Corresponding author: Dong-Su Choi, Institute of Symbiotic Science and Technology, Tokyo University of Agriculture and Technology, Tokyo 183-8509, Japan, Tel.: +81-42-367-5241, E-mail: [email protected]
실내환경에서 자외선 조사 및 원적외선 차단이 월계수의 생장에 미치는 영향
후지와라 케이스케*․토다 히로토**․최동수**
*동경농공대학교 대학원 농학부․**동경농공대학교 농학연구원
The Effect of UV-B Irradiation and Far-red Light Reduction on the Growth of Laurus nobilis in Indoor
Fujiwara, Keisuke*․Toda, Hiroto**․Choi, Dong-Su**
*Graduate School of Agriculture, Tokyo University of Agriculture and Technology
**Institute of Symbiotic Science and Technology, Tokyo University of Agriculture and Technology
국문초록
실내 광환경은 자연 상태와 달라 실내 조경 식물의 생장 및 생리활성에 영향을 줄 것으로 생각된다. 본 연구는 광질(光 質)이 월계수의 생장 및 생리활성에 미치는 영향을 평가하기 위해 실시하였다. 월계수 묘목은 4개의 다른 광원 처리구 (control, +UV, -FR, +UV-FR 처리구)에서 180일 동안 생육시켰다. 자외선 추가 처리구(+UV)에서 생육한 월계수는 건중량 및 잎 면적이 감소되었지만, 잎 두께 및 잎내 자외선 흡수 물질의 함유량은 증가했다. 반면, 원적외선 차단 처리구(-FR)에서 생육한 월계수는 큰 변화를 나타내지 않았다. 하지만, 자외선 추가 및 원적외선 차단 처리구(+UV-FR)에서 생육한 월계수는
신장생장 및 광합성 속도가 유의적으로 저하했다(P<0.05). 이상의 결과, 광질이 실내 조경 식물인 월계수의 생장 및 생리활
성에 큰 영향을 주는 것으로 밝혀졌다. 실내 조경 식물인 월계수를 관리할 때 광질을 이용한 관리의 기초 자료로 활용될 수 있을 것이다.
주제어: 광질(光質), 광합성, 자외선 흡수 물질, 실내 조경 식물
ABSTRACT
The main purpose of this research is to evaluate the effect of light quality on the growth and physiological activities of Laurus nobilis plants growing indoors, the L. nobilisseedlings were grown under four types of lighting for 180 days.
The seedlings were grown under 4 different treatments((n=9 seedlings per treatment): control, control supplemental UV-B irradiated(+UV), FR reduced(-FR) and simultaneously supplemental UV-B irradiated and FR reduced(+UV-FR)). It was found that UV-B irradiation(+UV) reduced dry weight and leaf area, and increased leaf thickness and the amount of UV-absorbing compounds per unit leaf area. In contrast, a reduction in far-red(FR) light did not affect any of these parameters. Interestingly, however, the elongation growth and net photosynthetic rate of the L. nobilis seedlings grown under simultaneous UV-B
irradiation and FR reduction(+UV-FR) were significantly decreased than the control treatment. From these results, it is concluded that the light quality has a large effect on the indoor growth ofL. nobilis. This study can suggest basic information for managed in the L. nobilis in indoor using light quality.
Key Words: Light Quality, Photosynthesis, UV-Absorbing Compounds, Interior Plant
Ⅰ. Introduction
Light is one of the most important factors for plant growth, with photosynthetically active radiation(PAR; 400-700 nm) being necessary for photosynthesis. Plants are also exposed to ultraviolet(UV) radiation(280-400 nm) and far-red(FR) light (700-800 nm), both of which affect plant physiology and morphology(Kasperbauer, 1988; Kondo, 2010).
It has been reported in many studies that enhanced UV-B radiation reaching the surface of the earth has very many adverse impacts on plant growth parameters(e.g. causing da- mage to DNA, cell membranes, and photosynthetic tissue) (Jansen, 2002; Jansen and Borman, 2012; Searles et al., 2001). Moreover, FR light also affects plant morphology and physiology. Since green leaves absorb most of the red(R) light and reflect much of the FR light, plants can sense competition from other plants by measuring the relative amount of FR light reflected from them(Kasperbauer, 1988).
It has been found that plants growing in a low R/FR ratio environment invest their biomass into stem elongation and increased shoot length(Fukuda et al., 2002).
