kfas
11
0
0
전체 글
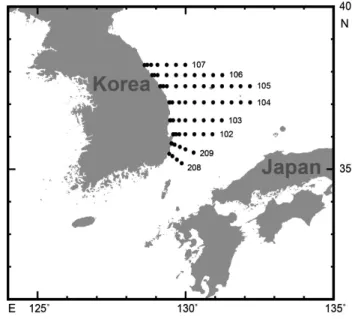
(2) 686. 조민진ㆍ김중진ㆍ양재형ㆍ김창신ㆍ강수경ㆍ차형기. 시작했고, 2015년 이후에는 급격히 감소하면서 2017년에 8만 7천톤을 기록했다(Fig. 1). 채낚기어업은 1970-1980년대까지 전체어획량의 약 80% 수준에 육박했으나, 1990년대 50% 수 준으로 감소했고, 최근에는 약 30% 수준을 기록하고 있다. 반 면, 1990년대 들어 급격히 증가한 대형트롤어업의 어획은 2000 년대 이후 40% 수준을 유지하고 있다. 또한 동해구트롤어업은 2000년대 이후 본격적인 조업을 시작했고, 최근까지 20% 내외 의 어획비율을 나타내고 있다. 1980년대 후반 이후의 살오징어 어획량 증가는 크게 두 가지 원인으로 알려져 있다. Sakurai et al. (2000)은 북서태평양 기 후체제전환(climate regime shift)에 따라 1980년대의 한랭한 해양환경과 비교해서, 1990년대에는 온난한 해양환경이 지속 되었으며, 이로 인해 살오징어의 산란가능(18-23°C) 면적이 확 대되면서 개체군의 크기가 증가하는 결과로 이어졌다고 주장하 였다. 또 다른 원인은 1980년대 이후 급속도로 진행된 연근해 어선의 어획성능 향상 및 어획노력량 증가를 들 수 있다. 오징어 어획의 대표업종인 채낚기어업의 경우 어선의 총톤수, 척당 마 력수 증가, 자동 조획기 도입 등의 어구발달, 집어등의 광력 증 가 등으로 어획성능이 크게 향상되면서 1990년대 어획량 증가 에 영향을 미친 것으로 평가된다(Oh et al., 2018). 한편, 1990 년대 이후 대형트롤과 동해구트롤 등 어획강도가 높은 저인망 류에서도 본격적으로 조업활동을 시작하면서 대상자원에 대한 어획노력량이 크게 높아진 것 또한 어획량 증가 요인 중 하나이 다(Hwang et al., 2012). 그러나 살오징어 어획량은 2003년 이후에는 지속적인 감소추 세를 나타내고 있다. 최근의 어획량 감소는 과도어획이 주요한 원인으로 알려진다. 주어기 동해에서는 채낚기 어선이 집어등 을 밝혀 어군을 모으고, 트롤어선이 그물로 한꺼번에 어획하는 공조조업이 1990년대부터 이어져 오면서 살오징어 자원이 급 격히 감소하였다(Choi et al., 2008). 이와 더불어 2004년부터 북·중국어업협정이 맺어지면서 중국어선은 북한해역에서 주어 기(10월-이듬해 1월)에 대량의 살오징어를 어획하고 있으며, 동 해북측해역을 비롯하여 한일 공동수역(대화퇴)에서까지 대규 모 불법/과도어획을 지속하고 있어 우리 조업어장으로 남하하 는 자원을 감소시켰다(Lee et al., 2017). 또한 어획량이 급감했 던 최근 3년간은 동중국해에서의 겨울철 수온이 산란적정수온 에 비해 낮게 형성되어, 산란량 감소 및 가입량 부진으로 인해 어획량 감소가 더 크게 나타났다(unpublished data from Prof. Sakurai). 살오징어 자원의 지속적 감소와 자원관리 필요성의 대두에 따 라 우리나라는 지난 2007년부터 근해채낚기, 동해구트롤, 대형 트롤, 대형선망 주요 4개 어업에 대해 총허용어획량제도(total allowable catch, TAC)를 실시하고 있다. 또한 2014년부터는 어린 오징어가 성장하는 봄철 4월 한 달 간을 금어기로 지정하 였고, 2016년에는 금어기를 4-5월(단, 연안복합어업과 근해채 낚기어업은 4월 한 달)로 연장하였으며, 금지체장[12 cm ML. (mantle length, ML)]을 신설한 바 있다. 한편, 최근의 급격한 자원감소 추세는 과거 우리 연근해의 명태와 말쥐치 자원처럼 살오징어 자원의 고갈 위험성에 대해 더욱 경각심을 갖게 되는 계기가 되었다. 살오징어는 지난해(2018년) 수산자원회복사업 (해양수산부) 대상어종으로 추가되었으며, 보다 정밀한 자원조 사 및 평가를 바탕으로 자원의 회복·관리 방안 마련을 추진하 고 있다. 본 연구에서는 장기적인 살오징어 자원변동 특성을 비교적 잘 나타내는 것으로 알려진 채낚기어업을 대상으로, 살오징어 어 획수준이 높았던 1990년대(고수준기, 1996-2000년)와 낮았던 2010년대(저수준기, 2013-2017년)의 주어기, 단위노력당 어획 량, 어장분포, 어장환경 등의 어업특성과 체장분포, 산란기, 성 숙체장 등의 생태학적 특성 변화를 비교·분석하였다. 이를 통 해, 살오징어 자원상태를 정확하게 진단하고, 향후 자원관리 방 안 마련 및 자원변동 예측을 위한 필수 정보를 제공하고자 한다.. 재료 및 방법 살오징어 어업특성 분석 우리나라 살오징어의 장기 어획량 변동을 알아보기 위해 1970-2017년의 연도별 월별 어획량 자료(KOSIS, 1970-2017) 를 사용하였다. 단위노력당어획량(catch per unit effort, CPUE) 은 동해안 12개 주요항구에서 오징어채낚기어업을 대상으로 한 국립수산과학원의 어황표본조사자료(채낚기어업 표본어획 량, 조업척수)를 이용하여 1990-2017년의 연도별 척당어획량 (CPUE)를 구하였다.. CPUE=. 어획량 연조업척수. 본 연구에서는 살오징어의 연도별 어획량 및 채낚기 어획량의 장기 변화를 바탕으로 살오징어 어획의 고수준기(1996-2000 년; 5년 평균 전체 어획량, 223천톤)와 저수준기(2013-2017년; 5년 평균 전체 어획량, 137천톤)를 구분하였고(Fig. 1), 두 기 간의 어업특성(주어기, CPUE, 어장분포, 어장환경 등) 변화를 비교·분석하였다. 각 기간의 살오징어 어장분포특성을 비교하 기 위해 국립수산과학원의 어황표본조사자료를 이용하여 채 낚기어업의 연도별 월별 해구별(0.5×0.5°) 어획량자료를 사 용하였고, 평균(5년) 해구별 어장분포와 여름철(6-8월), 가을 철(9-11월), 겨울철(12-2월)로 구분하여 계절적인 어장분포를 비교 분석하였다. 또한 주어장인 동해에서 각 기간의 어장환경 특성을 알아보기 위해 한국해양자료센터(KODC, 1996-2000, 2013-2017)에서 제공하는 격월(2, 4, 6, 8, 10, 12월) 정선해양 관측 수온자료를 활용하였다. 주어장의 영역은 34°15’N-38°N, 128°20’E-131°15’E 내의 정선조사(107-208 Line) 범위를 포함 하였다(Fig. 2). 수심별로는 살오징어의 주 서식수심인 0-50 m.
