Insulin-like Growth Factor-I Modulates BDNF Expression by Inhibition of Histone Deacetylase in C2C12 Skeletal Muscle Cells
Hye Jin Kim and Won Jun Lee*
Department of Kinesiology and Sports Studies, College of Science and Industry Convergence, Ewha Womans University, Seoul 03760, Korea Received July 24, 2017 /Revised August 8, 2017 /Accepted August 9, 2017
It is well established that brain-derived neurotrophic factor (BDNF) is expressed not only in the brain but also in skeletal muscle, and is required for normal neuromuscular system function. Histone deace- tylases (HDACs) and insulin-like growth factor-I (IGF-I) are potent regulators of skeletal muscle myo- genesis and muscle gene expression, but the mechanisms of HDAC and IGF-I in skeletal muscle-de- rived BDNF expression have not been examined. In this study, we examined the effect of IGF-I and suberoylanilide hydroxamic acid (SAHA), a pan-HDAC inhibitor, on BDNF induction. Proliferating or differentiating C2C12 skeletal muscle cells were treated with increasing concentrations (0-50 ng/ml) of IGF-I in the absence or presence of 5 μM SAHA for various time periods (3-24 hr). Treatment of C2C12 cells with IGF-I resulted in a dose- and time-dependent decrease in BDNF mRNA expression.
However, inhibition of HDAC led to a significant increase in the expression of BDNF mRNA levels.
In addition, immunocytochemistry revealed high BDNF protein levels in undifferentiated C2C12 skel- etal muscle cells, whether untreated, IGF-I-treated, or exposed to SAHA. These results represent the first evidence that IGF-I can suppress the mRNA and protein expression of BDNF; conversely, SAHA attenuates the effects of IGF-I. Consequently, SAHA upregulates BDNF expression in C2C12 skeletal muscle cells.
Key words : Brain-derived neurotrophic factor, C2C12 skeletal muscle cell, histone deacetylase inhibitor, insulin-like growth factor-I, suberoylanilide hydroxamic acid
*Corresponding author
*Tel : +82-2-3277-2563, Fax : +82-2-3277-2846
*E-mail : [email protected]
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Life Science 2017 Vol. 27. No. 8. 879~887 DOI : https://doi.org/10.5352/JLS.2017.27.8.879
Introduction
Histone deacetylases (HDACs) have been associated with multiple physiological processes in various cell types, in- cluding skeletal muscle cells. Many studies have demon- strated that HDAC can control metabolic-related processes and muscle-specific gene expression, including myocyte spe- cific enhancer factor-2 (MEF2), myogenic regulatory factors (MRFs) and myosin heavy chain (MHC) in skeletal muscle cells [7, 16, 32]. Therefore, many researchers have inves- tigated the function and molecular mechanisms of HDAC and its’ pharmacological inhibitors in skeletal muscle.
Several studies have reported that skeletal muscle has the capacity to express a number of myokines in response to exercise or contraction of muscle fibers. It is now clear that interleukin (IL)-6, IL-8, IL-15, brain-derived neurotrophic
factor (BDNF), leukemia inhibitory factor (LIF), myostatin and fibroblast growth factor 21 (FGF21) production is upre- gulated by muscle contraction [27, 28]. These myokines work in a hormone-like fashion and play an important role in skel- etal muscle mass and metabolic homeostasis [26, 30]. Among the myokines, BDNF is essential for neuronal survival, brain development, synaptic plasticity, and hippocampal function [25, 31]. Indeed, blocking BDNF signaling decreases memory function and increases anxiety and depression [9]. In skeletal muscle, BDNF is highly expressed in satellite cells. Several studies reported that BDNF levels are associated with devel- oping myofibers, and expression of myosin heavy chain, which is a marker of muscle growth [8, 15, 22]. In addition, BDNF has been considered as a possible mediator of the positive effects of HDAC inhibitors in neurodegenerative and psychiatric disorders [4]. Koppel and Timmusk [17] re- ported that the pan-HDAC inhibitor suberoylanilide hy- droxamic acid (SAHA, also known as vorinostat) sig- nificantly increased BDNF mRNA levels in primary rat cort- ical neurons. Due to the fact that BDNF is a well-known neurological factor (rather than a myokine), it is still not clear whether HDAC inhibitors are associated with changes in BDNF expression in skeletal muscle cells.
A
B
Fig 1. Schematic schedule of C2C12 skeletal muscle cell culture and drug treatments. (A) C2C12 cells were cultured in growth medium (GM) for 2 days, at which point the cells mainly consist of proliferating myoblasts. Cells were placed in GM or differentiation medium (DM) containing IGF-I (0-50 ng/ml) and/or HDAC inhibitor SAHA (5 μM) for 24 hr. (B) C2C12 cells were incubated in GM for 2days. For differentiation, 85-90% confluent cultures were placed in DM for 3days. The differentiated cells were treated with DM containing IGF-I (0-50 ng/ml) and/or SAHA (5 μM).
