118 책임저자:김군도, 608-737, 부산시 남구 대연3동 599-1번지
부경대학교 자연과학대학 미생물학과 Tel: 051-629-5618, Fax: 051-629-5619 E-mail: [email protected]
접수일:2010년 5월 20일, 1차수정일: 2010년 5월 25일, 2차수정일: 2010년 5월 28일, 게재승인일:2010년 5월 31일
Correspondence to:Gun-Do Kim
Department of Microbiology, College of Natural Sciences, Pukyung National University, 599-1, Daeyeon 3-dong, Nam-gu, Busan 608-737, Korea
Tel: +82-51-629-5618, Fax: +82-51-629-5619 E-mail: [email protected]
5-Aza-2’-deoxycytidine Induces Cell Cycle Arrest and the Cytoskeletal Reorganization by Recovery of Chimaerin2 in 267B1/K-ras Human Prostate Cancer Cells
Ji Young Lim1, Sook Jung Jeong2, Bo Yeon Kim2 and Gun-Do Kim1
Department of Microbiology, College of Natural Sciences, Pukyong National University, Busan 608-737,
Korea Research Institute of Bioscience and Biotechnology, Daejeon 305-806, Korea
5Aza2'deoxycytidine, a demethylating agent, induced recovery of the expression of tumor suppressor genes. Chimaerin2 among these genes down-regulated the expression of Rac, WAVE2, Arp2/3, and Villin1 in Racdependent cytoskeletal pathway and led to the changes of cell morphology, actin dynamics and cell migration in 267B1/Kras. 5-Aza-2'-deoxycytidine, also, caused cell cycle arrest at G1 to S phase by the inhibition of the expression of Cyclin-dependent kinases (CDKs), and Cdc25A. The results represent that 5-Aza-2'-deoxycytidine causes cell cycle arrest and regulates Rac-dependent actin polymerization on 267B1/K-ras human prostate cancer cells. (Cancer Prev Res 15, 118-126, 2010)
Key Words: 5-Aza-2’-deoxycytidine, Chimaerin2, Villin1, Cell cycle arrest, Rac
INTRODUCTION
Development of epigenomic technologies have made it possible to establish epigenetic differences between normal and cancer cells, thereby enhancing our understanding of the development of the disease.1)
Tumor specific methylation may provide means for detection and early diagnosis of cancer. If methylation of CpG islands is a critical parameter in tumor maintenance or progression, it would be desirable to reverse DNA hypermethylation.2,3) Changes in DNA methylation patterns are one of the most frequent events that occur in human tumors and altered CpG methylation patterns discriminate tumor tissue from its nonmalignant counterpart tissue or normal adjacent tissue.4) Therefore, demethylating agents are needed on epigenetic therapies for re-establish the expression of silenced tumor suppressor genes and clinical use against some cancers. The
cytosine methylation is one of most well studied modification in epigenetics.5)
5-Aza-2'-deoxycytidine, an inhibitor of DNA cytosine methyltransferases, is associated with DNA and DNA methyl- transferases and has been applied for epigenetic therapies poten- tially. Deoxynucleoside analogues as 5-Aza-2'-deoxycytidine are converted into the triphosphate inside S-phase cell and incorporated in place of base into DNA. Once in DNA, fraudulent bases form covalent bonds with DNA methyl- transferases, resulting in the depleting of active enzymes and demethylation of DNA.6) It has been reported to induce cell cycle arrest and have anti-cancer activities and shown to have significant cytotoxic and antineoplastic activities in many experimental tumors.7,8)
Also, the expression of several tumor suppressor genes have been changed by the treatment of 5-Aza-2'-deoxycytidine. Chi- maerin 2 (CHN2) of these genes is family of GTPase-activating proteins (GAPs) that is regulated by the lipid diacylglycerol
(DAG). Reducing of chimaerins in various types of cancers is important evidence about chimaerins as a tumor suppressor.9)
The interaction between Rac and chimaerin in cancer signaling is intuitive, based on the role of chimaerins to inhibit Rac that itself shows increased activity in a multiple human carcinomas.10∼14)
It is widely accepted that Rac and other Rho GTPase play critical roles in the regulation of cell morphology and movement, invasion, proliferation and malignant which are all crucial events in cancer development and progression.15,16) Cells move in response to signals which eventually affect cytoskeletal and adhesive structures of the cell.17) Typically, Rac regulates lamellipodia formation through activation of the phospholipid metabolism that generates PI(4,5)P2, which in turn regulates several actin binding proteins such as WAVE, Arp2/3 and gelsolin. Lamellipodia is large veil-like sheets, which contain highly branched and cross-linked actin filament. Also, it is generally associated with cell migration.18)
This study showed the changes in 267B1 human prostate cancer cells transformed by K-ras after treatments of 5-Aza- 2'-deoxycytidine. These changes, particularly, are confirmed in the aspects of cell cycle arrest and regulation of cytoskeleton associated with Rac.