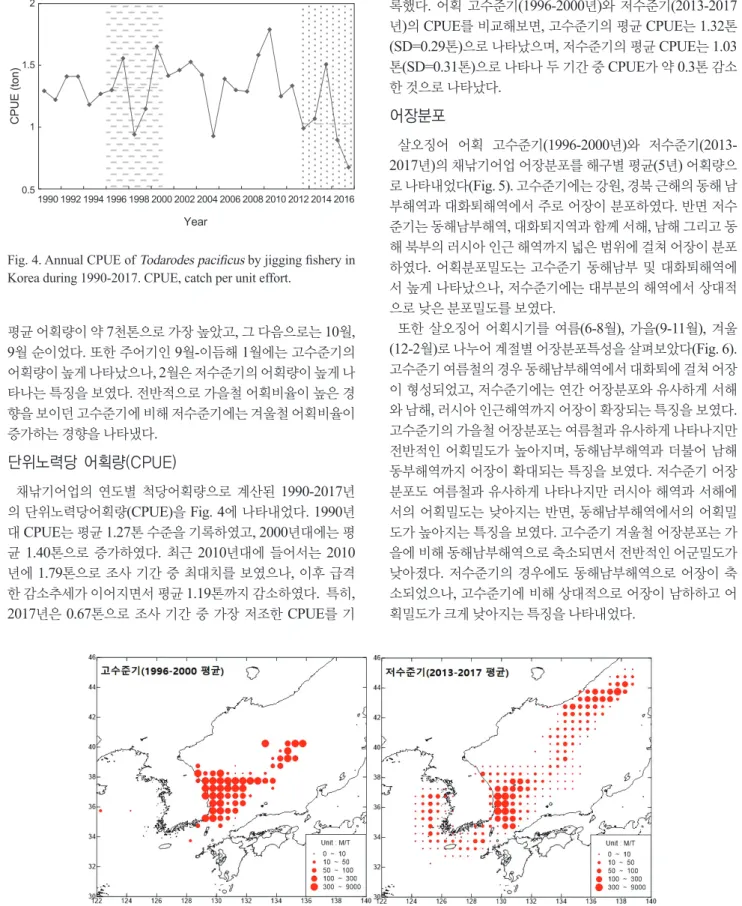
(3) 687. 살오징어 어업·생태학적 변화. 200. 100. 120. 80. 40. Total Average Jigging Average. 200. 100. 120. 80. 40. Jigging fishery Catch (ton). Total Average Jigging Average. Jigging fishery Catch (ton). Total Catch (×1,000 ton). 300. Total Catch (×1,000 ton). 300. 0 0 1970 1975 1980 1985 1990 1995 2000 2005 2010 2015. Time (year). 0 0 1970 1975 1980 1985 1990 1995 2000 2005 2010 2015. Time (year). Fig. 1. Annual catch of Todarodes pacificus in Korea and Jigging fisheries from 1970 to 2017.. Catch (×1,000 ton). 고수준기(1996-2000년)와 저수준기(2013-2017년)에 어획된 살오징어의 어체정밀측정(외투장, 체중량, 생식소 중량, 숙도 80 등) 자료를 바탕으로 1996-2000 각 기간의 생태학적 특징을 분석하였다. 각 기간에 어획된 살오징어 외투장의 빈도분포(length frequen2013-2017 60 cy)를 분석하기 위해 동해 채낚기어업에서 어획된 살오징어를 대상으로 한 월별 어체측정자료(1991-2017년)를 사용하였고, 40 전체 기간(각 5년)에 대한 외투장분포와 여름철(6-8월), 가을철 (9-11월), 겨울철(12-2월)로 구분한 계절별 외투장분포를 각각 20 비교하였다. 고수준기와 저수준기 간 평균외투장 차이는 등분 산 여부를 판단하기 위한 F 검정을 실시한 후, 이분산 가정에 의 0 Feb Mar 유의성 Apr May Jun Jul Aug Oct Nov. 또한 Dec 한 T-testJan 를 수행하여 이내 검증하였다 (P<0.05 )을Sep Time (Month) 어획수준으로 구분된 각 기간의 산란특성치를 비교하기 위해 생식소 숙도 지수(gonad somatic index, GSI)와 50% 성숙체장 을 분석하였다. GSI는 다음과 같은 식으로 계산하였다.. GSI=GW/BW×100 GW, 생식소 중량(g); BW, 체중(g). CPUE (ton). 생식소 중량은 암컷의 경우 난소, 수란관, 난포선의 합으로 구 했으며, 수컷의 경우 정소, 저정낭, 정협낭 및 수정관의 합으로 구하였다. 생식소의 숙도는 측정 시 암컷 개체의 생식소 크기와 색, 난의 투명도, 난립의 분리 상태 등을 육안으로 관찰하여 미 숙(immature), 중숙(maturing), 성숙(mature), 완숙(ripe), 방중 2 (spawning), 방후(spent)의 총 6단계로 구분하였고(Kim et al., 1997), 체장별 성숙 이상의 암컷개체 비율을 로지스틱(logistic) 곡선에 1.5 따라 계산하여 각 연도별 50% 성숙체장을 구하였다. 어 획수준에 따른 각 기간별 성숙체장은 어체측정자료의 연도별. 80 1996-2000. Catch (×1,000 ton). 살오징어 생태학적 특성 분석. Fig. 2. KODC serial oceanographic observation stations in East Sea. KODC, Korea oceanographic data center.. 2013-2017. 60. 40. 20. 0. Jan. Feb Mar. Apr May Jun. Jul. Aug Sep. Oct. Nov Dec. Time (Month). Fig. 3. Monthly catches of Todarodes pacificus by jigging fishery in Korea during 1996-2000 and 2013-2017.. 결측자료수와 로지스틱 곡선의 결정계수(R2) 값을 고려하여 통 계적인 신뢰수준이 높은 해를 선별하였으며, 어획량 고수준기 는 1997년과 1998년, 저수준기는 2014년과 2017년의 자료를 활용하여 각 기간의 50% 성숙체장을 계산하였다.. 결. 과. 2 주조업시기. 어획 고수준기(1996-2000년)와 저수준기(2013-2017년)간 채낚기어업의 주조업시기를 비교하기 위해 각 기간별 월평균 1.5 어획량을 나타내었다(Fig. 3). 고수준기의 경우 월평균어획량 은 10월이 약 2만 3천톤으로 가장 높게 나타났으며 다음으로 9 월, 12월 순서로 높았다. 그에 비해 저수준기의 경우는 12월의. CPUE (ton). 와 채낚기어업의 어구투하 깊이인 50-100 m를 고려하여 0 m, 50 m, 100 m 자료를 이용하였고(Choi et al., 1997), 주어기인 10월과 12월의 각 기간별 평균 수온을 비교하였다.. 1. 1 0.5. 1990 1992 1994 1996 1998 2000 2002 2004 2006 2008 2010 2012 2014 2016.
