48(2) : 155∼ 159 (2017)
155
꾸지뽕나무 뿌리 추출물의 파골세포 분화 억제 효과
김유경·정길생*
계명대학교 약학대학
Inhibitory Effects of the Roots of Cudrania tricuspidata Bureau on Osteoclast Differentiation
Yu-Gyeong Kim and Gil-Saeng Jeong*
College of Pharmacy, Keimyung University, 1095 Dalgubeol-daero, Daegu 42601, Korea
Abstract − Cudrania tricuspidata Bureau (Moraceae) is a traditional oriental medicine that has been widely used as anti-oxi- dant, anti-inflammatory and immunomodulatory in Korea. This study was performed that the 70% ethanol extract of the roots of C. tricuspidata (CTE) suppressed receptor activator of NF-κB ligand (RANKL)-induced osteoclastogenesis, actin ring for- mation in RAW 264.7 cell lines. CTE significantly inhibited the JNK/mitogen-activated protein kinase (MAPK) signaling path- way without affecting ERK and p38 signaling in RANKL-stimulated RAW 264.7 cells. Also, CTE inhibited RANKL-induced expression of c-Fos, an upstream activator of NFATc1. Consequently, CTE suppresses osteoclast differentiation by inhibiting RANKL induced MAPK signaling pathways and disrupts the actin rings in mature osteoclasts. Thus, CTE can be used for the development of osteoporosis treatment drug with a natural material.
Keywords − Cudrania tricuspidata Bureau, Osteoclastogenesis, RANKL, MAPK, c-Fos, NFATc1
꾸지뽕나무(Cudrania tricuspidata Bureau)는 뽕나무과의 낙엽성 소교목으로 잎과 근피에 함유되어 있는 플라보노이 드는 높은 항산화성을 가지고 있으며 예로부터 민간에서 자 양, 강장, 신체허약증, 정력 감퇴 등을 개선시키는 효과가 있는 것으로 알려져있으며, 특히 줄기껍질과 뿌리는 여성질 환에 사용되었다.1) 최근 꾸지뽕나무의 잎과 열매 및 줄기 추출물에서 산화적 손상에 대한 간세포 보호효과2) 연구와 항산화1), 항암3)등에 관한 연구가 보고되고 있다. 꾸지뽕나 무에서 분리한 주요 성분으로는 xanthones, flavonoids, benzenoids4) 등의 폴리페놀 화합물들이 보고 되었다.
골다공증은 골량 및 골밀도 감소를 특징으로 한 전신 골 격계 질환으로 골절되기 쉬운 질병으로 알려져있다.5) 골은 골을 형성하는 조골세포와 골을 흡수하는 파골세포에 의해 항상성을 유지하며 균형을 맞추며6) 골 질환은 조골세포의 골 형성 보다는 골을 흡수하는 파골세포 활성 증가가 주된 원인이다.7) 그러므로 골다공증을 예방 또는 치료하는데 있 어서 파골세포 분화를 억제하는 것은 중요한 수단이 될 것 이다. 파골세포의 지표라 할 수 있는 tartrate-resistant acid phosphatase(TRAP)은 파골세포에만 존재하는 골 흡수 효소
로서 TRAP 양성 세포는 파골세포의 분화를 판단하는 기준 이 된다.8) 또한 파골세포에 있어서 actin ring의 형성은 뼈 를 흡수할 수 있는 능력에 대한 중요한 표지이다.9) 조골세 포는 골 재형성을 위해 성숙 과정을 거치며, 성숙된 조골세 포에서 생성된 receptor activator of nuclear factor kappa-B ligand(RANKL)이 파골세포의 수용체 RANK에 결합하여 파골세포의 성숙과 분화가 이루어지며 RANKL/RANK/
TNFR associated factors(TRAF) 복합체가 형성이 되고 nuclear factor-κB(NF-κB)와 ERK, JNK, p38를 포함하는 mitogen-activated protein kinase(MAPK)를 활성화시킨다.10,11) 또한 RANKL은 파골세포 분화 필수 전사인자인 NFATc1와 c-Fos를 활성화시킨다.12)
따라서 본 연구에서는 파골세포 분화를 억제하여 골다공 증의 치료와 예방 가능한 천연물로 꾸지뽕나무 뿌리를 대 상으로 파골세포 분화에 대한 억제 효능과 작용기전을 규 명하고자 하였다.