MATERIALS AND METHODS 1. Cell culture and treatment
Human epithelial cell line (267B1) established from fetal prostate tissue can be malignantly transformed by a biological carcinoma, and can serve as a useful model for investigations of the progression steps of carcinogenesis. Activated K-ras was introduced into 267B1 cells by infection with the Kirsten murine sarcoma virus.19,20) These cells are immortalized and contained the essential characteristics of primary human prostate epithelial cells such as morphology, expression of cytokeratins.21) The transformed 267B1/K-ras human prostate cancer cells were used for the experiments. The cells was cultured in RPMI-1640 media (Hyclone, Logan, UT, USA) containing 10% heat-inactivated fetal bovine serum (FBS) and 100 U/ml penicillin and 100μg/ml streptomycin (PAA Laboratories GmbH, PA, Austria) at 37oC in a humidified atmosphere of 5% CO2. For the treatment, 5-Aza-2'-deoxy- cytidine (Sigma-Aldrich, St. Louis, MO, USA) was dissolved in dimethyl sulfoxide, DMSO (Fluka, Milwaukee, WI, USA) and
then kept at −20oC until required. 5-Aza-2'-deoxycytidine was enhanced by changing the media every 24 h up to 72 h. The final concentration of DMSO in the culture medium did not exceed 0.04% (v/v), and the same concentration of DMSO was added to the control dishes.
2. Cell proliferation assay
For the cell proliferation assay, 1×104 cells were resuspended in 100μl of medium and seeded onto each well of a 96-well plate. Various concentrations of 5-Aza-2’-deoxycytidine (0, 10, 20, 50 and 100μM) were treated for 72 h and 10μl of cell viability assay solution WST-1Ⓡ (Daeil Lab Service, Jong-No, Korea) was added, and further incubated for 3 h. The proliferation inhibitory rates were measured at 460 nm with ELISA reader (Molecular Devices, Silicon Valley, CA, USA).
3. Analysis of gene expression
Expression levels of selected tumor suppressor genes were examined by real-time PCR. mRNA purified by RNesay column (Qiagen, Valencia, CA, USA) was used for reverse transcription PCR using MuLV reverse transcriptase. Quanti- tative assessments of DNA amplification were performed by fluorescence-based real-time detection system (Takara Bio Inc., Shiga, Japan) using SYBR green.
4. Cell cycle analysis
Cells were treated with 5-Aza-2’-deoxycytidine for 12 h and then harvested after trypsinization. Cells were washed with PBS and centrifuged at 1,800 rpm for 3 min, with cold ethanol overnight at 4oC. RNase (10μg/ml) was treated and incubated at 37oC for 1 h. Then the cells were stained with 10μl of propidium iodide (Sigma-Aldrich). The distribution of various stage of the cell cycle was analysed by flow cytometry (Becton-Dickinson, San Jose, CA, USA).
5. Western blotting
For western blot analysis, the treated 267B1/K-ras cells were washed with ice-cold PBS and removed by scrapping. The cells were collected by centrifugation and lysed in ice-cold lysis buffer (PRO-PREPTM, Intron Biotech., Gyeonggi, Korea) containing proteinase inhibitors. After incubation on ice for 30 min, the proteins were collected by centrifugation at 14,000 rpm for 20 min at 4oC. The protein quantity of the cell lysates were determined by a Protein Quantification Kit (CBB solutionⓇ)
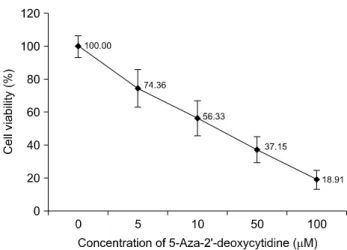
Fig. 1. The effects of 5-Aza-2'-deoxycytidine on cell proli- feration. Proliferation inhibition rates of 5-Aza-2'-deoxycytidine on 267B1/K-ras cells determined by MTT assay. The cells were treated with indicated concentrations of 5-Aza-2'-deoxy- cytidine for 72 h. 5-Aza-2'-deoxycytidine treatment inhibited the growth of 267B1/K-ras prostate cancer cells. The values from the assay were compared to those from untreated control as percentage (mean±SD) of at least three independent experiments.
