흰쥐 심장 발달 과정에서 Protein Kinase C 동종효소 mRNA의 발현 양상
9
0
0
전체 글
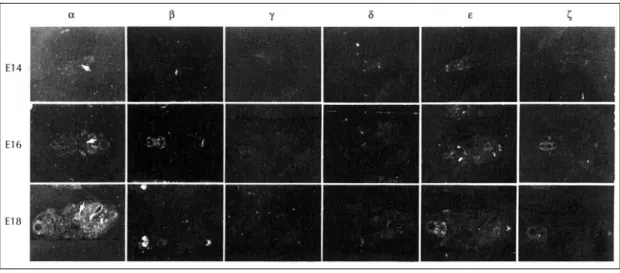
(2) 기도 한다.1-7). 등에 의한 차이도 있으나 가장 문제시되는 것은 심장에. 심장은 발달 단계에 따라 교감신경 자극에 대한 반응 이 다양하게 나타나는데 이러한 다양성은 막 수용체 복. 는 PKC가 비교적 낮은 농도로 존재하기 때문이라고 설명하고 있다.1). 합(membrane receptor complex)이나 세포내 전달. 이러한 점을 감안하여 흰쥐 심장 발달 및 성숙과정에. 과정이 연령에 따라 변화하기 때문이라고 설명하고 있. 서 in situ hybridization 조직화학법으로 일부 PKC 동. 다. α1-adreneric receptor에 대한 심장의 반응은 이. 종효소들(PKC-α , β, γ ,δ ,ε ,ζ )의 mRNA 발현양상을. 들 수용체가 활성화되어 membrane phosphoinositi-. 관찰하여 이미 보고된 다른 방법에 의한 결과와 비교. des의 phospholipase C-의존성 가수분해가 유발하여. 분석하고자 한다.. 두가지 세포내 전달물질(intracellular second mess-. 연구방법. enger)이 유리된다. 그 첫째가 inositol triphosphate (IP3)로서 세포내 칼슘을 움직이고, 두번째가 diacylgycerol(DAG)로서 PKC활성화를 유도한다. 이렇게. 실험동물. 분비된 PKC는 신호 전달의 조절과 세포 발육과 분화. 동물교배는 한 동물장에 몸무게 220~250 gm 사이. 에 관여하게 되고, 어떤 종류의 세포에서는 암세포 발. 의 Sprague-Dawley계 흰쥐 암컷 4마리와 300 gm내. 생을 촉진하기도 한다. 현재까지 심근세포에서의 PKC. 외의 수컷 흰쥐 한마리를 오후 6시부터 다음날 오전 8시. 는 이온 channel, 세포내 이온 농도의 조절, 근 수축력. 까지 합방한 후 생리식염수로 질 도말표본을 만들어 광. 의 조절, 유전자 발현의 조절, 심비대등에 관여하는 것. 학현미경으로 보아 정자가 확인된 날을 임신 제 0 일로. 1-7). 으로 알려져 있다.. 하여 분리 사육하였다. 실험동물로는 출생전 배자기 14. PKC는 크게 classical, novel, atypical subfamily로. 일(E14), 16일(E16), 18일(E18)과 출생 직후(P0), 출. 분류된다.1) Classical(group Ⅰ) PKC subfamily에는. 생 후 7일(P7), 14일(P14), 21일(P21) 및 성숙한(adt). ++. PKC-α,β1,β2, γ 가 있으며, Ca. , phospholipid와. 흰쥐를 사용하였다.. DAG 또는 phorbol ester가 활성화에 필요하다. Novel(group Ⅱ) PKC subfamily에는 PKC-δ,ε,. In situ hybridization 조직화학법을 위한 조직절편 제작. η,θ,μ가 속하며 구조적으로 C2가 없고 PKC-δ,ε. 배자기 14일, 16일, 18일 된 배자는 임신된 어미쥐. 은 활성화하는데 phosphatidyl serine과 DAG가 필요. 를 임신 제 14일, 16일, 18일에 각각 단두한 후 배자를. 하지만 Ca++은 필요하지 않다. Atypical(group Ⅲ). 적출하였으며, 출생직후, 생후 7일, 14일, 21일 및 성숙. PKC subfamily에는 PKC-ζ,ι,λ가 이에 속하며. 한 흰쥐는 단두하여 심장을 적출하여 미리 -30℃로. PKC-ζ는 활성화하는데 phosphatidyl serine이 필요. 냉각된 isopentan에 담궈 얼린 후 조직절편 제작 때까. ++. 