재료 및 방법
실험 재료 − 본 실험에 사용된 꾸지뽕나무 뿌리는 2014 년 6월 대구광역시 약령 시장에서 건조된 것을 구입하여 사
*교신저자(E-mail):[email protected] (Tel): +82-53-580-6649
용하였다. 형태학적 평가를 통하여 동정하였고 표본시료 (KMP-2014-21)는 계명대학교 약학대학 생약학연구실에 보 관하였다.
시약 및 기기 − Recombinant soluble RANKL ligand (RANKL)는 PeproTech사 제품을 사용하였으며, 3'-(4,5- dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide(MTT) 는 Amresco사, dimethyl sulfoxide(DMSO)는 AppliChem사 에서 구입 하였다. Minimum essential medium α(α-MEM), Dulbecco’s modified Eagle’s medium(DMEM)은 Gibco Laboratories 사에서 구입하였으며, phosphate-buffered saline (PBS), fetal bovine serum(FBS)는 Welgene사에서 구입하였 다. 4-6-Diamidino-2-phenylindole(DAPI)는 Sigma Aldrich 사 제품을 사용하였으며, Alexa flour 488은 Life Technologies 사 제품을 사용하였다. Primary antibody로 phosphor-JNK, JNK, phosphor-ERK, ERK, phosphor-p38, p-38 그리고 rabbit polyclonal 항체는 Cell Signaling Technology사 제품 을 사용하였으며, c-Fos는 Santa cruz사, anti-nuclear factor of activated T cells-c1(NFATc1)은 BD biosciences사 제품 을 사용하였다. Chemilu-minescence(ECL) western blotting detection system은 Advansta사에서 구입하였으며, 흡광도는 BioRad사의 microplate reader를 이용하여 측정하였다.
추출 − 건조된 꾸지뽕나무 뿌리 600 g을 70% 에탄올로 용 매 6L를 넣어 추출, 여과하고 rotary vacuum evaporator로 농축하여 추출물(CTE) 118.12 g을 얻었다. 시료는 -80oC에 서 보관하여 사용하였다.
세포배양 − 마우스 대식세포인 RAW 264.7 세포주는 American Type Culutre Collection(ATCC)사에서 구매하여 사용하였으며, DMEM 배지에 10% FBS와 1% penicillin streptomycin을 첨가하여 37oC, 5% CO2 인큐베이터에서 1 일 마다 계대배양 하면서 실험에 사용하였다.
세포 독성 평가 − CTE의 RAW 264.7 세포주 독성을 확 인하기 위해 꾸지뽕나무 뿌리의 농도를 1, 2.5, 5, 10 µg/mL 로 처리하여 MTT assay를 실행하였다. 96-well plate에 1×106cells/well로 seeding하여 37oC, 5% CO2 인큐베이터에 1일 동안 배양시키고, MTT 시약 처리 4시간 후, 제거하고 DMSO 200 µL를 첨가하여 microplate reader를 통해 측정 하였다.
파골세포 생성 측정 − RAW 264.7 세포주를 α-MEM에 배양하여 CTE 1, 2.5, 5, 10 µg/mL의 농도로 전처리하고 RANKL 50 ng/mL로 처리하여 파골세포로 분화시킨 후, 성 숙한 파골세포의 발현 marker로 알려진 TRAP을 염색하여 TRAP-postive한 세포를 확인하였다. 분화시킨 파골세포를 10% 포름알데히드로 10분 간 고정시킨 후 TRAP solution 을 통해 염색하였다. 그 중 3개 이상의 핵을 함유하는 TRAP- postive 다핵 세포를 현미경을 통해 수를 세어 파골세포 생 성지표로 삼았다.
Actin Ring 형성 측정 − RAW 264.7 세포주를 α-MEM 에 배양하여 CTE 1, 2.5, 5, 10 µg/mL의 농도로 전처리하 고 RANKL 50 ng/mL로 처리하여 파골세포로 분화시킨 후, 성숙한 파골세포를 Alexa Flour 488이 결합된 phallodin으로 염색시킨 후, 30분 동안 DAPI로 염색하였다. Actin ring과 DAPI 염색은 형광현미경으로 관찰하였다.