Insulin-like growth factor-I (IGF-I) is a potent regulator of skeletal muscle mass in mammals. In fact, IGF-I alters transcription and translation of myogenic factors that regu- late myocyte proliferation and differentiation [5, 14]. In addi- tion, administration of IGF-I to skeletal muscle increases muscle cross-sectional area and force generation in animals [2, 14]. Therefore, IGF-I can act through multiple mecha- nisms to enhance skeletal muscle function. Mitsiades et al.
[19] reported a possible interaction between IGF-I and HDAC inhibitors in a human multiple myeloma (MM) cell line. They observed that SAHA suppressed autocrine IGF-I production and IGF-I gene expression in MM cells. Even though such HDAC inhibitor modulations of IGF-I have been studied in neuronal cells, little is known about its’ mo- lecular regulation in skeletal muscle cells.
As previously mentioned, HDAC inhibitors can modulate BDNF expression. Although previous studies suggest a pos- sible interaction between HDAC and IGF-I in nerve cells, the effects of IGF-I and HDAC on the induction of BDNF have not been demonstrated in skeletal muscle. Therefore, the purpose of this study was to investigate the effect of IGF-I and pan-HDAC inhibitor SAHA on BDNF expression in C2C12 skeletal muscle cells. Here, we found the sup- pression of BDNF at the myogenic differentiation stage.
Furthermore, IGF-I administration decreased BDNF ex- pression. Finally, treatment with SAHA rescued BDNF mRNA and protein expression in IGF-I-induced suppression of BDNF.
Materials and Methods
Cell cultures, IGF-I and drug treatment
C2C12 mouse skeletal muscle cells were obtained from the American Type of Culture Collection (ATCC, Rockville, MD). Cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM) (Welgene, KOREA) supplemented with 10% fetal bovine serum (FBS) (Thermo Fisher Scientific, Waltham, MA) and antibiotics (100 U/ml of penicillin G and 100 μg/ml streptomycin) (Welgene, KOREA) in an atmos- phere composed of 95% air and 5% CO2 at 95% humidity and 37°C. The cells used in all experiments conducted for this study were at passage 4 to 7. For the experiments, C2C12 myoblasts were plated in six-well culture plates at a density of 2.5×105 cells/well in growth medium (GM; DMEM, 10%
FBS). For all experiments, cells at 90% confluence were treat- ed with DMEM (Welgene, KOREA) supplemented with 2%
horse serum (Sigma, St. Louis, MO), antibiotics (100 U/ml of penicillin G and 100 μg/ml streptomycin) (Welgene, KOREA), and various concentrations (0-50 ng/ml) of IGF-I (Sigma, St. Louis, MO) for 0 to 24 hr in the absence or pres- ence of 5 μM of SAHA (Sigma, St. Louis, MO) as indicated in the Fig. 1.
Western blot
Cells were lysed and scraped in ice-cold RIPA buffer (50 mM Tris, pH 7.5, 150 mM NaCl, 1% Triton X-100, 0.1% SDS, 0.5% Sodium deoxycholate, 2 mM EDTA, 1 mM PMSF, com-
plete protease inhibitor and phosphatase inhibitor cocktails).
The cell extracts were then centrifuged at 13,000 rpm for 15 min at 4°C. Next, the protein in the supernatant was quantified using a Bradford protein assay kit (Bio-Rad, Hercules, CA). Sixty micrograms of total protein were re- solved on 12% SDS-PAGE gel (90 V, 25°C, 20min, after 150 V, 25°C, 1 hr) and then transferred to PVDF membranes (12 V, 25°C, 1 hr). All of the blots were then incubated with Ponceau S (Sigma, St. Louis, MO) to ensure equal loading in all lanes (data not shown). For the detection of BDNF and GAPDH proteins, the membranes were blocked with 5% skim milk in Tris-buffered saline (TBS) with 0.1%
Tween20 (0.1% TBST) for 1 h at room temperature and sub- sequently incubated with BDNF (sc-546) (polyclonal rabbit antibody, 1:2,000) (Santa Cruz Biotechnology, Santa Cruz, CA), or GAPDH (#2118) (polyclonal rabbit antibody, 1:5,000) (Cell Signaling, Beverley, MA) antibodies overnight at 4°C in 5% BSA in 0.1% TBST. The membranes were then washed three times for 5 min each in 0.1% TBST, after which they were incubated for 1 hr with anti-rabbit IgG horse-radish peroxidase-linked secondary antibody (1:2,500) (Cell signal- ing, Beverley, MA). The membranes were then washed as described above, after which EZ-western Lumi Pico reagent (Daeillab Service Co, Ltd, Korea) was applied according to the manufacturer’s instructions to develop a signal that was subsequently detected using the Chemiluminescence imag- ing system (ATTO, Tokyo, Japan) and quantified by densi- tometry using Image J software (NIH, USA). The target pro- tein levels were then normalized against the GAPDH protein levels.