Today, plants are used in many indoor spaces, including domestic and commercial dwellings, botanical gardens, and plant factories(e.g.Laurus nobilis,Ficus elastica,Ficus benjamina).
However, many studies indicate that plant growth under indoor conditions is more sensitive to increases in UV-B radiation than field conditions(Rozema et al., 1997). Especially, the level of UV-B radiation in indoor is lower than in the field because it is filtered by window glass(Suzuki and Kondo, 2005) and is not provided by artificial light sources. The R/FR ratio also varies, depending on the type of artificial light source used(Fukuda et al., 2002). Consequently, indoor environments may affect both the morphology and physiology of plants. For example, it has been reported that indoor plants tend to be spindly and have over-large leaves(Oide and Kondo, 2001), which may be of particular concern for deco- rative plants, which are required to be as beautiful as possible
to maximize their aesthetic value.
L. nobilis, it is commonly grown indoor for the interior landscaping in Japan, was resistant against UV-B radiation damage, as UV-B did not affect it biomass production and allocation photochemical efficiency and chlorophyll content, and increased the thickness of leaves and cuticles in this plant(Grammatikopouloset al., 1998).
To enable us to manage the growth of indoor plants and enhance the beauty of decorative plants, it is crucial that we understand the effect of light quality on their physiology and morphology. Therefore, the aim of this study was to deter- mine the effect of light quality on the indoor growth and leaf shape of L. nobilisusing an indoor pot experiment.
Ⅱ. Materials and Methods
1. Plant Materials and Experimental Design
Seedlings ofLaurus nobiliswere planted in pots(Φ17 cm × 20 cm high) that were filled with fertile soil from the nursery of Tokyo University of Agriculture and Technology in May 2011. To evaluate the effect of UV-B irradiation and R/FR ratio on the growth and physiological characteristics of L.
nobilis, seedlings were grown under 4 different treatments for 180 days(n=9 seedlings per treatment): control supplemental UV-B irradiated(+UV) FR reduced(-FR) and simultaneously supplemental UV-B irradiated and FR reduced(+UV-FR).
The experimental light environment depended on solar PAR, with no artificial PAR being irradiated. UV-B was irradiated with a UV-B fluorescent lamp(GL15E, Sankyo Denki, Hiratsuka, Kanagawa, Japan) at 0.35 W/m2 for 8 hours per day during the experimental period, and cellulose acetate was used to remove harmful wavelengths shorter than 290 nm. Solar heat cutting film(SL999Cyber-reps, Nagareyama, Chiba, Japan) was placed on the window to reduce the FR light by approximately 40%. The R/FR ratio was estimated to be approximately 1.0 for the control and +UV treatments, and
approximately 2.0 for the -FR and +UV-FR treatments.
2. Measurement of Growth
For each seedling, six seedlings were selected from each treatment, the height was measured and the number of leaves was counted every 1-2 months. At the end of the experiment, all seedlings were harvested, and their dry weight and plant height were measured. Leaves were divided into upper and lower leaves for each seedling. Leaf area was then measured using image analysis(Image j NHI, Bethesda, MD, USA) and leaf thickness was assessed using a coolant proof micrometer(MDC-25MJ, Mitutoyo, Kawasaki, Kanagawa, Japan).
The measurements were repeated 5 times.
3. Measurement of Photosynthesis
Photosynthetic light response curves, six seedlings were selected from each treatment, were measured using a LI-6400XT portable photosynthesis system(LI-cor, Lincoln, NE, USA). The photo- synthesis photon flux density(PPFD) was changed from high to low(1,500, 1,200, 1,000, 800, 500, 300, 200, 100, 70, 50, 30, 20, 10, and 0 µmol․m-2․s-1) under controlled levels of CO2
diffusion(370 µmol․m-2․s-1), flow rate(500 µmol․s-1), leaf temperature(25℃), and relative humidity(60-70%). The data were approximated to follow a non-rectangular hyperbola (Johnson and Thornley, 1984), using the equation:
Pn=[(fI+Pmax-((fI+Pmax)2-4fIθPmax)0.5)/2θ-R]
where Pn=net photosynthetic rate, f=initial slope of the photosynthetic curve, I=light intensity, Pmax=maximum value of the photosynthetic rate, θ=convex degree of the photosynthetic curve, and R=respiration rate.