(Dojindo Molecular Technologies, Rockvile, MD, USA) with bovine serum albumin (BSA) as standard. An aliquot from each sample were boiled for 4 min and then resolved by 12%
SDS-PAGE. The proteins were electrotransferred to a cellulose nitrate membrane (Schleicher & Schuell, Keene, NH, USA).
After transfer, the membrane was blocked in PBST buffer (135 mM Sodium chloride, 2.7 mM Potassium chloride, 4.3 mM Sodium phosphate, 1.4 mM Potassium dihydrogen phosphate, 0.5% Tween-20) containing 5% skim milk for overnight at 4oC. The blots were probed with the primary antibody (Cell signaling, Beverly, MA, USA) overnight at 4oC. After the reaction with the primary antibody, the membrane was washed and incubated with HRP conjugated IgG as second antibody (Cell signaling) for 1 h. The blots were then washed with PBST buffer and the signals were visualized by ECL detection system (Pierce, Rockford, IL, USA).
6. Immunofluorescence of cytoskeletal protein
Cells were cultured (at 37oC, 5% CO2) on coverglassBottom dishes (SPL Lifesciences, Gyeonggi, Korea) in RPMI1640 media (Hyclone, Logan, UT, USA) containing 10% FBS and penicillinstreptomycin (penicillin: 100μg/ml, streptomycin:
100 units/ml). (Hyclone, Logan, UT, USA) for 72 h. For the experiment, cells were fixed with 4% formaldehyde (Junsei Chemical Ltd., JAPAN) for 15 min at room temperature and then blocked for 1 h in 5% normal serum of the host against primary antibodies and 0.3% Triton X100. The fixed and blocked cells were incubated with 0.1μg/ml of primary antibodies (antiWAVE2 and antiVillin1) (Cell Signaling Technology Inc., Beverly, MA, USA) for 3 h and then with 0.1μg/ml of antirabbit IgG (H+L), F(ab') fragment (Alexa Fluor 488 conjugate) for 1 h. Stained cells on the slides were mounted in Prolong Gold Antifade Reagent (Invitrogen, Eugene, OR, USA) followed by observation under a Nikon ECLIPS 50i microscope equipped with chargedcoupled device (CDD) camera. Images were captured and processed with HighContent Analysis Software (Cambridge Healthtech Insti., Needham, MA, USA).
RESULTS
1. Effects of 5-Aza-2'-deoxycytidine on cell proliferation
The results of MTT assay showed that 5-Aza-2’-deoxcycytidine
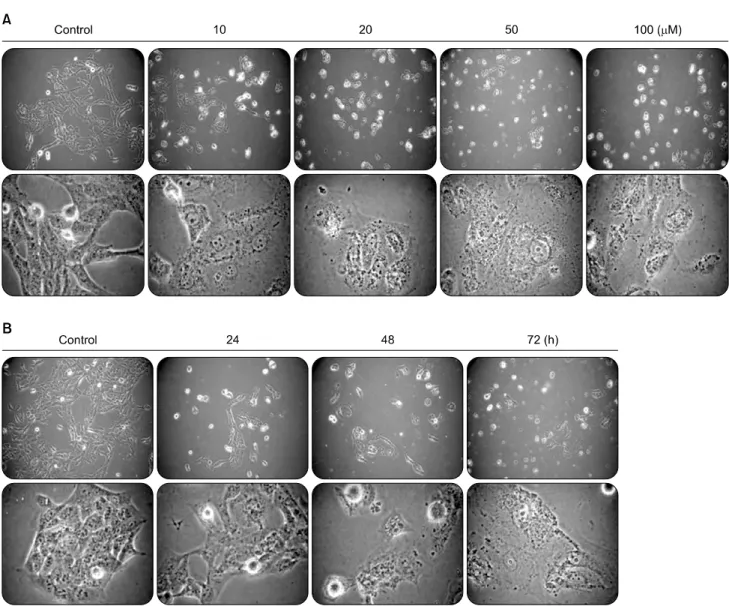
inhibited proliferation of 267B1/K-ras cells after 72 h of treat- ment. The observed inhibitory effect was in dose-dependent manner. The IC50 value of 5-Aza-2’-deoxycytidine was 10μM (Fig. 1). Additionally, the effects of higher concentration of 5-Aza-2’-deoxycytidine and different treatment time were examined (Fig. 2). Most of the cells were dead and floated under higher concentration. Thus, the IC50 value, 10μM of 5-Aza-2’-deoxycytidine was used in subsequent experiments.