(4) 688. 조민진ㆍ김중진ㆍ양재형ㆍ김창신ㆍ강수경ㆍ차형기. 록했다. 어획 고수준기(1996-2000년)와 저수준기(2013-2017 년)의 CPUE를 비교해보면, 고수준기의 평균 CPUE는 1.32톤 (SD=0.29톤)으로 나타났으며, 저수준기의 평균 CPUE는 1.03 톤(SD=0.31톤)으로 나타나 두 기간 중 CPUE가 약 0.3톤 감소 한 것으로 나타났다.. 2. CPUE (ton). 1.5. 어장분포. 1. 0.5. 1990 1992 1994 1996 1998 2000 2002 2004 2006 2008 2010 2012 2014 2016. Year. Fig. 4. Annual CPUE of Todarodes pacificus by jigging fishery in Korea during 1990-2017. CPUE, catch per unit effort.. Relative Length Frequency (%). 평균 어획량이 약 7천톤으로 가장 높았고, 그 다음으로는 10월, 9월 순이었다. 또한 주어기인 9월-이듬해 1월에는 고수준기의 어획량이 높게 나타났으나, 2월은 저수준기의 어획량이 높게 나 타나는 특징을 보였다. 전반적으로 가을철 어획비율이 높은 경 향을 보이던 고수준기에 비해 저수준기에는 겨울철 어획비율이 15 증가하는 경향을 나타냈다. 고수준기(1996-2000). 단위노력당 어획량(CPUE) 10. N=4890 저수준기(2013-2017) N=5803. Relative Length Frequency (%). 25. (%). 채낚기어업의 연도별 척당어획량으로 계산된 1990-2017년 의 단위노력당어획량 (CPUE)을 Fig. 4에 나타내었다. 1990년 5 대 CPUE는 평균 1.27톤 수준을 기록하였고, 2000년대에는 평 균 1.40톤으로 증가하였다. 최근 2010년대에 들어서는 2010 0 5 톤으로 10 조사15기간 중 20 최대치를 25 보였으나 30 35 40 년에 1.79 급격 , 이후 length (cm) 감소하였다. 특히, 한 감소추세가 이어지면서 Mantle 평균 1.19 톤까지 2017년은 0.67톤으로 조사 기간 중 가장 저조한 CPUE를 기. 25. Jun-Aug. 20 15. 살오징어 어획 고수준기(1996-2000년)와 저수준기(20132017년)의 채낚기어업 어장분포를 해구별 평균(5년) 어획량으 로 나타내었다(Fig. 5). 고수준기에는 강원, 경북 근해의 동해 남 부해역과 대화퇴해역에서 주로 어장이 분포하였다. 반면 저수 준기는 동해남부해역, 대화퇴지역과 함께 서해, 남해 그리고 동 해 북부의 러시아 인근 해역까지 넓은 범위에 걸쳐 어장이 분포 하였다. 어획분포밀도는 고수준기 동해남부 및 대화퇴해역에 서 높게 나타났으나, 저수준기에는 대부분의 해역에서 상대적 으로 낮은 분포밀도를 보였다. 또한 살오징어 어획시기를 여름(6-8월), 가을(9-11월), 겨울 (12-2월)로 나누어 계절별 어장분포특성을 살펴보았다(Fig. 6). 고수준기 여름철의 경우 동해남부해역에서 대화퇴에 걸쳐 어장 이 형성되었고, 저수준기에는 연간 어장분포와 유사하게 서해 와 남해, 러시아 인근해역까지 어장이 확장되는 특징을 보였다. 고수준기의 가을철 어장분포는 여름철과 유사하게 나타나지만 전반적인 어획밀도가 높아지며, 동해남부해역과 더불어 남해 동부해역까지 어장이 확대되는 특징을 보였다. 저수준기 어장 분포도 여름철과 유사하게 나타나지만 러시아 해역과 서해에 서의 어획밀도는 낮아지는 반면, 동해남부해역에서의 어획밀 도가 높아지는 특징을 보였다. 고수준기 겨울철 어장분포는 가 을에 비해 동해남부해역으로 축소되면서 전반적인 어군밀도가 낮아졌다. 저수준기의 경우에도 동해남부해역으로 어장이 축 소되었으나, 고수준기에 비해 상대적으로 어장이 남하하고 어 획밀도가 크게 낮아지는 특징을 나타내었다.. 고수준기(1996-2000) N=1186 저수준기(2013-2017) N=1989. Fig. 10 5. Catch distribution of Todarodes pacificus by jigging fishery during 1996-2000 (left) and 2013-2017 (right). 5 0. Sep-Nov. 고수준기(1996-2000).