RNA extract and real-time PCR
Total RNA was extracted from C2C12 cells using the phe- nol-chloroform extraction method with TRIzol Reagent (Invitrogen Life Technologies, Carlsbad, CA) according to the manufacturer’s instructions, after which the RNA was quantified using a spectrophotometer (Nanodrop-2000, Thermo Fisher Scientific, Waltham, MA). Next, cDNA was synthesized from 1 μg of total RNA in the presence of ran- dom primer, 2.5 mM dNTP, Rnase inhibitor, and reverse transcriptase (Invitrogen Life Technologies, Carlsbad, CA) in a final volume of 20 μl at 25°C for 10 min, followed by 42°C for 60 min and 95°C for 5 min. The sequences of the primers were as follows: BDNF, forward (F) 5’-TCATACTTCGG TTGCATGAAGG-3’, and reverse (R) 5’-AGACCTCTCGAAC CTGCCC-3’; GAPDH (F) 5’-ATGACAATGAATACGGCT
ACAGCAA-3’, (R) 5’-GCAGCGAACTTTATTGATGGTATT- 3’. The primers were purchased from Macrogen (Macrogen, KOREA). Real-time qPCR was performed in duplicate using the SYBR Green PCR master mix (Kappa, USA) according to the manufacturer’s instructions. All PCR amplifications were conducted using an ABI Step One Plus system (Applied Biosystems Inc., Foster City, CA). The expression of the target genes was then normalized against the ex- pression of glyceraldehydes 3-phosphate dehydrogenase (GAPDH).
Immunocytochemistry
The cultured and treated cells were then fixed in 4% form- aldehyde for 20 min at room temperature. Next, the cells were washed three times each in PBS, after which they were permeabilized with PBS containing 0.2% triton X-100 (0.2%
PBST) for 10 min at room temperature. The cells were then washed three times for 5 min each in PBS and then blocked with 5% BSA in PBS for 1 hr at room temperature. Next, the cells were washed once with PBS, after which they were probed with BDNF (sc-546) polyclonal rabbit antibody (Santa Cruz Biotechnology, Santa Cruz, CA) at a dilution of 1:500 overnight at 4°C in 5% BSA in PBS. The cells were then washed three times for 5 min each in 0.1% PBST, after which they were incubated with Alex488-conjugated goat anti-rabbit IgG secondary antibody (Invitrogen Life Technol- ogies, Carlsbad, CA) diluted 1:200 for 20 min at room tem- perature in PBS that contained 5% BSA. Next, the cells were washed three times with 0.1% PBST, after which they were stained with mounting media containing 4’,6-diamidino- 2phenylinodole (DAPI) at a concentration of 1.5 μg/ml to localize the nuclei. The cells were washed three times with PBS. After, the cells were viewed and photographed using an EVOS® FL Cell Imaging System (Thermo Fisher Scientific, Waltham, MA).
Data analysis
All values are reported as means ± SE. Statistical sig- nificance was determined by t-test and/or one-way ANOVA with post hoc Tukey’s test using SPSS 22.0. Differences be- tween groups were considered significant at p<0.05.
Results
BDNF mRNA levels are higher in myoblasts than differentiating or differentiated myotubes
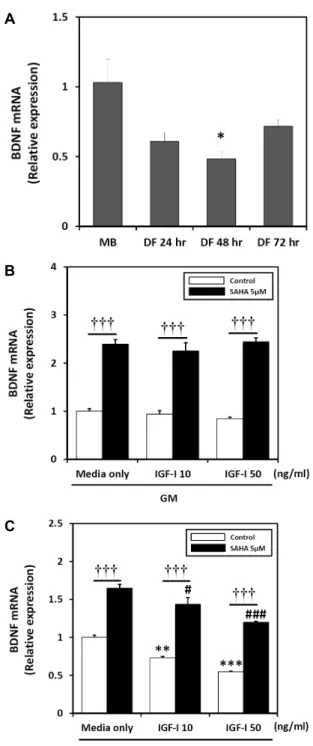
We first observed, by real-time qPCR, that differentiation media (DM) conditions suppress BDNF gene expression (Fig. 2A).