4. Leaf Nutrients
Chlorophyll was extracted from leaves using dimethyl sul- foxide(DMSO), according to Barnes et al.(1992), and the absorbance was measured using a spectrophotometer(1800U HITACHI, Chiyoda, Tokyo, Japan). The following formula was then used to calculate the approximate concentrations of chlorophyll a and b(Shinano et al., 1996):
Chl a (µg․mg-1)=(14.85A665-5.14A648)×(a/b) Chl b (µg․mg-1)=(25.48A648-7.36A665)×(a/b),
where A665=absorbance at 665 nm, A648=absorbance at 648 nm, a=volume of DMSO solution(mL), and b=dry mass of leaves(mg).
Following the measurement of photosynthetic rate, leaves were dried at 60℃ for 1 week and then ground. The nitrogen concentration in the leaves was measured using an NC analyzer(MT-700, Yanaco, Kyoto, Kuse, Japan).
To measure the amount of UV-absorbing compounds in the leaves, we followed a previously published methodology (Kanget al., 1998; Liet al., 1993). Leaves from 6 individuals per treatment were homogenized in a 10-fold volume of 50 mM sodium phosphate buffer(pH 7.2) using a pestle and mortar. An aliquot of the homogenate was then extracted with 1%(vol/vol) HCl in 70%(vol/vol) methanol(final con- centration) at room temperature for 3 h. The mixture was centrifuged at 2,900 rpm for 10 min, and the amount of UV- absorbing compounds in the supernatant was determined using a spectrophotometer(1800U HITACHI, Chiyoda, Tokyo, Japan).
5. Statistical Analysis
All statistical analyses were performed using Statistical Analysis System(SAS) programs(Version 9.1 SAS Institute Inc., Cary, NC, USA). The significance of the differences between measurements was tested with Tukey’s studentized range test, with a 0.05% probability of error.
Ⅲ. Results
1. Plant Growth
Figure 1 shows the elongation growth of Laurus nobilis under each treatment. Plants in the +UV and -FR treatment groups had elongation growth of 13.13 cm and 15.25 cm, respectively, which were not significantly different from the control group(13.88 cm). However, there was a significant decrease in elongation growth in plants in the +UV-FR treatment group(P<0.05).
Figure 2 shows the dry weight of roots, stems, and leaves.
The amount of growth in plants in the -FR treatment group (18.0 cm) was not much different from the control group.
However, +UV treatment significantly decreased the dry weight of seedlings.
Figure 1. Elongation growth during the growing period of Laurus nobilis grown under different light conditions. Values re- present the mean±SE of 6 plants. Bars with different letters are significantly different(P<0.05). See Materials and Methods for an explanation of the different treat- ments.
Figure 2. Dry weight of Laurus nobilis grown under different light conditions after harvest. Values represent the mean ± SE of 6 plants. Bars with different letters are signifi- cantly different(P<0.05). See Materials and Methods for an explanation of the different treatments.
Legend: Leaf, □ Stem, ■ Root
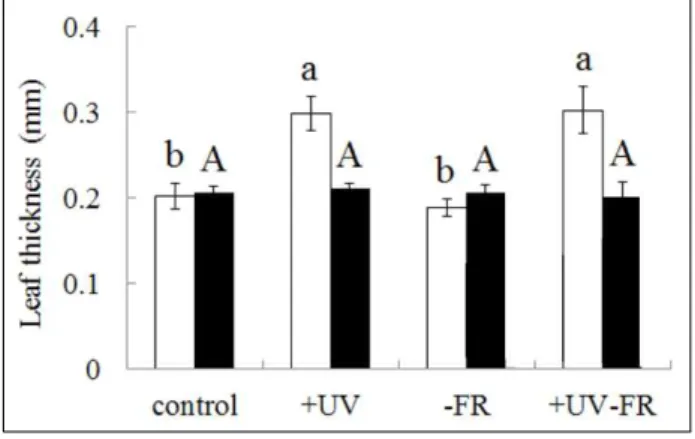
2. Leaf Morphology
Figure 3 shows the leaf mass per area(LMA) and Figure 4 shows the leaf thickness and Figure 5 shows the leaf shape of L. nobilis grown under each treatment. Decreases in LMA and leaf thickness in response to UV-B radiation have been observed previously in the Olea europaea under field and greenhouse condition(Liakouraet al., 1999). However, in this study, for the upper leaves of plants grown in the +UV and +UV-FR treatment groups, there was a significant increase in leaf area and increase in leaf thickness compared with the control group(P<0.05). In contrast, the area and thickness of
Figure 3. Leaf mass per area(LMA) of Laurus nobilis grown under different light conditions for 180 days. Values represent the mean±SE of 6 plants. Bars with different lower-case letters and capital letters are significantly different among upper leaves and lower leaves, respectively(P<0.05). See Materials and Methods for an explanation of the different treatments.