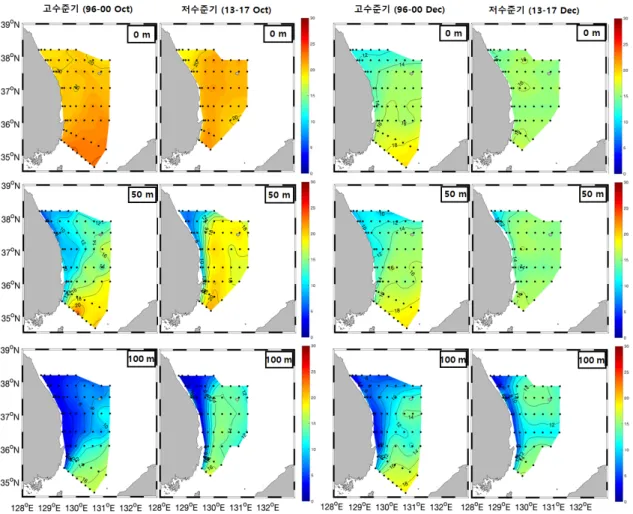
(5) 살오징어 어업·생태학적 변화. 689. Fig. 6. Catch distribution of Todarodes pacificus by jigging fishery in the Korea in summer group (up), autumn group (middle), winter group (bottom) during 1996-2000 (left) and 2013-2017 (right).. 어장환경(수온변동) 어획 고수준기(1996-2000년)와 저수준기(2013-2017년) 동 안의 어장환경변화를 알아보기 위해 주어기(10월-이듬해 1월) 동안 중심어장이 형성되는 동해남부해역에서의 10월과 12월. 의 수심별(0 m, 50 m, 100 m) 수온분포를 나타냈다(Fig. 7). 10 월의 표층수온은 두 기간 모두 20°C 내외의 수온이 분포하면서 큰 차이를 보이지 않았으나 북위 36-38°범위에서는 저수준기 의 수온이 다소 높게 나타났다. 50 m 수심은 고수준기의 경우 8-18°C 수온분포가 동해전역에 걸쳐 넓게 분포하는 것으로 나.
(6) Catc. 20. 0. 690. Jan. Feb Mar. Apr May Jun. 조민진ㆍ김중진ㆍ양재형ㆍ김창신ㆍ강수경ㆍ차형기. Jul. Aug Sep. Oct. Nov Dec. Time (Month). 2. CPUE (ton). 1.5. 1. 0.5. 1990 1992 1994 1996 1998 2000 2002 2004 2006 2008 2010 2012 2014 2016. Year. 15. 고수준기(1996-2000) N=4890 저수준기(2013-2017) N=5803. 10. 5. 0. 5. 10. 15. 20. 25. 30. 35. 40. Mantle length (cm). Fig. 8. Length frequency of Todarodes pacificus by jigging fishery during 1996-2000 and 2013-2017.. 외투장 빈도분포 어획 고수준기(1996-2000년)와 저수준기(2013-2017년)간 동해 채낚기어업에서 어획된 살오징어의 외투장 빈도분포를 비교하였다(Fig. 8). 각 기간의 외투장 범위는 고수준기에 6.4-. (%). 타난 반면, 저수준기에는 18°C 등온선이 북상하면서 8-18°C 범 위의 등온선이 동해 연안으로 밀집되었고 고수준기에 비해 전 반적으로 어장의 수온이 2-4°C 높아졌음을 알 수 있었다. 100 m 수심의 경우에도 50 m 수심과 비슷한 경향으로, 고수준기에 는 4-14°C 범위의 등온선이 외해까지 넓게 분포하였으나, 저수 준기에는 14°C 등온선이 북상하면서 4-14°C 등온선이 연안 쪽 으로 밀집하여 분포하는 특징을 보였다. 전반적으로 100 m 수 온은 고수준기에 비해 저수준기가 2-6°C 높아졌다. 12월의 수 온은 10월에 비해 표층과 50 m 수층에서 계절적으로 낮은 수온 분포를 보인다. 12월의 표층수온은 고수준기에는 12-18°C 범위 의 등온선이 넓게 나타난 것에 비해 저수준기에는 16°C 등온선 이 북상하여 동해남부 전역이 16°C 이상의 수온분포를 나타내 었다. 50 m 수심의 경우에도 10-18°C의 등온선이 넓게 분포한 고수준기에 비해 저수준기에는 14°C 등온선이 북상하면서 동 해 전역이 14°C 이상의 수온분포를 기록하고 있다. 100 m 수심 에서는 12°C 등온선이 북상하면서 전반적으로 북위 37°이북에 서 저수준기의 수온이 높게 나타났다.. Relative Length Frequency (%). Fig. 7. Horizontal distribution of the water temperature at the depth of 0 m, 50 m, 100 m in main fishing season during 1996-2000 and 2013-2017.. 25. Jun-Aug. 고수준기(1996-2000).
(7) 691. Relative Length Frequency (%). 25. Relative Length Frequency (%). 25. Relative Length Frequency (%). 살오징어 어업·생태학적 변화. 25. Jun-Aug. 고수준기(1996-2000) N=1186 저수준기(2013-2017) N=1989. Sep-Nov. 고수준기(1996-2000) N=1873 저수준기(2013-2017) N=2304. Dec-Feb. 고수준기(1996-2000) N=1066 저수준기(2013-2017) N=1007. 20 15. tion)=3.53 cm], 저수준기 22.21cm (SD=3.60cm)로 저수준기 에 약 1.3cm가량 감소한 것으로 나타났다(P<0.05). 이러한 경향은 어획시기를 계절별(여름, 가을, 겨울)로 구분 한 외투장 빈도분포에서도 나타난다(Fig. 9). 여름철(6-8월) 의 외투장 범위는 고수준기 10.6-31.3 cm ML, 저수준기 10.129.3 cm ML이었고, 모드(mode)는 고수준기 19-20 cm, 저수 준기 17-18 cm로 나타났다. 평균외투장은 고수준기에 19.97 cm (SD=2.68 cm), 저수준기에 18.91 cm (SD=2.58 cm)로 약 1.1cm 감소하였다(P<0.05). 가을철(9-11월) 외투장범위는 고 수준기 6.4-30.6 cm ML, 저수준기 13.4-32.5 cm ML이며 모드 (mode)는 고수준기 25-26 cm, 저수준기 24-25cm였다. 평균외 투장은 고수준기에 25.58 cm (SD=2.49 cm), 저수준기에 24.61 cm (SD=2.62 cm)로 약 0.9 cm 감소했다(P<0.05). 한편, 겨울 철(12-2월) 외투장범위는 고수준기 14.1-29.9 cm, 저수준기 14.7-28.5 cm였으며, 모드(mode)는 고수준기 25-26 cm, 저수 준기 23-24 cm로 나타났다. 평균외투장은 고수준기 24.92 cm (SD=2.75 cm), 저수준기 23.71 cm (SD=2.48 cm)로 약 1.2cm 감소했다(P<0.05).. 10 5 0. 20 15 10 5 0. 20 15. 생식소 숙도지수 어획 고수준기(1996-2000년)와 저수준기(2013-2017년)간 산란특성 변화를 확인하기 위해 각 기간의 월평균 생식소 숙 도지수(GSI)를 비교하였다(Fig. 10). 암컷의 경우 고수준기 (n=2,170)에 비해 저수준기(n=2,457)의 GSI가 전반적으로 높 게 나타났고, 반면 수컷의 경우는 고수준기(n=1,701)의 GSI가 모든 관측 기간에 걸쳐 저수준기(n=2,281)에 비해 높게 나타나 는 특징을 보였다. 고수준기의 암컷 GSI는 8월에 7.4로 가장 높 게 나타났고 이후 10월부터는 급격히 감소하여 12월에는 2.2로 낮아졌다. 반면 저수준기의 GSI는 9월에 11.3으로 최고치를 보 였고, 10월 이후 감소하여 12월에는 2.2까지 감소하였다. 