Western Blot 분석 − RAW 264.7 세포주를 6-well plate 에 1×106 cells/well, 5×103 cells/well로 각각 seeding하여 24 시간 배양시킨 후, CTE를 농도별로 전처리한 후 RANKL (100 ng/mL)을 처리하였다. 상등액을 제거하고 PBS로 세척 한 후, RIPA buffer를 첨가하여 세포를 용해시키고 원심 분 리하여 얻은 단백질은 Bradford assay로 정량하여 12%, 10%
SDS-PAGE를 이용하여 전기영동 한 후 PVDF membrane에 옮긴 다음 5% skim milk로 30분간 blocking한 후 1차 항체 를 1:1000으로 희석하여 넣고 12시간 동안 반응한 다음, 다 시 2차 항체를 반응시키고, ECL 용액을 1:1로 잘 섞어서 membrane 위에 가하여 발광시키고 LAS4000을 이용하여 확인하였다.
통계처리 − 본 실험의 통계처리는 GraphPad Prism, version 3.03(GraphPad Software Inc, San Diego, CA, USA) 을 사용하였다. 각 실험군의 결과는 평균치와 표준오차로 나타내었으며, 각 실험군 간의 결과는 student’s t-test를 사 용하여 분석하고 유의적인 차이가 있는 항목에 대해서만 검 정하였다. 실험군 간의 차이는 95% 수준(P<0.05)에서 유의 성 있는 것으로 하였다.
결과 및 고찰
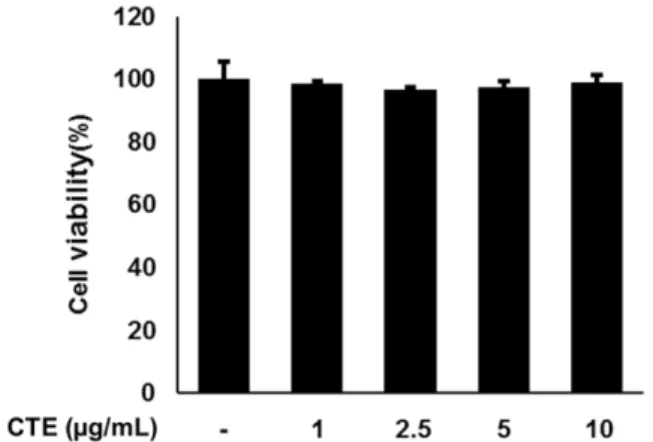
CTE의 세포 독성 효과 − CTE의 RAW 264.7 세포주에서 세포 생존에 미치는 영향을 확인하기 위해 CTE를 농도별 로 처리하여 세포 독성을 확인하였다(Fig. 1). 관찰한 결과,
Fig. 1. Viability of CTE treated in RAW 264.7 cells. Incu- bated RAW 264.7 cells with CTE (1, 2.5, 5 and 10 µg/mL) for 24 h and performed MTT assay.
24시간 배양 시 모든 농도에서 세포 독성은 나타나지 않았다.
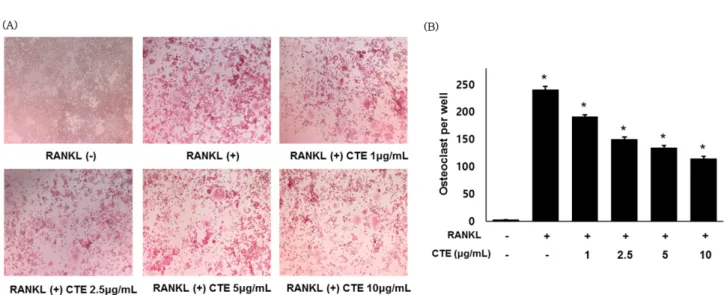
CTE의 파골세포 분화 억제 효과 − RAW 264.7 세포주에 CTE와 RANKL(50 ng/mL)을 5일 동안 처리한 후, TRAP 염색을 하여 현미경으로 관찰한 결과 농도 의존적으로 TRAP positive cell의 크기가 감소함을 확인할 수 있었다(Fig. 2A).
그리고 TRAP positive cell 수를 측정한 결과 마찬가지로 농 도 의존적으로 억제됨이 관찰되었다(Fig. 2B).