Legend: □ Upper leaves, ■ Lower leaves
Figure 4. Leaf thickness of Laurus nobilisgrown under different light conditions for 180 days. Values represent the mean±SE of 6 plants. Bars with different lower-case and capital letters are significantly different among upper and lower leaves, respectively(P<0.05). See Materials and Methods for an explanation of the different treatments.
Legend: □ Upper leaves, ■ Lower leaves
Figure 5. The shape of upper leaves of Laurus nobilis grown under different light condition for 180 days.
lower leaves were not affected by light conditions. A reduction in FR light also had little effect on leaf area and thickness.
3. Leaf Nutrients
Table 1 shows the nitrogen content, chlorophyll content, and the amount of UV-absorbing compounds in the upper leaves of L. nobilisplants. Plants in the +UV-FR treatment group had lower nitrogen concentrations and higher C/N ratios than other treatment groups, although these differences were not significant.
Chlorophyll a+b and the chlorophyll a/b ratio decreased significantly in plants in the +UV treatment group, mainly as a result of a decrease in the amount of chlorophyll a. In this treatment group, the amount of UV-absorbing compounds per leaf area was also significantly higher than for the control and -FR treatment groups(P<0.05). Similar tendencies have been observed inL. nobilis grown under exposed UV-B radiation in greenhouse condition(Liakoura et al., 1999).
4. Net Photosynthetic Rate
Figure 6 shows the net photosynthetic rate ofL. nobilis
Figure 6. Net photosynthetic rate of Laurus nobilis grown under different light conditions for 180 days. See Materials and Methods for an explanation of the different treatments.
Table 1. Nutrient and chlorophyll(Chl) content of leaves ofLaurus nobilisplants grown under different light conditions
Treatment N in leaves
(mg․g-1) Chl a+b
(μg․g-1) Chl a/b UV-absorbing compounds
(A330․mm-2)
Control 17.1 a 86.4 a 3.7 a 0.062 bc
+UV 16.4 a 45.7 b 3.2 b 0.075 a
-FR 16.8 a 64.2 ab 3.8 a 0.059 c
+UV-FR 14.6 a 55.4 a 3.7 a 0.071 ab
Values represent the mean of 6 plants. Values with different letters within a column are significantly different(P<0.05). See Materials and Methods for an explanation of the different treatments.
grown under each treatment. At light saturation, the maxi- mum photosynthetic rate for plants grown under control conditions was approximately 6 µmol․m-2․s-1. This increased to 8 µmol․m-2․s-1under the +UV treatment, but decreased to 3.8 µmol․m-2․s-1 under the +UV-FR treatment.
Ⅳ. Discussion
Seedlings ofLaurus nobilisthat were grown indoors were affected both morphologically and physiologically by the levels of UV-B irradiation and FR light. UV-B irradiation had large effects on the dry weight, leaf area, leaf thickness, chlorophyll a+b content and a/b ratio, and the amount of UV-absorbing compounds. Experimental studies have shown that different mechanisms are involved in plant protection against UV-B radiation damage(Bornman, 1989; Paoletti, 2005). Since UV- absorbing compounds accumulate in epidermal tissue to prevent UV-B from penetrating the cells, this latter result indicates that these seedlings had adapted to the UV-B irradiation. It was also found that it was only the area and thickness of the upper leaves that were affected by UV-B irradiation, which may indicate either that the upper leaves of seedlings are more exposed to UV-B radiation than the lower leaves, or that the upper young leaves are more sensitive to UV-B radiation than the lower mature leaves, i.e., UV-B tolerance increases with maturity.