1월과 2월은 상대적으로 낮은 GSI를 보이는 시기이나, 7-12월의 경향 과는 다르게 고수준기의 GSI가 상대적으로 높게 나타났다. 수 컷의 GSI의 경우 고수준기는 8-9월이 6.2로 가장 높았고 이후 감소하여 12월에 4.0로 최저치를 나타냈으며, 1월부터 다시 증. 10 5 0. 5. 10. 15. 20. 25. 30. 35. 40. Mantle length (cm). Fig. 9. Length frequency of Todarodes pacificus by jigging fishery in summer, autumn and winter seasons during 1996-2000 and 2013-2017.. 31.3 cm ML, 저수준기에 10.1-32.5 cm ML였으며, 가장 높은 빈도를 보인 외투장 구간(mode)은 고수준기에 25-26 cm, 저수 준기에 22-23 cm로 나타나 저수준기에 좌편향되는 특징을 보 였다. 평균외투장은 고수준기 23.50 cm [SD (standard devia-. 12. Male. Female. GSI. 8. 4. 0. Jul. Aug. Sep. Oct. Nov. Dec. Jan. Feb. Jul. Aug. Sep. Oct. Nov. Dec. Jan. Feb. Month. Fig. 10. Monthly variation of GSI of Todarodes pacificus by jigging fishery during 1996-2000 and 2013-2017( 2017). GSI, gonad somatic index.. : 1996-2000,. : 2013-.
(8) 692. 조민진ㆍ김중진ㆍ양재형ㆍ김창신ㆍ강수경ㆍ차형기. 1.0. 1.0. S(L)=1/(1+exp(8.377-0.380L). S(L)=1/(1+exp(11.537-0.615L). R =0.815. R2=0.953. Maturity ratio. Maturity ratio. 2. 0.5. 0.0. 0. 10. 20. 30. 40. Mantle length (cm). 0.5. 0.0. 0. 10. 20. 30. 40. Mantle length (cm). Fig. 11. Logistic curves based on proportion of mature female of Todarodes pacificus by jigging fishery. Left, 1997, 1998; Right, 2014, 2017.. 가추세를 보였다. 반면 저수준기에는 7월부터 증가하기 시작해 서 10월에 5.0로 가장 높은 값을 나타냈으며, 이후 2월까지 감 소하였다. 고수준기는 암컷과 숫컷의 GSI가 월별로 유사한 증 감패턴을 나타냈으나, 저수준기에는 암컷과 수컷의 GSI 변화 가 불일치하는 특징을 보였다.. 성숙체장 살오징어 어획 고수준기와 저수준기의 산란특성 변화를 알아 보기 위해 두 기간의 성숙체장을 비교하였다(Fig. 11). 최초 성 숙개체가 나타나는 외투장은 고수준기(n=848)에 17.2 cm ML, 저수준기(n=984)는 15.9 cm ML로 저수준기에 보다 작은 크 기에서 최초 성숙이 관찰되었다. 암컷의 50% 성숙체장은 고수 준기의 경우 22.06 cm ML, 저수준기는 18.77 cm ML로 추정 되었으며 고수준기에 비해 저수준기에 약 3.3 cm 감소한 것으 로 나타났다.. 고. 찰. 살오징어 어획 고수준기(1996-2000년)의 경우 10월에 가장 높은 어획량을 나타냈다. 이는 국내 오징어 어획의 주 대상이 가 을산란군이라는 이전연구 결과와 일치하였다(Kim and Kang, 1995). 반면 어획 저수준기(2013-2017년)의 경우 12월의 어획 량이 가장 높게 나타나는 특징을 보였다. 고수준기와 저수준기 의 주어기 어획량 변화를 비교해보면, 9-11월은 77-81%까지 고수준기에 비해 저수준기 어획량이 감소하였으나, 12월과 1월 에는 각각 52%, 72%로 감소폭이 낮아졌고, 2월의 경우는 오히 려 저수준기 어획량이 높게 나타났다. 이는 주로 9-11월에 어획 되는 가을산란군의 어획량은 고수준기에 비해 크게 감소하였 고, 12-2월에 주로 어획되는 겨울산란군의 어획량은 상대적으 로 감소폭이 적거나 증가했음을 알 수 있다. 이는 최근의 국내 어획량의 지속적인 감소는 9-11월의 가을산란군에 대한 어획량 감소가 큰 영향을 미친 것으로 판단된다. 그러나 실제 각 산란군 별 개체군 크기 변동에 대한 연구를 위해서는 산란군별 어미개 체군의 산란자원량, 적정산란환경의 형성 유무, 유생밀도분포,. 가입량 및 회유경로 등에 대한 추가적인 조사 및 분석이 필요할 것으로 생각된다. 또한 국내에서의 산란군별(어획시기별) 어획 변동에 영향을 줄 수 있는 외부적인 요인으로서 남하회유시기 북한수역에서 조업하고 있는 중국어선 및 북한어선의 월별 어 업동향 파악이 반드시 필요하다. 채낚기어업의 어장은 어획 고수준기에는 동해와 대화퇴해역 에 주로 분포하였으나, 저수준기에는 동해와 대화퇴 해역에서 의 어획밀도가 감소한 반면 남해와 서해 그리고 러시아 인근해 역까지 어장이 확장되었다. 이러한 어장변화의 근본적인 원인 은 기존어장에서의 어군밀도 저하로 인한 어획량 감소에 있다. 살오징어를 대상으로 조업하는 채낚기어업은 기존 어장에서의 어획량 감소로 인해 새로운 어장을 찾아 먼 바다로 이동해야 하 는 상황에 직면하게 되었고, 1980-1990년대 어선성능향상(척 당마력수 증가)으로 원거리 어장으로 출어가 용이했기 때문에 (Oh et al., 2018), 서해와 남해, 동해 북부해역까지 어장의 범위 를 넓히게 되었다. 또한 한·러어업협정 체결로 2001년부터 러 시아 EEZ내에서의 살오징어 조업이 가능해졌기 때문에 주로 7 월-9월까지 러시아 수역에서도 조업이 이루어졌다. 한편 서해 안으로의 어장확대는 서해 오징어 자원 증가와도 관련이 있는 것으로 판단된다. 최근 5년간 서해 살오징어 어획량은 큰 폭으 로 증가 한 것으로 보고된다(Unpublished data, from NIFS). Rosa et al. (2011)은 살오징어의 자원상태가 좋지 않았던 1980 년대에는 한랭한 기후조건으로 인해 동중국해에서 큐슈 서부 해역까지 이어지는 산란장분포가 공간적으로 격리되었고, 이 러한 환경에서는 살오징어 어군이 동해보다는 서해 쪽으로 회 유가 발생한다고 주장하였다. 실제 산란장의 면적이 상대적으 로 좁았던 것으로 알려진 1980년대에는 서해의 어획량이 증가 하였고, 1987년에는 동해보다 많은 어획량을 기록한 바 있다 (Choi et al., 1997). 어획량이 감소한 시기에 어장의 분포범위가 넓어지는 것은 어장면적과 살오징어 자원 밀도, 즉 CPUE는 반 비례한다는 이전연구결과(Hwang et al., 2012)와도 일치한다. Kishi et al. (2009)은 살오징어의 어획분포의 변화는 장기적인 기후변화와 관련된 산란장 변화 및 회유경로 변화와 밀접한 연 관이 있다고 보고한 바 있다. 최근 살오징어 어획량 감소 및 본.