BDNF mRNA expression is decreased by IGF-I, but upregulated by SAHA in cultured C2C12 cells in growth medium (GM) and DM
Next, to test the effect of IGF-I and/or SAHA on BDNF gene expression in GM or DM conditions, we added 0-50 ng/ml of IGF-I with or without 5 μM of SAHA. BDNF mRNA levels were determined by real-time qPCR and GAPDH mRNA abundance was examined as a correction factor. The results revealed no change in BDNF mRNA lev- els in response to 24 hr IGF-I treatment in GM. Conversely, when cells were co-treated with SAHA and IGF-I in GM, the expression of BDNF mRNA was upregulated. BDNF mRNA expression increased by 139% after 24 hr of SAHA treatment compared to GM alone. HDAC inhibitor and IGF-I- (10 or 50 ng/ml) treated cells increased BDNF mRNA expression by 132% and 159% compared to 10 ng/ml of IGF-I and 50 ng/ml of IGF-I, respectively (Fig. 2B). Next, we examined cells exposed to IGF-I and/or SAHA during differentiation for 24 hr. We found significant decreases in BDNF mRNA expression at every tested IGF-I concentration in DM. However, SAHA significantly increased BDNF mRNA levels 24 hr after treatment with IGF-I in DM (Fig.
2C).
The effect of IGF-I on BDNF protein expression in C2C12 skeletal muscle cells depends on the growth or differentiation stage
We next analyzed BDNF protein expression in response to IGF-I and/or SAHA in C2C12 cells under different media conditions. As expected, the expression of BDNF was sup- pressed in IGF-I treated cells. Conversely, SAHA treatment resulted in induction of BDNF protein expression. BDNF was highly expressed in C2C12 cells in response to SAHA without IGF-I in GM (Fig. 3A), but not in IGF-I treated cells, especially when cells were incubated in DM (Fig. 3B and Fig. 3C).
IGF-I inhibits BDNF mRNA expression, but not in SAHA-treated differentiated myotubes at different time points
Real-time qPCR analysis revealed that BDNF gene ex- pression varied in response to treatment with IGF-I and/or
A
B
C
Fig 2. BDNF mRNA levels determined by real-time qPCR in C2C12 cells cultured for 24 hr in the absence (control) or presence of IGF-I (0-50 ng/ml) with HDAC inhibitor SAHA (5 µM) at different medium condition. (A) Cells in different media conditions without IGF-I and SAHA.
(B) Growth medium. (C) Differentiation medium. Target mRNA values are shown normalized to the GAPDH mRNA level for each sample. Samples were analyzed in parallel with GAPDH. Values are means ± SE of three independent experiments. **p<0.01, ***p<0.001; medium only control vs. IGF-I. #p<0.05, ###p<0.001; medium on- ly SAHA vs. IGF-I with SAHA. †††p<0.001; IGF-I vs.
IGF-I with SAHA.
A
B
C
Fig 3. Immunocytochemistry images showing BDNF protein expression. All images were acquired at 24 hr after treat- ment, and BDNF was detected by immunofluorescence (green). (A) C2C12 cells in IGF-I free and/or 5 μM of SAHA at different media conditions. (B) Cells in me- dium containing 10 ng/ml IGF-I and/or 5 μM of SAHA at GM or DM. C: C2C12 cells treated with 50 ng/ml IGF-I and/or 5 μM of SAHA at different media con- ditions.
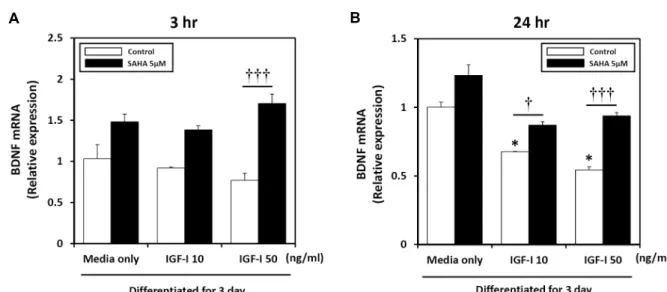
SAHA for different periods of time (Fig. 4). Specifically, treatment of C2C12 cells with 0-50 ng/ml of IGF-I for 3 hr (Fig. 4A) or 24 hr (Fig. 4B) resulted in a time- and concen- tration-dependent decrease in BDNF mRNA expression,
with the maximal effect being observed after 24 hr with 50 ng/ml IGF-I treatment. BDNF mRNA expression was eval- uated to further investigate the modulating effect of SAHA on BDNF gene expression levels in IGF-I-treated myotubes.
BDNF mRNA expression increased significantly by 94% af- ter 3 hr of SAHA treatment compared to 50 ng/ml IGF-I treatment (Fig. 4A). Similarly, expression of BDNF increased by 23% after 24 hr of SAHA treatment compared to medium only (Fig. 4B). In addition, reduced BDNF mRNA levels in- duced by IGF-I returned to basal levels after 24 hr SAHA treatment. BDNF expression increased significantly by 20%
and 40% after 24 hr of SAHA treatment compared to 10 and 50 ng/ml of IGF-I. Taken together, these data suggest that SAHA induces BDNF expression, which was suppressed by IGF-I.