2. Effects of 5-Aza-2’-deoxycytidine on cell cycle
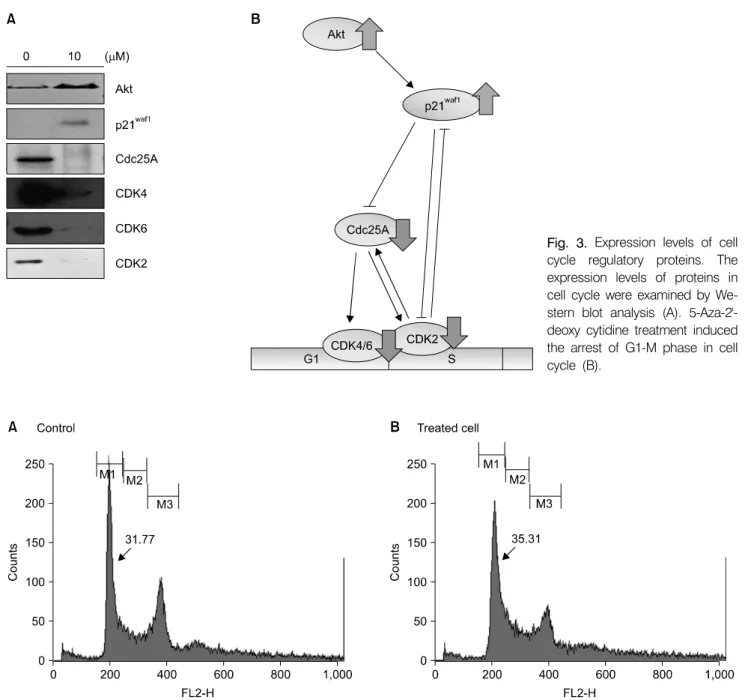
When the cells were treated with 10 µM of 5-Aza-2’-deo- xycytidine for 72 h, it leads to the expression of cell cycle regulator proteins such as Cyclin-dependent kinases (CDKs).
Especially, the expression of Cdc25A was completely arrested (Fig. 3A). Cdc25A regulates the G1 to S phase transition by dephosphorylating and activating the Cdk2/cyclinE and Cdk2/CyclinA complexes. The decreased expression of CDK4, CDK6 and Cdc25A were confirmed the arrest of cell cycle progression. In order to identify the upstream signal controller, the expression level of p21waf1, inhibitor of CDK, was examined and its expression was increased in the treated cells. It has also been detected that 5-Aza-2’-deoxycytidine induced the
Fig. 2. Morphological changes in different treatment conditions of 5-Aza-2'-deoxycytidine. Morphological changes of 267B1/K-ras cells depend on dose (A) and treatment time (B) of 5-Aza-2'-deoxycytidine. On panels A and B, the above pictures are magnified to 100 times, and others in the bottom are 400 times by a phase microscope.
expression of Akt (Protein kinase B) in the cells. A proposed pathway of cell cycle control by 5-Aza-2’-deoxycytidine in 267B1/K-ras cells was shown in Fig. 3B. Flow cytometry analysis showed the most of the cells accumulated in the G1 phase and then the populations were decreased in the G2/M phase (Fig. 4).