(9) 살오징어 어업·생태학적 변화. 연구에서 관찰된 어획 저수준기의 어장범위 확대 또한 장기적 인 기후변화 및 해양환경 변동에 따른 산란장 형성 및 회유경로 변화와 직·간접적인 관련이 있을 것으로 추정된다. 살오징어의 주 어장인 동해는 장기간(1968-2008년)에 걸쳐 표층수온이 상승한 것으로 알려지며, 이는 지난 50년간의 전반 기에 비해 후반기에 더 큰 폭으로 상승한 것으로 밝혀진 바 있 다(Seong et al., 2010). 동해 표층수온 상승의 원인으로는 지 구온난화로 인한 전반적인 기온상승과 동해 상층부의 수온에 영향을 주는 대마난류의 세력이 강해지고 영향을 미치는 범위 가 확장되었기 때문이다(Hong and Cho, 1983). 살오징어의 분 포 및 회유는 서식환경의 수온과 밀접한 관계를 가진다(Cho et al, 2004). 1980년대 후반 북서태평양에서 발생한 기후체제전 환(climate regime shift)에 의한 한랭한 기후조건(1980년대)에 서의 온난한 기후조건으로 변화는 난류성 어종인 살오징어의 산란 및 서식환경에 긍정적인 영향을 미쳐 개체군 크기를 증가 시킨 것으로 평가된다(Kidokoro et al., 2010). 그러나 본 연구 에서는 저수준기(2013-2017년) 동안 동해 주어장에서의 전반 적인 수온상승에도 불구하고 살오징어 어획량은 급격히 감소하 고 있는 것으로 밝혀졌다. 이는 최근의 수온증가는 1990년대와 는 다른 양상으로 살오징어 어획량 변동에 영향을 주고 있는 것 으로 판단된다. Choi et al. (1997)은 주 어획시기에 동해 50 m 수심의 수온전선(thermal front)이 동-서 방향으로 형성되면 전 선이 수온장벽역할을 하면서 전선주변에서 어군이 밀집해 어 획이 좋아지며, 반면 전선이 연안으로 접안하여 남-북 방향으로 형성되면 어군이 북측으로 밀집하여 국내 어황이 좋지 않았다 고 보고했다. 본 연구의 결과에서 어획량 저수준기의 수온분포 는 고수준기에 비해 전반적으로 10°C 등온선이 북쪽으로 이동 하면서 수온이 상승하였고, 수온전선의 방향이 남-북 방향으로 연안에 접안하여 형성된 것을 확인할 수 있었다. 따라서 저수준 기의 어획량 감소는 전반적인 수온상승의 영향에 따라 남-북방 향으로 형성된 수온전선이 수온장벽 역할을 하지 못하면서 어 군이 밀집하지 못하고 북측 등 외양역으로 분산되었기 때문으 로 추정된다. 채낚기어업에서 어획된 살오징어의 평균외투장은 어획량 고 수준기에 비해 저수준기에 약 1.3 cm 감소한 것으로 나타났다. 또한 체장빈도분포(또는 모드)는 고수준기 대비 저수준기에 전 체적으로 좌측으로 치우쳐 나타나는 좌편향(좌경화) 현상이 관 찰되었다. Gong et al. (2006)은 동해 채낚기 오징어를 대상으로 실시한 장기 체장변화 조사를 통해 자원수준이 낮았던 1980년 대 살오징어의 외투장이 자원수준이 높았던 1990년대에 비해 전반적으로 작았다고 보고하였다. 또한 Kasahara (1987)는 일 본의 살오징어 자원수준이 높았던 1960년대와 낮았던 1980년 대 살오징어 측정자료의 비교를 통해, 자원수준이 낮았던 1980 년대의 평균체장 감소와 체장빈도분포의 좌편향 현상을 보고한 바 있다. Yeon and Park (1991)과 Kim et al. (1993)은 우리나라 주요 수산자원인 참조기의 자원 감소시기에 평균체장이 감소. 693. 하고 체장빈도분포는 좌편향 현상이 나타났다고 보고하였다. 자원감소 및 남획의 징후 중 하나로 알려져 있는 어획물의 소 형화가 우리 연근해를 대표하는 주요 어종들에서 관찰되는 점 은 수산자원관리의 관점에서 시사하는 바가 크다고 할 수 있다. 살오징어의 성숙체장은 어획량 고수준기(22 cm)에 비해 저수 준기(19 cm)에 약 3 cm 감소한 것으로 나타났다. 일반적으로 성숙연령은 자원량의 변화에 따라 달라지며, 자원이 감소하면 개체의 영양 상태가 좋아짐으로써 성장률이 높아져서 성숙연령 이 저하된다(Zhang, 2010). Bang (2018)은 우리나라 명태 자원 량 변동에 따른 생태학적 변화 연구에서 자원량이 감소하는 시 기에 성숙체장이 감소하여 성숙이 빨라졌다고 보고하였다. 또 한 참조기의 경우도 자원감소로 인해 개체의 성장이 좋아지고 성숙체장이 소형화된다는 보고가 있었다(Kim et al., 2009). 살 오징어는 명태, 참조기와 같은 어류와는 달리 연체동물에 속하 며, 단년생 어종으로 이들의 생물·생태학적 특징은 상당부분 다 른 것으로 알려져 있다. 그럼에도 불구하고, 자원감소에 따른 생 식학적 반응은 성숙체장 감소라는 유사한 결과로서 나타났다. 앞서 본 연구에서는 어획량 저수준기(2013-2017년)에 동해 주 어장에서의 수온이 고수준기(1993-1997년)에 비해 상승하였 음을 보였다. Richard (1966)는 고수온이 대상자원의 성 성숙 을 가속화 시킬 수 있다고 주장하였고, O’dor and Wells (1987) 는 이에 따른 대사율과 섭이율 사이의 균형 변화가 개체의 성장 을 감소시킬 수 있는 가능성이 있음을 제시한 바 있다. 살오징어 는 저서성 어류인 명태에 비해 해양환경의 변화에 쉽게 노출되 어 있으며, 환경변동에 민감하게 반응하는 것으로 알려져 있는 만큼 고수온 특성이 살오징어의 성장 및 생식특성에 영향을 미 칠 수 있을 것으로 생각된다. 본 연구에서는 살오징어 어획물의 소형화에 대해서는 결과로서 제시하였으나, 성장률에 대한 자 료는 보여주지 못했다. 향후 연구에서는 자원감소와 고수온이 동시에 진행되었을 때 성숙체장 감소와 함께 성장률의 변화를 검증해 볼 필요가 있다. 살오징어는 국내에서는 공조조업 등의 불법어업이 성행하고, 대외적으로는 중국어선의 북한수역 조업이 확대 되는 등 대상 자원에 대한 과도한 어획이 우려되는 상황이며, 장기적인 해양 환경 변화는 살오징어의 자원감소를 가속화하고 있는 것으로 판단된다. Zhang (2010)은 과도한 어획에 따른 남획의 징후로 서 단위노력당 어획량의 감소, 어장면적 축소, 어획물의 소형 화, 성숙연령 감소를 지적하였다. 본 연구결과에서는 어장면적 의 축소를 제외하고는 대부분 남획의 징후에 해당하는 어업/생 태학적 특성의 변화를 확인 할 수 있었다. 어장분포의 경우는 남 획의 징후와는 반대로 어장면적이 확대되는 특징을 나타냈는 데, 이는 과도어획 이외에 장기적인 기후변화 및 해양환경 변동 의 영향이 작용한 것으로 판단된다. 현재 급격한 자원변동으로 어업/생태학적 특성 변화가 진행되고 있는 살오징어 자원은 향 후에도 그 변화를 연속적으로 모니터링하여 자원상태를 신속하 게 진단할 필요가 있다. 또한 자원의 지속 가능한 이용 및 보호.
(10) 694. 조민진ㆍ김중진ㆍ양재형ㆍ김창신ㆍ강수경ㆍ차형기. 를 위해서는 이러한 자원상태 진단을 바탕으로 보다 예방적인 차원에서 자원관리방안을 마련하고 이를 철저하게 이행해야 할 것이다. 아울러, 장기적인 기후변화와 해양환경 변동이 향후 살 오징어 자원에 미치는 영향을 정확하게 예측하기 위한 체계적 인 연구가 반드시 필요하다.. 사. 사. 