Although previous studies have shown that FR light also affects plants(e.g.Petunia hybrid,Glycine maxandNicotiana tabacum)(Fukudaet al., 2002; Kasperbauer, 1988), there was no obvious effect of FR light on leaf morphology or phy- siology in this study. This difference is likely to have been due to the FR cut-off ratio and/or the sensitivity of this species being lower than for previous studies. Many studies have also reported that UV-B irradiation and FR light affect elongation growth when applied on their own(Piriet al., 2011;
Fukuda et al., 2002; Suzuki and Kondo, 2005; Sumitomoet al., 2009; Yoshimura et al., 2002), which also contrasts with our findings. We did, however, find that the simultaneous treatment of plants with UV-B irradiation and FR reduction inhibited elongation growth, which indicated that these light conditions have a complementary effect on elongation growth inL. nobilisseedlings. It has been suggested that the reason for the reduction in height, leaf area and plant dry weight of L. nobilisseedling gruwon in UV-B irradiation and FR reduction is the decreasing photosynthetic activity, such as photo- synthetic electron transport with PS II, and nutrient status of L. nobilisseedlings(Piriet al., 2011). These changes are likely to affect the gross weight of seedlings, and it has also been shown that UV-B irradiation reduces photosynthetic activity in plants(Kondo, 2010).
Several nonsignificant results were also observed in response to supplemental UV-B radiation, including decreased biomass accumulation and photosynthetic rate, and increased UV-B absorbing capacity of the several plants(Manetaset al., 1997;
Grammatikopouloset al., 1998; Liakouraet al., 1999; Stephanou and Manetas, 1997). In this study, the photosynthetic capacity of plants grown under the +UV-FR treatment was lower than for all other treatments, possibly as a result of a change in the amount and/or use of nitrogen in the leaves. Conse- quently, the +UV-FR treatment, which affected plants mor- phologically and physiologically by decreasing the leaf area and weight, and concentration of chlorophyll a, could result in the inhibition of seedling growth. It has previously been reported that UV-B also interacts with other environmental factors to affect the growth of cultivars across years(Murase et al., 1997). Better understanding of the influence of light quality on the growth, leaf shape and physiological activities of Laurus nobilis plants growing indoors is important in plant-light management systems. Therefore, to further investi- gate the complementary effect of UV-B irradiation and FR reduction on the elongation growth ofL. nobilisseedlings, a longer term experiment needs to be conducted that also considers the effect of other environmental factors.
Acknowledgements
This study was supported partly by the Improvement of Research Environment for Young Researchers project of MEXT Japan and the Grant-in-Aid for Scientific Research(type B
24380076, H. Toda) of JSPS Japan.
References
1. Barnes, J. D., L. Balaguer, E. Manrique, S. Elvira and A. W. Davison (1992) A reappraisal of the use of DMSO for the extraction and determination of chlorophylls a and b in lichens and higher plants.
Environmental and Experimental Botany 32(2): 85-100.
2. Bornman, J. F.(1989) Target sites of UV-B radiation on photosynthesis of higher plants. Journal of Photochemistry and Photobiology B: Bioloby 4: 145-158.
3. Fukuda, N., M. Kobayashi, M. Ubukawa and K. Takayanagi(2002) Effects of light quality, intensity and duration from different artificial light sources on the growth of petunia(Petunia×hybrida Vilm.). Journal of the Japanese Society for Horticultural Science 71(4): 509-516.
4. Grammatikopoulos, G., R. Karousou, S. Kokkini and Y. Manetas(1998) Differential effects of enhanced UV-B radiation on reproductive effort in two chemotypes ofMentha spicata under fiend conditions. Australian Journal of Plant Physiology 25: 345-351.
5. Jansen, M. A. K. and J. F. Bornman(2012) UV-V radiation: From generic stressor to specific regulator. Physiologia Plantarum 145: 501-504.
6. Jansen, M. A. K.(2002) Ultraviolet-B radiation effects on plants: Induction of morphogenic responses. Physiologia Plantarum 116: 423-429.
7. Johnson, I. R. and J. H. M. Thornley(1984) A model of instantaneous and daily canopy photosynthesis. Journal of Theoretical Biology 107(4):
531-545.
8. Kang, H., J. Hidema and T. Kumagai(1998) Effects of light environ- ment during culture on UV-lnduced cyclobutyl pyrimidine dimers and their photorepair in rice(Oryza sativa L.). Photochemistry and Photobiology Volume 68(1): 71-77.