3. 5-Aza-2'-deoxycytidine induced expression of tumor suppressor genes
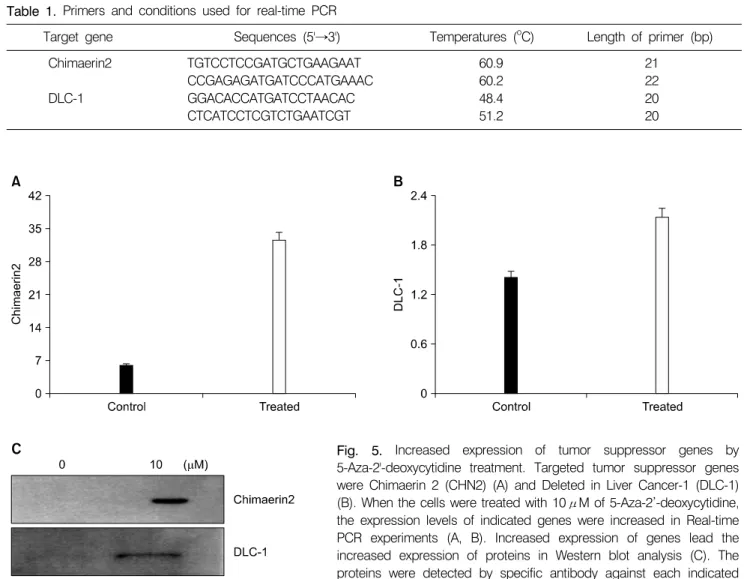
Inhibition of DNA methylation by 5-Aza-2’-deoxycytidine may induce the expression of tumor suppressor genes. To confirm the recovery expression of inhibited tumor suppressor genes, real-time PCR analysis was performed. Chimaerin (CHN2) and Deleted in Liver cancer 1 (DLC-1) were the
targeted genes for this analysis. Primer sequences and annealing temperatures are indicated in Table 1. The results showed that the expression levels of DLC-1 and CHN2 genes were increased after the treatment (Fig. 5A, B). The same results were noticed in protein level through Western blot analysis (Fig. 5C). The results confirmed 5-Aza-2’-deoxycytidine induces up-regulated expression of the inhibited tumor suppressor genes in both gene and protein levels.
4. Increased expression of Chimaerin2 by 5- Aza-2'-deoxycytidine induced the change of Rac-dependent cytoskeleton
Dynamic changes of cytoskeleton are basis in cellular
Fig. 3. Expression levels of cell cycle regulatory proteins. The expression levels of proteins in cell cycle were examined by We- stern blot analysis (A). 5-Aza-2'- deoxy cytidine treatment induced the arrest of G1-M phase in cell cycle (B).
Fig. 4. FACS analysis for the cell cycle of 267B1/K-ras cells treated with 10μM 5-Aza-2’-deoxycytidine. The degree of cell cycle arrest by 5-Aza-2’-deoxycytidine was analyzed by flow cytometry. G1 (M1) areas of cells treated with 5-Aza-2’-deoxycytidine (B) increased compare to the untreated cells (A). Increasing of G1 (M1) areas by treatment of 5-Aza-2'-deoxycytidine shows the arrest of G1 phase in Cell cycle. The result is from one representative experiment of three independent experiments that shown the similar patterns.
reactions including cell migration, motility and cancer invasion.
Motility and invasion are induced in pseudopodia such as lamellipodia, filopodia by changing in cytoskeleton. It has been known that Rac and other Rho GTPases play critical roles in cancer invasion and motility.21) Chimaerin2 increased in both real-time PCR and western blot analysis and it has been widely accepted as an inhibitor to Rac. As shown in Fig. 6, the
expression of Rac was decreased when 5-Aza-2'-deoxycytidine was treated at 10μM for 72 h. Based on these results, the WASP/WAVE family of actin cytoskeletal regulatory protein, WAVE2 and Actin-Realted Protein, Arp2/3 and actin-binding protein, Villin1 were checked in the expression levels of protein by western blot analysis (Fig. 6A). In the same condition, increasing of Chimaerin2 inhibited Rac-dependent pathway
Table 1. Primers and conditions used for real-time PCR
Target gene Sequences (5'→3') Temperatures (oC) Length of primer (bp)
Chimaerin2 TGTCCTCCGATGCTGAAGAAT 60.9 21
CCGAGAGATGATCCCATGAAAC 60.2 22
DLC-1 GGACACCATGATCCTAACAC 48.4 20
CTCATCCTCGTCTGAATCGT 51.2 20
(Fig. 6B) and led the changes of cell morphology such as indistinct cell membrane and cell adjunction (Fig. 7). The results suggested that the recovery of Chimaerin2 by 5-Aza-2'- deoxycytidine treatments may control the pseudopodia such as lamellipodia and finally invasion of cancer cells can be regulated.