본 논문은 국립수산과학원 수산과학연구사업인 한국 연근해 어황예측능력 향상 연구(R20199029)의 지원으로 수행되었습 니다.. References Bang. 2018. Changes in ecological characteristics of walleye Pollock Gadus chalcogrammus in accordance with the biomass fluctuation. Ph. D. Dissertation, Pukyoung National University, Busan, Korea. Choi KH, Hwang SD and Kim JI. 1997. Fishing conditions of common squid (Todarodes pacificus Steenstrup) in Korean Waters. I. Spatio-temporal distribution of common squid related to the changes in oceanographic condition. J Kor Fish Soc 30, 513-522. Choi KH, Lee CI, Hwang KS, Kim SW, Park JH and Gong Y. 2008. Distribution and migration of Japanese common squid, Todarodes pacificus, in the southwestern part of the East (Japan) Sea. Fish Res 91, 281-290. http://dx.doi. org/10.1016/j.fisheris.2007.12.009. Gong Y, Jeong HD, Choi KH, Seong KT and Kim SW. 2006. Fluctuations in the abundance of common squid, Todarodes pacificus and environmental conditional in the far east regions during 52 years. J Ecol Environ 29, 1-16. http://doi. org/10.5141/JEFB.2006.29.1.001. Hong CH and Cho KD. 1983. The northern boundary of the Tsushima current and its fluctuation. J Oceanol Soc Kor 18, 1-9. Hwang KS, Kang SK, Oh TY, Choi KH and Lee DW. 2012. Change in the fishing ground and the relationship between the abundance of the common Squid Todarodes pacificus and the distribution of zooplankton in the East Sea. Korean J Fish Aquat Sci 45, 173-179. http://dx.doi.org/10.5657/ KFAS .2012.0173. Kasahara S. 1987. Fishing conditions and stock status of common squid in the Japan Sea for 1986. Suisan Sekai 36, 4248. Kidokoro H, Goto T, Nagasawa T, Nishida H, Akamine T and Sakurai Y. 2010. Impact of climate regime shift on the migration of Japanese common squid (Todarodes pacificus) in the Sea of Japan. ICES J Mar Sci 67, 1314-1322. http://doi. org/10.1093/icesjms/fsq043. Kim JJ, Lee HH, Kim SA and Park C. 2011. Distribution of lar-. vae of the common Squid Todarodes pacificus in the northern East China Sea. Korean J Fish Aquat Sci 44, 267-275. http://dx.doi.org/10.5657/KFAS.2011.0267. Kim JJ, Kim CH, Lee JS and Kim SA. 2014. Seasonal characteristics of Todarodes pacificus paralarval distribution in the northern East China Sea. Korean J Fish Aquat Sci 47, 059071. http://dx. doi.org/10.5657/KFAS.2014.0059. Kim YH and Kang YJ. 1995. Population analysis of the common squid, Todarodes pacificus Steenstrup in Korean Waters-1. Korean J Fish Aquat Sci 28, 163-173. Kim YH, Kang YJ, Choi SH, Park CS and Baek CI. 1997. Population analysis by the reproductive ecological method for the common squid, Todarodes pacificus in Korean waters. 1997. J Kor Fish Soc 30, 523-527. Kim YM, Kim JY, Kim JI, Lee DW, Park CS, Yeon IJ and Park JH. 1993. Assessment of fisheries resources and fishery management in Korean water. Fish Res Rep 12, NIFS, Sinheung Publishing co, Seoul, Korea, 1-181. Kishi MJ, Nakajima K, Fuji M and Hashioka T. 2009. Environmental factors which affect growth of Japanese common squid, Todarodes pacificus, analyzed by a bioenergetics model coupled with a lower trophic ecosystem model. J Mar Sys 78, 278-287. http://doi.org/10.1016/j.jmarsys.2009.02.012 KODC (Korea Oceanographic data center). 1996-2000. Retrieved from http://www.nifs.go.kr/kodc/index.kodc on Sep 16, 2019. KODC (Korea Oceanographic data center). 2013-2017. Retrived from http://www.nifs.go.kr/kodc/index.kodc on Sep 16, 2019. KOSIS (Korean statistical information service). 1970-2017. Retrieved from http://www.kosis.kr on Sep 9, 2019. Lee JS, Ryu JG and Kee HY. 2017. A study on the status of Chinese fishing in the East Sea off North Korea and directions for countermeasures. J Fish Bus Adm 48, 61-74. http:// dx.doi.org/10.12939/FBA.2017.48.3.061. Murata M. 1989. Population assessment, management and fishery forecasting for the Japanese common squid, Todarodes pacificus. In: Marine invertebrate fisheries: their assessment and management. Caddy JR, rd. Jhon Wiley and Sons, New york, NY, U.S.A., 613-616. O’Dor PK, Wells MJ. 1987. Energy and nutrient flow in cephalopods. In: Cephalopod Life Cycles, Vol. 2 (Boyle P, ed). Academic Press, London, U.K., 109-133. Okutani T. 1983. Todarodes pacificus, In: Cephalopod Life Cycles 1, 201-216. Oh TY, Seo YI, Cha HK, Jo HS, An YS and Lee YW. 2018. Change of fishing power index by technological development in the offshore squid jigging fishery. J Korean Soc Fish Ocean Technol 54, 224-230. http://dx.doi.org/10.3796/KSFOT.2018.54.3.224. Richard A. 1966. Action de la temperature sur lévolution genital de Sepia offcinalis L.C.R. Acad Sci Paris 263, 1998-2001..