CTE의 Actin Ring 형성 − 파골세포는 뼈 흡수 과정동안 골격형성을 수행하며 뼈에 부착를 하면서, 일반적인 세포 외 공간을 구분하기 위해 파골세포의 actin이 하나의 큰 ring으 로 조직화되므로, actin ring의 형성은 파골세포의 뼈 흡수 능력에 대한 중요한 표지이다.13) 분화된 파골세포에 CTE를 농도별로 처리하여 actin ring 형성을 관찰한 결과, 농도 의 존적으로 형성이 억제됨을 확인하였다(Fig. 3).
파골세포 분화의 신호 전달 경로 − 파골세포 분화 시 RANKL/RANK 상호작용에 의해 다양한 신호전달 체계를 경유하여 조절되는데,10) 어떠한 경로를 경유한 것인지 확인 하기 위해 western blot을 통해 확인한 결과 CTE는 JNK, p38의 인산화를 억제하는 반면, ERK의 인산화는 억제하지 못하였다(Fig. 4). 이로써, CTE에 의한 파골세포 분화 억제 경로는 JNK, p38임을 증명하였다.
c-Fos와 NFATc1 발현 − c-Fos와 NFATc1은 파골세포 분 화 필수 전사인자로 파골세포 분화에 가장 핵심적인 요소 이다.12) CTE의 파골세포 분화 억제 효과를 검증하기 위해 western blot을 통해 확인한 결과 농도 의존적으로 c-Fos와 NFATc1의 발현이 감소되었다(Fig. 5). 이 결과로, c-Fos와 NFATc1의 발현 억제가 CTE의 파골세포 분화 억제 작용 기 전과 관련 있음을 증명하였다.
Fig. 2. Effect of CTE on RANKL-induced osteoclast differentiation in RAW 264.7 cells. (A) RAW 264.7 cells were treated with CTE (1, 2.5, 5 and 10 µg/mL) followed by RANKL (50 ng/mL) stimulation for 5 days and then stained for TRAP. (B) The number of TRAP-positive multinuclear cells. *p<0.05 vs. used positive control.
Fig. 3. Effect of CTE on RANKL-induced actin ring formation and nuclear staining by using DAPI. RAW 264.7 cells were treated with CTE (1, 2.5, 5 and 10 µg/mL) in presence of RANKL (50 ng/mL) for 5 days. (A) After fixation incubated with Alex Flour 488-conjugated Phallodin or (B) stained with DAPI.
결 론
본 연구는 꾸지뽕나무 뿌리 70% 에탄올 추출물의 파골세 포 분화 억제와 그 메커니즘에 관하여 연구하였으며, 그 결 과 RAW 264.7 세포주를 RANKL로 유도하여 파골세포로 분화시킨 후, 꾸지뽕나무 뿌리 70% 에탄올 추출물을 처리 하였을 때, 파골세포 분화가 억제됨을 TRAP, actin ring 형 성을 통해 확인할 수 있었으며, 분화된 파골세포에서 JNK, p38의 인산화를 통해서 억제됨을 나타내었고, 또한 파골세
포 분화의 필수 전사인자인 c-Fos와 NFATc1의 발현이 억 제됨을 확인하였다.
이상의 연구 결과로, 꾸지뽕나무 뿌리 70% 에탄올 추출 물은 RANKL로 유도된 파골세포 분화를 억제하며, 파골세 포의 분화 및 활성화에 관련된 유전자의 발현을 억제하는 것으로 나타나 골다공증의 예방 및 치료에 효과가 있을 것 이라 생각된다.
사 사
본 논문은 농림축산식품부의 재원으로 농림수산식품기술 기획평가원의 고부가가치식품기술개발사업의 지원을 받아 연구되었음(116001-03-2-HD030).
인용문헌
1. Choi, S. R., You, D. H., Kim, J. Y., Park, C. B., Kim, D. Y.
and Ryu, J. (2009) Antioxidant activity of methanol extracts from Cudrania tricuspidata Bureau according to harvesting parts and time. Korean J. Medicianl Crop. Sci. 17: 115-120.
2. Kim, O. K., Ho, J. N., Nam, D. E., Jun, W. J., Hwang, K. T., Kang, J. E., Chae, O. S. and Lee, J. M. (2012) Hepato- protective effect of Cudrania tricuspidata extracts against oxidative damage. J. Korean Soc. Food Sci. Nutr. 41: 7-13.