DISCUSSION AND CONCLUSION
Because epigenetic therapies have raised considerable interests in cancer, 5-Aza-2’-deoxycytidine was targeted and studied.
Many studies have reported that 5-Aza-2’-deoxycytidine treat- ment inhibited Dnmts and induced cell proliferation, and apoptosis in tumors for cancer therapies, but their exact in vivo mechanism was required for more studies and results.
In this study, the results showed that the 5-Aza-2’-deo- xycytidine inhibited the cell proliferation in dose dependent manner. The cell proliferation was arrested at the inhibitory concentration of 10μM and the exposure time was 72 h. At the 10μM concentration, most of the cells were dead and floated.
The progression of cell proliferation encounters the cell cycle check points. Cyclin-dependent kinases (CDKs) are an internal part of the cell cycle control machinery that regulates the proliferation of eukaryotic cells. Inhibition of Cdc25A by 5-Aza-2’-deoxycytidine arrests the G1-S phase transition.
CDK4, CDK6 and CDK2 are important regulators in the control of that how cells leave the quiescent state and transgress through the G0/G1 restriction point. The essential role of p21Waf1 relies upon its well known ability to inhibit Fig. 5. Increased expression of tumor suppressor genes by 5-Aza-2'-deoxycytidine treatment. Targeted tumor suppressor genes were Chimaerin 2 (CHN2) (A) and Deleted in Liver Cancer-1 (DLC-1) (B). When the cells were treated with 10μM of 5-Aza-2’-deoxycytidine, the expression levels of indicated genes were increased in Real-time PCR experiments (A, B). Increased expression of genes lead the increased expression of proteins in Western blot analysis (C). The proteins were detected by specific antibody against each indicated protein.
Fig. 6. Proposed signaling cas- cades of cytoskeleton modifica- tion mediated by Chimaerin2. The expression levels of proteins in Rac-dependent cytoskeleton mo- dification were examined by Western blot analysis (A). In- creased expression of Chimaerin2 by 5-Aza-2'-deoxycytidine treat- ment regulates the expression of each component in Rac-depen- dent cytoskeleton modification cascade (B).
CDK and DNA replication, thereby inducing cell cycle arrest.
Also, p21Waf1 is recruited to the Cdc25A for inhibiting its activation.22)
Cdc25A is a tyrosine phosphatase that functions as a key regulator in cell cycle progression and DNA damage response in eukaryotes. Cdc25A regulates the G1 to S phase transition by dephosphorylating and activating the Cdk/CyclinE and Cdk2/CyclinA complexs.23) Also, Cdc25A has been reported that the expression is up-regulated in human prostate cancer.24) Flow cytometry result accumulation of cells in G1 phase and decreased population in G2 phase.
Inhibition of DNA methylation, ultimately induced the expression of tumor suppressor genes. Real-time PCR analysis showed the higher expression of Chimaerin2 and Deleted in Liver Cancer-1 (DLC-1) after the treatment. The expression of Chimaerin2 was remarkably increased and led the regulation of cytoskeleton on Rac-dependent manner. It acts as GTPase-activating protein (GAP) for Rac and inactivates Rac signaling, cell proliferation and cytoskeletal reorganization by promoting the GDP-bound form and as tumor suppressor gene. Many studies have proposed a role for Rac in the control of mitogenesis through its ability to regulate G1-S transition and cyclin D1 expression. Moreover, Rac and other members of the Rho GTPase family are over-expressed in human tumors and hypermethylation of Rac leading to a higher rate of cell proliferation has been found in cellular models of human cancer.25)
Similarly, downstream molecules such as Wiscott-Aldrich Syndrome protein (WAVE), Actin-related protein (Arp) and Villin in Rac-dependent pathway were examined in the experiment. WAVE mediates actin dynamics by activating the Arp2/3 actin nucleation complex in response to activated Rho family GTPase. Villin of a member of gelsolin family has function about regulation of cell shape in response to external stimulation. Increased expression of these genes were seen in breast and colorectal cancers.26,27) Cytoskeletal changes are the key mechanism for metastasis in cancer cells. Regulation of Rac-dependent pathway by 5-Aza-2’-deoxycytidine represents potential possibility besides inhibition of cell proliferation and induction of apoptosis.