하지만 Ca. 은 필요하지 않으며, DAG나 phorbol 1,8-11). ester는 활성화에 필요하지 않다.. 지 -70℃에서 보관하였다. 냉동박절기를 이용해 12 μm 두께의 조직절편을 얻. .. 이러한 PKC 동종효소들은 조직내 분포, 여러 인자의. 어 젤라틴을 입힌 슬라이드에 붙였다. 이 절편들을 고. 조절, 활성화에 필요한 자극제에 대한 반응에 대한. 정액(4% paraformaldehyde in 1×PBS;phosphate. 감수성이 서로 다르게 나타나게 되어 서로 다른 별개. buffered saline)에서 10분간 고정한 후 PBS로 두차. 의 생물학적 기능을 보이게 되며, 이들 PKC 동종효. 례 헹구었다. 정전기에 의한 비특이적 결합을 방지하기. 소의 기능을 이해하는데 심장에서의 PKC의 단백 발. 위해 acetylation 용액(0.25% acetic anhydride in. 현 양상을 확인하는 것이 중요하다. 이러한 PKC 동. 0.1M triethanolamine-HCl, pH 8.0)에 10분간 처리. 종효소들의 발현양상을 보는 방법으로 chromatog-. 한 후 탈수 및 탈지 과정[70% ethanol(1분), 80%. raphy에 의한 분리, immunoblotting, 또는 immun-. ethanol(1분), 95% ethanol(2분), 100% ethanol(1. ocytochemistry등의 방법이 사용되어 왔으나 보고. 분), 100% chloroform(5분), 100% ethanol(1분),. 8,12-17). 자에 따라 약간씩 다른 결과를 보고하고 있다.. 95% ethanol(1분)]을 거쳐 공기 중에서 말린 후 in situ. 이러한 차이는 사용된 실험동물이나 실험동물의 연령. hybridization 조직화학법을 시행하기 전까지 -20℃ 또. 1342. Korean Circulation J 1998;28(8):1341-1349.
(3) l TE 완충액에 용해시켜 liquid scintilation counter로 방. 는 -70℃에 보관하였다.. 사능활성도(radioactivity)를 측정하였다.. Oligonucleotide probe 제작 PKC isoform(α, β, γ, δ, ε 및 ζ)의 염기서열. In situ hybridization 조직화학법. 에 상보적인 염기서열의 oligonucleotide를 DNA 합성. 표지된 probe가 함유된 hybridization 용액(50% fo-. 기(Applied Biosystems DNA synthesizer)로 제작하. rmaldehyde, 10% dextran sulfate, 0.7% ficoll, 0.7%. 고, 암모니아를 제거하여 말린 후 50 μl 물에 용해시. polyvinyl pyrrolidone, 0.7% bovine serum albumin,. 키고 같은 양의 formamide를 넣었다. 잘 혼합하여. 0.15 mg/ml yeast tRNA, 0.33 mg/ml denatured. 55℃에서 5분간 가열하여 2차 구조 형성을 억제하고. salmon sperm DNA, 20 μM dithiothreitol)을 각 슬. tracking dyes(0.05% xylene cyanol FF and 0.05%. 라이드 당 60 μl씩(1×106 cpm) 점적하고 덮개유리. bromophenol blue)를 넣어 준 다음 1.5 mm 두께의. 로 덮은 후 37~42℃에서 16~24시간 동안 반응(hy-. 15×20cm 10% polyacrylamide/urea gel (60ml. bridization)시켰다. 반응이 끝나면 1×SSC로 30초간. solut-ion;25.2 g urea, 6 ml 10×TBE, 20 ml 30%. 