SAHA rescues BDNF protein expression in IGF-I- induced BDNF suppression
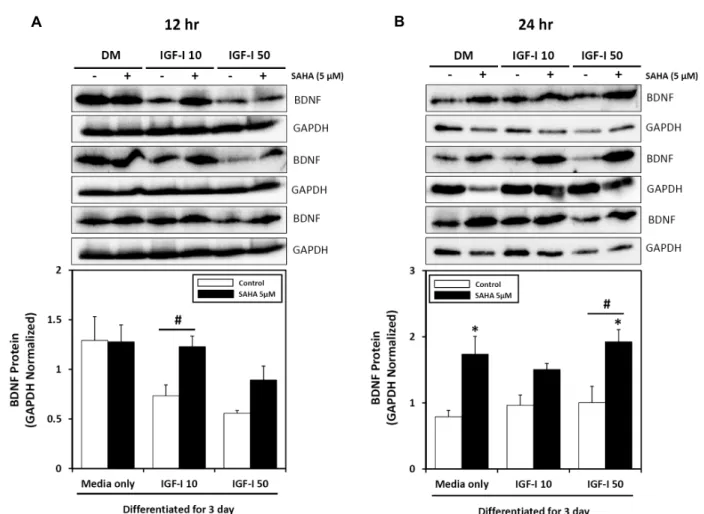
To determine whether BDNF protein followed the same trend as its’ transcript after myogenic differentiation, we per- formed western blot analysis after IGF-I treatment, with or without SAHA treatment on 3 day differentiated myotubes.
Treatment of IGF-I for 12 hr resulted in decreased BDNF protein levels in differentiated C2C12 cells (Fig. 5A). In con- trast, IGF-I and SAHA co-treatment increased BDNF protein expression in differentiated cells. Furthermore, treatment of IGF-I in differentiated C2C12 myotubes for 24 hr did not alter BDNF protein expression. However, supplementation of SAHA increased total BDNF protein expression up to 80%
(Fig. 5B). These data suggest that SAHA modulates BDNF expression in differentiated C2C12 skeletal muscle cells.
Discussion
Skeletal muscle-derived BDNF is a myokine and a puta- tive regulatory factor in skeletal muscle. BDNF is a potent regulator of skeletal muscle satellite cell function, muscle re- generation, survival of motor neurons and lipid metabolism, as well as neurogenesis, synaptic plasticity, and cognitive ability [10, 25, 28, 31]. However, very little is known about BDNF and its’ interactions with various muscle factors such as IGF-I or other hormones in skeletal muscle. According to several recent studies, BDNF expression is regulated by epigenetic modifications in various cell lines, including skel- etal muscle cells. For example, BDNF mRNA is upregulated by HDAC inhibitors, such as MS-275, MC1568 and SAHA
A B
Fig 4. Real-time qPCR was performed at different time points to assess endogenous BDNF mRNA levels by the administration of IGF-I and HDAC inhibitor. (A) Differentiated C2C12 cells were treated with 0-50 ng/ml of IGF-I and 5 μM of SAHA for up to 3 hr. (B) Effect of IGF-I and/or HDAC inhibitor on BDNF mRNA expression in differentiated C2C12 cells. Cells were treated with IGF-I and SAHA for 24 hr. Target mRNA values are shown normalized to the GAPDH mRNA level for each sample. Samples were analyzed in parallel with GAPDH. Values are means ± SE of three independent experiments.
*p<0.05; medium only control vs. IGF-I. †p<0.05, †††p<0.001; IGF-I vs. IGF-I with SAHA.
in primary rat cortical neurons [17]. However, the modulat- ing effect of IGF-I and SAHA in the regulation of HDAC expression in skeletal muscle remains poorly understood.
Here, we report that BDNF mRNA and protein levels are regulated by IGF-I and HDAC inhibitor SAHA.
The primary finding of this study is that BDNF mRNA and protein levels were higher in proliferating myoblasts compared to differentiating or differentiated myotubes.
Mousavi and Jasmin [21] determined that BDNF mRNA lev- els were highly correlated with the level of the skeletal mus- cle satellite cell marker, Pax3. Moreover, Pax7 expression is reduced and satellite cell function is defective in BDNFMKO (muscle-specific conditional knockout) mice. As a result, muscle regeneration is significantly delayed [6]. Conversely, BDNF mRNA and protein levels decreased 5-fold in 4-day- differentiated L6 rat skeletal muscle cells [21]. Another re- cent study has also shown that BDNF expression is reduced in C2C12 cells during myogenic differentiation in a time-de- pendent manner [20]. Therefore, our results are consistent with previous studies showing higher BDNF expression re- lated to satellite cells or myoblasts being inhibited in differ- entiated cells.