In conclusion, the results of the present study showed that the anti-cancer effect of 5-Aza-2’-deoxycytidine in the arrest of the cell cycle, and regulation of cytoskeleton by recovering expression of Chimaerin2 in 267B1/K-ras human prostate cancer cells. This study may offer possible molecular bases for further research in the action of methylation on other human cancer.
ACKNOWLEDGMENT
This work was supported by the Research Program for New Drug Target Discovery (M10748000346-07N4800-34610) from the Ministry of Education, Science & Technology, Republic of Korea.
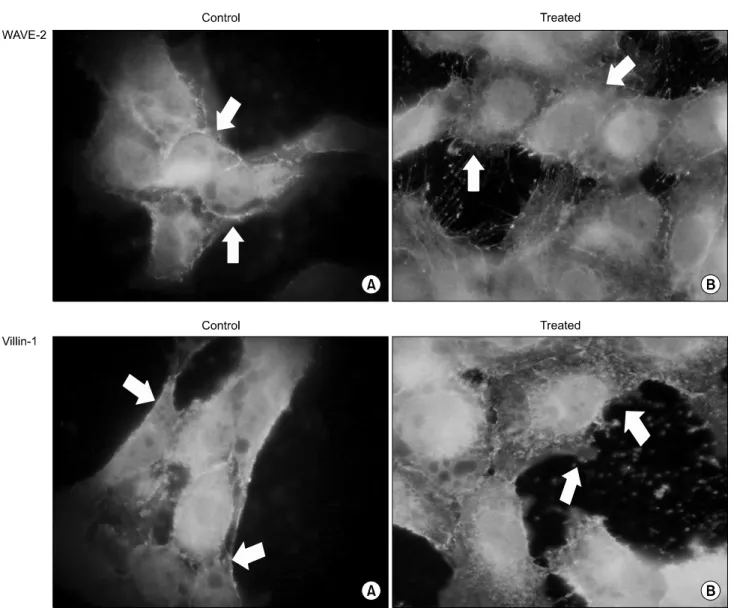
Fig. 7. Immunofluorescence of WAVE2 and Villin1 on cytoskeletal modification. In Rac-dependent cytoskeletal modification pathway, WAVE2 and Villin1 effected on the changes of cell morphology. As shown on control (A) and treated cells (B), decreased expression levels of WAVE2 and Villin1 resulted in the shape of cell is indistinct and adjunction of cell to cell was come off. Arrows are indicated the differences on cytosckeleton of the cells.
REFERENCES
1) Frage MF, Esteller M. DNA methylation: a profile of methods and applications. Biotechnigues 33, 632-636, 2002.
2) Pfeifer GP, Rauch TA. DNA methylation patterns in lung carcinomas. Seminars in Cancer Biology 19, 181-187, 2009.
3) Yen RW, Vertino PM, Nelkin BD, Yu JJ, el-Deiry W, Cumaraswamy A, Lennon GG, Trask BJ, Celano P, Baylin SB. Isolation and characterization of the cDNA encoding human DNA methyltrasnferase. Nucleic Acids Res 20, 2287-2291, 1992.
4) Jones PA, Baylin SB. The epigenomics of cancer. Cell 92,
128-683, 2007.
5) Mulero-Navarro S, Esteller M. Epigenetic biomarkers for human cancer: the time is now. Critical Reviews in Oncology/
Hematology 68, 1-11, 2008.
6) Egger G, Liang G, Aparicio A, Jones PA. Epigenetics in human disease and prospects for epigenetic therapy. Nature 429, 457-463, 2004.
7) Bender CM, Zingg JM, Jones PA. DNA methylation as a target for drug design. Pharm Res 5, 175-187, 1998.
8) Natsume A, Wakabayashi T, Tsujimura K, Shimato S, Ito M, Kuzushima K, Kondo Y, Sekido Y, Kawatsura H, Narita Y, Yoshida J. The demethylating agent 5-Aza-2'-deoxycytidine activates NY-ESO-1 antigenicity in orthotopic human glioma.
