. National Institute of Anmal Science, RDA, Suwon, Korea (2012)
19. P. I. L. Scot, Training manual, Glasgow Univ, UK (1994)
20. S. O. Park, B. S. Park, Effect of feeding the high levels of microcapsulated inulin on egg and blood lipid profile in laying hens,
Korean J. Oil Chemists’ Soc
, 29, 214-223 (2012)21. IBM SPSS, IBM SPSS statistics 22, Algorithms, Chicago: IBM SPSS Inc (2013) 22. L. A. Rubio, R. Ruiz, M. J. Peinado, A.
Echavarri, Morphology and enzymatic activity of the small intestinal mucosa of Iberian pigs as compared with a lean pig strain,
J. Anim. Sci
, 88, 3590-3597 (2010) 23. H. M. Jayaprakasha, Y. C. Yoon, H. D.Paik, Probiotic functional dairy foods and health claims: an overview,
Food Sci.
Biotechnol
, 14, 523-528 (2005)24. X. H. Gu, Y. Hao, X. L. Wang, Overexpression of heat shock protein 70 and its relationship to intestine under acute heat stress in broilers: 2. Intestinal oxidative stress,
Poult. Sci
, 91, 790-799 (2012)25. T. Zeng, J. J. Li, D. Q. Wang, G. Q. Li, G. L. Wang, L. Z. Lu, Effects of heat stress on antioxidant defense system, inflammatory injury, and heat shock proteins of Muscovy and Pekin ducks:
evidence for differential thermal sensitivities,