Many studies have investigated the regulation of skeletal muscle cell proliferation, differentiation and expression of muscle-related genes by epigenetic modifications, including histone acetylation/methylation, DNA methylation, and ex-
pression of micro RNAs. For example, in the absence of dif- ferentiation-promoting signals, HDACs interact with MRFs and MEF2 in proliferating myoblasts [3, 7, 12, 18]. In addi- tion, exercise-induced skeletal muscle contraction can regu- late histone acetyltransferases (HATs) and HDAC activity [23, 24]. Zwetsloot et al. [32] reported that MHC gene ex- pression can be altered by differential epigenetic mod- ifications of histones in fast and slow skeletal muscle fibers.
This suggests that the balance of HAT and HDAC activity, or the balance of histone methyl transferases (HMTs) and histone demethylases (HDMs) modulate chromatin structure and transcriptional activity of MHC genes. Based on our ini- tial findings and previous publications, we investigated whether or not histone acetylation through the inhibition of HDAC can modulate BDNF mRNA and protein expression.
As expected, the level of BDNF mRNA and protein were higher in C2C12 cells treated with the pan-HDAC inhibitor SAHA compared to untreated cells. This result was more significant in proliferating cells than differentiated cells. Our current data clearly demonstrates that BDNF expression can be influenced by HDAC in C2C12 skeletal muscle cells.
The second finding of this study is that IGF-I treatment led to decreased BDNF expression. In skeletal muscle, IGF-I and BDNF have been studied and discussed for decades, but we know almost nothing about their interactions in skel- etal muscle. IGF-I is a well-known effector in skeletal muscle.
A B
Fig. 5. Influence of treatment with IGF-I and HDAC inhibitor for different lengths of time on BDNF protein expression in differentiated C2C12 skeletal muscle cells. Total cell lysates were obtained and analyzed by western blotting for the presence of BDNF.
(A) Differentiated C2C12 myotubes were treated with IGF-I (0-50 ng/ml) for 12 hr in the absence or presence of HDAC inhibitor (5 μM of SAHA) with DM. (B) Differentiated C2C12 myotubes were treated with IGF-I (0-50 ng/ml) for 24 hr in the absence or presence of HDAC inhibitor (5 μM of SAHA) with DM. Values are means ± SE with n = 3 for each condition. *p<0.05; vs. SAHA. #p<0.05; IGF-I vs. IGF-I with SAHA.
Indeed, IGF-I stimulation maintains skeletal muscle volume via various anabolic actions or inhibition of ubiquitin ligase (e.g., MURF-1 and MAFbx/Atrogin-1) expression and the protein degradation [5, 14, 29]. IGF-I also induces satellite cell activation, myoblast proliferation, differentiation and in- crease of myotube size [2, 13, 14]. In addition, satellite cells stimulate IGF-I for muscle regeneration in response to injury [1]. Moreover, resistance exercise training can also induce IGF-I in skeletal muscle [2, 5, 29]. Despite IGF-I and BDNF being crucial effectors in muscle, their molecular interactions have not been demonstrated. Therefore, we sought to eluci- date how IGF-I influences BDNF expression. Surprisingly, IGF-I reduced BDNF expression in C2C12 cells. This is a somewhat unexpected finding, as many studies have re- ported that both IGF-I and BDNF play pivotal roles in skel- etal muscle cell activation. In a similar finding, exercise can
increase pro-inflammatory cytokines such as IL-6 and/or IL-1β (also known as myokines) and elevating these signals could produce feedback to inhibit IGF-I [1, 11]. However, there are insufficient reports supporting our data. Finally, we investigated whether or not SAHA could modulate the IGF-I-induced BDNF suppression. As a previously men- tioned, we demonstrated that SAHA can upregulate ex- pression of BDNF. As expected, SAHA rescued the sup- pression of BDNF mRNA and protein levels due to the IGF-I treatment. Similar research has demonstrated that SAHA downregulates IGF-I and its receptor (IGF-IR) in human MM cells [19].
Most notably in the present study, HDAC inhibitor SAHA effectively modulated BDNF expression. We also found the SAHA can attenuate IGF-I-induced BDNF suppression in skeletal muscle cells. Therefore, our study clearly demon-
strates the effects of IGF-I and SAHA on BDNF mRNA and protein expression in C2C12 skeletal muscle cells. Taken to- gether, our data provide evidence that the primary role of SAHA in skeletal muscle cells is to upregulate BDNF by in- hibiting IGF-I. Therefore, SAHA and IGF-I would appear to be putative mediators of other myokines in skeletal muscle cells. The limitation of this study is that the molecular mech- anisms of the signaling pathways integrating IGF-I and SAHA regulation of BDNF expression in skeletal muscle cells have not been investigated. Thus, further studies will be necessary to elucidate the mechanisms by which IGF-I and specific histone deacetylases or acetyltransferases act on BDNF expression in skeletal muscle.