Int J Cancer 122, 2542-2553, 2008.
9) Bruinsma SP, Baranski TJ. β2-Chimaerin in Cancer Signaling, Connecting cell adhesion and MAP kinase activation. Cell Cycle 6, 2440-2444, 2007.
10) Burbelo P, Wellstein A, Pestell RG. Altered Rho GTPase signaling pathways in breast cancer cells. Breast Cancer Res Treat 84, 43-48, 2004.
11) Fernandez-Zapico ME, Gonzalez-Pan NC, Welss E, Savoy DN, Molina JR, Fonseca R, Smyrk TC, Chari ST, Urrutia R, Billadequ DD. Ectopic expression of VAV1 reveals an unexpected role in pancreatic cancer tumorigenesis. Cancer Cell 7, 39-49, 2005.
12) Jordan BA, Devi LA. G-protein coupled receptor heterodi- merization modulates receptor function. Nature 700, 339-697, 1999.
13) Lozano E, Betson M, Braga VMM. Tumor progression: Small GTPase and loss of cell-cell adhesion. BioEssays 25, 452-463, 2003.
14) Sahai E, Marshall CJ. Rho-GTPases and cancer. Nat Rev Cancer 2, 133-142, 2002.
15) Gomez PT, Benitah SA, Valeron PF, Espina C, Lacal JC. Rho GTPase expression in tumorigenesis : evidence for a significant link. BioEssays 27, 602-613, 2005.
16) Ridley AJ. Rho proteins and cancer. Breast Cancer Res Treat 84, 13-19, 2004.
17) Lambrechts A, Van Troys M, Ampe C. The actin cytoskeleton in normal and pathological cell motility. J Biol Chem and Cell Biology 36, 1890-1909, 2004.
18) Small JV. Lamellipodia architecture : actin filament turnover and the lateral flow of actin filaments during motility. Seminars in Cell Biology 5, 157-163, 1994.
19) Kim BY, Kim KA, Kwon O, Kim SO, Kim MS, Kim BS, Oh WK, Kim GD, Jung, M, Ahn JS. NF-kB inhibition
radiosensitizes Ki-Ras-transformed cells to ionizing radiation.
Carcinogenesis 26, 1395-1403, 2005.
20) Parda DS, Thraves PJ, Kuettel MR, Lee MS, Arnstein P, Kaighn ME, Rhim JS, Dritschllo A. Neoplastic transformation of a human prostate epithelial cell line by the v-Ki-ras oncogene. The Prostate 23, 91-98, 1993.
21) Servitja JM, Marinissen MJ, Sodhi A, Bustelo XR, Gutkind JS. Rac1 function is required for Src-induced transformation.
Evidence of a role for Tiam1 and Vav2 in Rac activation by Src. J Biol Chem 278, 34339-34346, 2003.
22) Vigneron A, Cherier J, Barre B, Gamelin E, Coqueret O. The cell cycle inhibitor p21Waf1 binds to the myc and Cdc25A promoters upon DNA damage and induces transcriptional repression. J Biol Chem 281, 34742-34750, 2006.
23) Zhang P. The cell cycle and development: redundant roles of cell cycle regulators. Cell Biology 11, 655-662, 1999.
24) Chiu YT, Han, HY, Leung SC, Yuen HF, Chau CW, Guo Z, Qiu Y, Chan KW, Wang X, Wong YC, Ling MT.
Cdc25A functions as a novel ar corepressor in prostate cancer cells. J Mol Biol 285, 446-456, 2009.
25) Yang C, Liu Y, Leskow FC, Weaver VM, Kazanietz MG.
Rac-GAP-dependent inhibition of breast cancer cell prolifera- tion by β2-Chimaerin. J Bio Chem 280, 24363-24370, 2005.
26) Wang Y, Srinivasn K, Siddiqui MR, George SP, Tomar A, Khurana S. A novel role for villin intestinal epithelial cell survival and homeostasis. J Bio Chem 283, 9454-9464, 2008.
27) Sossey-Alaoui K, Su G, Malaj E, Roe B, Cowell JK. WAVE3, an actin-polymerization gene, is truncated and inactivated as a result of a constitutional t(1;13)(q21;q12) chromosome translocation in a patient with ganglioneuroblastoma. Oncogene 21, 5967-5974, 2002.