3. Lee, H. J., Do, J. R., Kwon, J. H. and Kim, H. Y. (2011) Physiological activities of extracts from different parts of Cudrania tricuspidata. J. Korean Soc. Food Sci. Nutr. 40:
942-948.
4. Kwon, J. Y., Hiep, N., Kim, D. W., Hong, S. G., Guo, Y., Hwang, B. Y., Lee, H. J., Mar, W. C. and Lee, D. H. (2016) Chemical constituents isolated from the root bark of Cudra- nia tricuspidata and their potential neuroprotective effects. J.
Nat. Prod. 79: 1938-1951.
5. Geng, X. L., Yang, L. B., Zhang, C., Qin, H. and Liang, Q, D. (2015) Wogonin inhibits osteoclast differentiation by inhibiting NFATc1 translocation into the nucleus. Exp. Ther.
Med. 10: 1066-1070.
6. Goltzman, D. (2002) Discoveries, drugs and skeletal dis- orders. Nat. Rev. Drug. Discov. 1: 784-796.
7. Nie, S. B., Xu, J. W., Zhang, C. H., Xu, C., Liu, M. and Yu, D. G. (2015) Salicortin inhibits osteoclast differentiation and bone resorption by down-reuglating JNK and NF-κB/
NFATc1 signaling pathways. Biochem. Biophys. Res. Com- mun. 470: 61-67.
8. Wu, Y. B., Zheng, C. J., Qin, J. P., Sun, L. N., Han, T., Jiao, L., Zhang, Q. Y. and Wu, J. Z. (2009) Antiosteoporotic activ- ity of anthraquinones from Morinda officinalis on osteoblasts and osteoclasts. Molecules 1: 573-583.
9. Murakiami, H., Takahashi, N., Tanaka, S., Nakamura, I., Udagawa, N., Nakajo, S., Nakaya, K., Abe, M., Yuda, Y., Fig. 4. Effect of CTE on RANKL-induced expression of
MAPK pathways. (A) RAW 264.7 cells were treated with CTE (1, 2.5, 5 and 10 µg/mL) for 1 h, and then treated with RANKL (100 ng/mL) for 30 min. Then, total protein was extracted and subjected to western blot analysis. CTE down- regulates the phosphorylation of JNK and p38 induced by RANKL.
Fig. 5. Effect of CTE on RANKL-induced c-Fos and NFATc1.
(A), (B) RAW 264.7 cells were treated with CTE (1, 2.5, 5 and 10 µg/mL) for 1 h and then treated with RANKL (100 ng/
mL) for 24 h. Total protein extracted and specific antibodies against c-Fos and NFATc1 subjected to western blot analysis.
Cells were lysed for western blotting with specific antibodies against c-Fos and NFATc1.
Konno, F., Barbier, A. and Suda, T. (1997) Tiludronate inhib- its protein tyrosine phosphatase activity in osteoclasts. Bone 20: 399-404.
10. Hadjdaksis, D. J. and Androulakis, I. I. (2006) Bone remod- eling. Ann. N. Y. Acad. Sci. 1092: 385-396.
11. Li, C. H., Yang, Z. F., Li, Z. X., Ma, Y., Zhang, L. P. Zheng, C. B., Qiu, W. W., Wu, X. A., Wang, X., Li, H., Tang, J., Qian, M., Li, D., Wang, P., Luo, J. A. and Liu, M. G. (2011) Maslinic acid suppresses osteoclastogenesis and prevents ovariectomy-induced bone loss by regulating RANKL-medi-
ated NF-κB and MAPK signaling pathways. J. Bone Miner.
Res. 26: 644-656.
12. Lee, J. H., Jin, H. X., Shim, H. E., Kim, H. N., Ha, H. I. and Lee, Z. H. (2010) Epigallocatechin-3-gallate inhibits osteo- clastogenesis by down-regulating c-Fos expression and sup- pressing the nuclear factor-κB signal. E. Mol. Pharmacol. 77:
17-25.
13. Teitelbaum, S. L. (2007) Osteoclasts: What do they do how do they do it? Am. J. Pathol. 170: 427-435.
(2017. 6. 10 접수; 2017. 6. 15 심사; 2017. 6. 20 게재확정)