References
1. Adams, G. R. 2002. Autocrine/paracrine IGF-I and skeletal muscle adaptation. J. Appl. Physiol. 93, 1159-1167.
2. Anderson, B. C., Christiansen, S. P., Grandt, S., Grange, R.
W. and McLoon, L. K. 2006. Increased extraocular muscle strength with direct injection of insulin-like growth factor-I.
Invest. Ophthalmol. Vis. Sci. 47, 2461-2467.
3. Bharathy, N., Ling, B. M. and Taneja, R. 2013. Epigenetic regulation of skeletal muscle development and differ- entiation. Subcell. Biochem. 61, 139-150.
4. Chen, P. S., Peng, G. S., Li, G., Yang, S., Wu, X., Wang, C. C., Wilson, B., Lu, R. B., Gean, P. W., Chuang, D. M.
and Hong, J. S. 2006. Valproate protects dopaminergic neu- rons in midbrain neuron/glia cultures by stimulating the release of neurotrophic factors from astrocytes. Mol.
Psychiatry 11, 1116-1125.
5. Clemmons, D. R. 2009. Role of IGF-I in skeletal muscle mass maintenance. Trends Endocrinol. Metab. 20, 349-356.
6. Clow, C. and Jasmin, B. J. 2010. Brain-derived neurotrophic factor regulates satellite cell differentiation and skeletal muscle regeneration. Mol. Biol. Cell. 21, 2182-2190.
7. Cohen, T. J., Barrientos, T., Hartman, Z. C., Garvey, S. M., Cox, G. A. and Yao, T. P. 2009. The deacetylase HDAC4 controls myocyte enhancing factor-2-dependent structural gene expression in response to neural activity. FASEB. J. 23, 99-106.
8. Colombo, E., Bedogni, F., Lorenzetti, I., Landsberger, N., Previtali, S. C. and Farina, C. 2013. Autocrine and immune cell-derived BDNF in human skeletal muscle: implications for myogenesis and tissue regeneration. J. Pathol. 231, 190-198.
9. Cotman, C. W. and Berchtold, N. C. 2002. Exercise: a behav- ioral intervention to enhance brain health and plasticity.
Trends Neurosci. 25, 295-301.
10. Cotman, C. W., Berchtold, N. C. and Christie, L. A. 2007.
Exercise builds brain health: key roles of growth factor cas- cades and inflammation. Trends Neurosci. 30, 464-472.
11. Dey, B. R., Furlanetto, R. W. and Nissley, P. 2000. Suppres-
sor of cytokine signaling (SOCS)-3 protein interacts with the insulin-like growth factor-I receptor. Biochem. Biophys. Res.
Commun. 278, 38-43.
12. Fu, X., Zhao, J. X., Liang, J., Zhu, M. J., Foretz, M., Viollet, B. and Du, M. 2013. AMP-activated protein kinase mediates myogenin expression and myogenesis via histone deacety- lase 5. Am. J. Physiol. 305, C887-895.
13. Galvin, C. D., Hardiman, O. and Nolan, C. M. 2003. IGF-I receptor mediates differentiation of primary cultures of mouse skeletal myoblasts. Mol. Cell. Endocrinol. 200, 19-29.
14. Glass, D. J. 2003. Signalling pathways that mediate skeletal muscle hypertrophy and atrophy. Nat. Cell. Biol. 5, 87-90.
15. Griesbeck, O., Parsadanian, A. S., Sendtner, M. and Thoenen, H. 1995. Expression of neurotrophins in skeletal muscle:
quantitative comparison and significance for motoneuron survival and maintenance of function. J. Neurosci. Res. 42, 21-33.
16. Iezzi, S., Cossu, G., Nervi, C., Sartorelli, V. and Puri, P. L.
2002. Stage-specific modulation of skeletal myogenesis by inhibitors of nuclear deacetylases. Proc. Natl. Acad. Sci. USA 99, 7757-7762.
17. Koppel, I. and Timmusk, T. 2013. Differential regulation of BDNF expression in cortical neurons by class-selective his- tone deacetylase inhibitors. Neuropharmacology 75, 106-115.
18. McKinsey, T. A., Zhang, C. L. and Olson, E. N. 2001. Control of muscle development by dueling HATs and HDACs. Curr.
Opin. Genet. Dev. 11, 497-504.
19. Mitsiades, C. S., Mitsiades, N. S., McMullan, C. J., Poulaki, V., Shringarpure, R., Hideshima, T., Akiyama, M., Chauhan, D., Munshi, N., Gu, X., Bailey, C., Joseph, M., Libermann, T. A., Richon, V. M., Marks, P. A. and Anderson, K. C. 2004.