9. Kasperbauer, M. J.(1988) Phytochrome involvement in regulation of the photosynthetic apparatus and plant adaptation. Plant Physiology and Biochemistry 26(4): 519-524.
10. Kondo, N.(2010) Effects of enhanced UV-B radiation on plants(in Japanese with English abstract). Teikyo University of Science and Technology Bulletin 6: 1-7.
11. Li, J., T. M. Ou-Lee, R. Raba, R. G. Amundson and R. L. Last(1993) Arabidopsis flavonoid mutants are hypersensitive to UV-B irradiation.
The Plant Cell 5(2): 171-179.
12. Liakoura, V., S. Stavrianakou, G. Liakopoulos, G. Karabouniotis and Y.
Manetas(1999) Effects of UV-B radiation onOlea europaea: Com- parisons between a greenhouse and a field experiment. Tree Physiology 19: 905-908.
13. Manetas, Y., Y. Petropoulou, K. Stamatakis, D. Nikolopoulos, E. Levizou, G. Psaras and G. Karabourniotis(1997) Beneficial effects of enhanced UV-B radiation under field conditions: improvement of needle water relations and survival capacity inPinus pinea L. seedlings during the dry Mediterranean summer. Plant Ecology 128: 100-108.
14. Murase, N., N. Kondo, H. Shimizu, N. Nakajima, T. Izuta and T.
Totsuka(1997) Effects of UV-B irradiation on growth and physiological activities of cucumber(Cucumis sativus L.) first leaves(in Japanese with English abstract). Journal of Japan Society for Atmospheric Environment 32(1) 38-45.
15. Oide, E. and M. Kondo(2001) Viability of four landscaping tree species under interior low light conditions(Papers of the 19th Scientific Research Meeting) (in Japanese with English abstract). Journal of the Japanese Institute of Landscape Architecture 64(5): 537-540.
16. Paoletti, E.(2005) UV-B and mediterranean forest species: Direct effects and ecological consequences. Environmental Pollution 137: 372-379.
17. Piri, E., M. Babaeian, A. Tavassoli and Y. Esmaeilian(2011) Effects of UV irradiation on plants. African Journal of Microbiology Reseach 5(14): 1710-1716.
18. Rozema, J., J. Van de Staaij, L. O. Bjorn and M. M. Caldwell(1997) UV-B as an environmental factor in plant life: Stress and regulation.
Trends in Ecology & Evolution 12: 22-28.
19. Searles, P. S., S. D. Flint and M. M. Caldwell(2001) A meta-analysis of plant field studies simulating stratospheric ozone depletion. Oecologia 127: 1-10.
20. Shinano, T., T. T. Lei, T. Kawamukai, M. T. Inoue, T. Koike, and T.
Tadano(1996) Dimethylsulfoxide method for the extraction of chlorophylls a and b from the leaves of wheat, field bean, dwarf bamboo, and oak.
Photosynthetica 32(3): 409-415.
21. Stephanou, M. and Y. Manetas(1997) The effects of seasons, exposure, enhanced UV-B radiation and water stress on leaf epicuticular and
internal UV-B absorbing capacity ofCistus creticus: A Mediterranean field study. Journal of Experimental Botany 48: 1977-1985.
22. Sumitomo, K., A. Yamagata, K. Shima, M. Kishimoto and T. Hisamatsu (2009) Effect of end-of-day far-red light treatment on flowering and stem elongation in certain cut flowers(in Japanese with English abstract).
Bulltein National Institute Flore Science 9: 1-11.
23. Suzuki, H. and M. Kondo(2005) The effect of UV-B radiation on the growth of plants in interior planting spaces(in Japanese with English abstract). Journal of the Japanese Institute of Landscape Architecture 68(5): 533-536.
24. Yoshimura, T., M. Nishiyama and K. Kanahama(2002) Effects of red or far-red light and red/far-red ratio on the shoot growth and flowering ofMatthiola incane(in Japanese with English abstract). Journal of the Japanese Society for Horticultural Science 71(4): 575-582.
원 고 접 수 일
심 사 일
게 재 확 정 일 3 인 익 명 심 사 필 : : :
2 0 1 2 년 8 월 8 일 2 0 1 2 년 9 월 2 8 일 2 0 1 2 년 1 0 월 1 5 일 2 0 1 2 년 1 0 월 1 6 일
(1차)(2차)