(11) 살오징어 어업·생태학적 변화. Roper CFE, Sweeny MJ and Nauen CE. 1984. Family Ommastrephidae. In: FAO Species Catalogue. Vol. 3. Cephalopods of the World. An annotated and illustrated catalogue of species of interest to fisheries. FAO Fisheries Synopsis, Rome, Italy, 156-186. Rosa AL, Yamamoto J and Sakurai Y. 2011. Effect of environmental variability on the spawning areas, catch, and recruitment of the Japanese common squid, Todarodes pacificus (Cephalopoda: Ommastrephidae), from the 1970s to the 2000s. ICES J Mar Sci 68, 1114-1121. http://doi. org/10.1093/icesjms/fsr037. Sakurai Y, Kiyofuji H, Saitoh S, Goto T and Hiyama Y. 2000. Change in inferred spawning areas of Todarodes pacificus (Cephalopoda: Ommastrephidae) due to changing environmental conditions. ICES J Mar Sci 57, 24-30. https://doi. org/10.1006/jmsc.2000.0667. Sakurai Y. 2006. How climate change might impact squid populations and ecosystems: a case study of the Japanese common squid, Todarodes pacificus. Globec Rep 24, 33-34. Seong KT, Hwang JD, Han IS, Suh YS and Lee JY. 2010. Characteristics for long-term trends temperature in the Korean waters. J Korean Soc Mar Environ Saf 16, 353-360. Song HJ. 2018. Fluctuations of common squid Todarodes pacificus catches in the northwestern Pacific under changing climate and habitat temperature. Korean J Fish Aquat Sci 51, 338-343. https://doi.org/10.5657/KFAS.2018.0338. Song JY, Lee JS, Kim JJ, Lee HJ, Park MH and Han IS. 2017. Transport process and directly entrainment possibility into the Yellow Sea of Todarodes Pacificus winter cohort. Korean J Fish Aquat Sci 50, 183-194. https://doi.org/10.5657/ KFAS.2018.0338. Yeon IJ and Park CS. 1991. Changes in distribution areas on the stock fluctuation of yellow croaker, Pseudosciaena polyactis Bleeker. Nat’ l Fish Res Inst 45, 85-100. Watanabe T. 1965. Ecological distribution of rhynchoteution larva of common squid, Todarodes pacificus Steenstrup, in the southeastern waters off Japan during the winters, 19591962. Bull Tokai Reg Fish Res Lab 43, 1-12. Zhang CI. 2010. Marine fisheries resource ecology. In: Fisheries science series #1. Pukyoung National University Press, Busan, Korea, 153-187.. 695.
(12)
수치

+3


관련 문서
Expression of human lactoferrin N-lobe in Pichia pastoris and its antibacterial activity.. Su-Jin Won, Jae-Hyung Jo, Seung-Hwan Kim, Hyuk-Jin Kwon,
Purification of Angiotensin I-Converting Enzyme Inhibitory Peptide from Squid Todarodes pacificus Skin.. Jung Kwon Lee, Joong-Kyun Jeon,
Development of Energy Consumption Estimation Model Using Multiple Regression Analysis.. Won-Jae Shin, Yong-Jun Jung 1) , Ye-Jin
Icpyo Kim, Jin-Young Park, Chang Hyeon Kim, Jin-Woo Park, Jae-Pyoung Ahn, Jou-Hyeon Ahn, Ki-Won Kim, Hyo-Jun Ahn, “A room temperature Na/S battery using a β- alumina
Biological Control of Tomato and Red Pepper Powdery Mildew using Paenibacillus polymyxa CW.. Yong-Ki Kim *, Eun-Jung Choi 1 , Sung-Jun Hong, Chang-Ki Shim, Min-Jeong
Properties of Calcium Lactate Prepared from Calcined Littleneck Clam Ruditapes philippinarum Shell Powder.. Gyoon-Woo Lee, In Seong Yoon, Hyun Ji Lee, Jung Suck Lee 1 , Jin-Soo Kim 1
Species Composition of Fish Collected by a Gape Net with Wings in the Coastal Waters of Jindo, Korea.. Jae Mook Jeong, Joon-Taek Yoo, Heeyong Kim 1 , Sun-Kil Lee, Woo-Jin Go
DENTAL MANAGEMENT ASSOCIATED WITH ERUPTION DISORDERS IN A PATIENT WITH EHLERS-DANLOS SYNDROME : A CASE REPORT.. Dallae Jin, Chong-Chul Kim, Sang-Hoon Lee, Jung-Wook Kim, Young-Jae