Transcriptional signature of histone deacetylase inhibition in multiple myeloma: biological and clinical implications.
Proc. Natl. Acad. Sci. USA 101, 540-545.
20. Miura, P., Amirouche, A., Clow, C., Bélanger, G. and Jasmin, B. J. 2012. Brain-derived neurotrophic factor ex- pression is repressed during myogenic differentiation by miR-206. J. Neurochem. 120, 230-238.
21. Mousavi, K. and Jasmin, B. J. 2006. BDNF is expressed in skeletal muscle satellite cells and inhibits myogenic differ- entiation. J. Neurosci. 26, 5739-5749.
22. Mousavi, K., Parry, D. J. and Jasmin, B. J. 2004. BDNF res- cues myosin heavy chain IIB muscle fibers after neonatal nerve injury. Am. J. Physiol. 287, C22-29.
23. Ntanasis-Stathopoulos, J., Tzanninis, J. G., Philippou, A. and Koutsilieris, M. 2013. Epigenetic regulation on gene ex- pression induced by physical exercise. J. Musculoskelet.
Neuronal. Interact. 13, 133-146.
24. Pandorf, C. E., Haddad, F., Wright, C., Bodell, P. W. and Baldwin, K. M. 2009. Differential epigenetic modifications of histones at the myosin heavy chain genes in fast and slow skeletal muscle fibers and in response to muscle unloading.
Am. J. Physiol. 297, C6-16.
25. Park, H. and Poo, M. M. 2013. Neurotrophin regulation of neural circuit development and function. Nat. Rev. Neurosci.
14, 7-23.
초록:C2C12 골격근 세포에서 히스톤 탈 아세틸 효소의 억제가 인슐린 유사성장인자(IGF-I)에 의한 BDNF 발현 조절에 미치는 영향
김혜진․이원준*
(이화여자대학교 신산업융합대학 체육과학과)
히스톤 탈 아세틸 효소(HDAC)와 인슐린유사성장인자(IGF-I)는 근육 관련 유전자들의 활성 및 발현을 조절하 여 골격근의 성장 및 발달을 조절하지만 이들이 근신경계 발달 및 대사 기능에 중요한 역할을 담당하는 뇌신경성 장인자(BDNF)의 발현에 미치는 영향에 관한 연구는 거의 이루어지지 않았다. 따라서 본 연구에서는 IGF-I과 HDAC의 억제제인 SAHA가 C2C12 골격근 세포에서 BDNF 발현에 미치는 영향을 알아보고자 하였다. 그 결과 IGF-I은 농도와 시간 의존적으로 BDNF의 mRNA 및 단백질 발현을 감소시켰지만 HDAC을 억제하자 IGF-I에 의 해 감소되었던 BDNF의 발현이 증가하는 경향을 관찰할 수 있었다. 따라서 IGF-I은 BDNF의 발현을 억제하며, HDAC의 억제는 IGF-I에 의한 BDNF의 발현 억제를 감소시킬 수 있다는 사실을 확인할 수 있었다.
26. Pedersen, B. K. and Febbraio, M. A. 2008. Muscle as an en- docrine organ: focus on muscle-derived interleukin-6.
Physiol. Rev. 88, 1379-1406.
27. Pedersen, B. K. 2009. Muscle as an endocrine organ: IL-6 and other myokines. J. Appl. Physiol. 107, 1006-1014.
28. Pedersen, B. K. and Febbraio, M. A. 2012. Muscles, exercise and obesity: skeletal muscle as a secretory organ. Nat. Rev.
Endocrinol. 8, 457-465.
29. Sacheck, J. M., Ohtsuka, A., McLary, S. C. and Goldberg, A. L. 2004. IGF-I stimulates muscle growth by suppressing protein breakdown and expression of atrophy-related ubiq- uitin ligases, atrogin-1 and MuRF1. Am. J. Physiol. 287, E591- 601.
30. Wrann, C. D., White, J. P., Salogiannnis, J., Laznik-Bogoslav- ski, D., Wu, J., Ma, D., Lin, J. D., Greenberg, M. E. and Spiegelman, B. M. 2013. Exercise induces hippocampal BDNF through a PGC-1α/FNDC5 pathway. Cell. Metab. 18, 649-659.
31. Xu, B. 2013. BDNF (I)rising from exercise. Cell. Metab. 18, 612-614.
32. Zwetsloot, K. A., Laye, M. J. and Booth, F. W. 2009. Novel epigenetic regulation of skeletal muscle myosin heavy chain genes. Focus on "Differential epigenetic modifications of his- tones at the myosin heavy chain genes in fast and slow skel- etal muscle fibers and in response to muscle unloading".
Am. J. Physiol. 297, C1-3.