행군 다음 42℃에서 2×SSC/50% formamide에 15. acry-lamide mix, 200 μll 10% ammo-nium. 분씩 네번 수세한 후 상온에서 1×SSC로 30분씩 두번. persulfate, and 30 μl N, N, N’, N’-tetramethyl-. 수세하고 증류수와 70% ethanol로 한번씩 헹군 다음 공. ethylene diamine)에 400 볼트의 전압으로 전기영. 기 중에서 말렸다. 이 슬라이드들을 β-max hype-. 동한 후 fluore-scent thinlayer chromatographic. rfilm(Amersham)에 2주간 노출시킨 후 D-19(Ko-. plate 위에 gel을 얹고 암실에서 long-wavelength. dak) 및 Kodak Rapid Fixer로 현상하여 결과를 분석하. ultraviolet lamp로 관찰하였다. 뚜렷한 단일 밴드를. 였다. 필름자가방사능법(film autoradiography)이 끝난. 확인하여 칼로 잘라내어 0.7% low meling agarose gel. 슬라이드들은 nuclear emulsion 용액에 담근 후 8주간. 에 60볼트로 전기영동하였다. 다시 oligonucleotide. 노출한 후 현상하여 슬라이드 자가방사능법(slide auto-. band를 분리한 다음 tube에 넣고 같은 양의 TE 완충액. adiography) 결과를 현미경으로 관찰하였다.. (TrisEDTA, pH 8.0)을 넣어 agarose를 65℃에서 녹. 결. 인 후 phenol과 chloroform/isoamyl alcohol로 추출하. 과. 고 에탄올로 침전시켜 말린 후 물에 녹여 흡광기로 농도. 필름 자가방사능법(film autoradiography)으로 관찰한. 를 측정하였다.. PKC 동종효소(α, β, γ, δ, ε, ζ) mRNA 발현양상 Oligonucleotide probe labeling. (Fig. 1, 2). 정제된 oligonucleotide를 terminal deoxynucleotidyl transferase (TdT)와[35S]dATP (New Eng-. PKC α. land Nuclear, 1300 Ci/mmol 이상)를 이용하여 3’ 끝. 발현을 관찰한 E14에서 성숙한 흰쥐에 이르기 까지. 표지법(3’tailing)으로 방사선 표지하였다. 반응 혼합. 심장에서 발현하는 것을 확인할 수 있었다. E14에서. 물은 TdT buffer(100mM potassium cacodylate, pH. 낮은 발현이 관찰되기 시작하여 P7까지 지속적으로 발. 7.2, 10mM CoCl2, 0.2 mM dithiothreitol), 0.1 μM. 현이 증가하여 비교적 높은 발현을 보였으며, 그 이후. 35. oligonucleotide 및 1 μM [ S]dATP와 TdT 100 unit. 약간 감소하여 성숙한 흰쥐 심장에서 보이는 정도의 발. 를 첨가하고 2차 증류수로 양을 50 μl로 맞추어 37℃에. 현을 보였다.. 서 5분간 반응시킨 후 400 μl의 TE buffer(10 mM Tris-HCl, pH 7.5/1 mM EDTA)로 반응을 종식하였다.. PKC β. 표지된 oligonucleotide는 phenol /chloro-form-. 전신 조직절편에서 mRNA 발현을 관찰한 배자기에. /isoamyl alcohol(50:49:1)과 chloroform/isoamyl. 서 신경계에는 높은 발현을 보이는 데 비해 심장에서는. alcohol(24:1)로 추출하여 에탄올로 침전시킨 후 50 μ. 전혀 발현하지 않았으며, 출생후에도 P7까지는 배자기 1343.
(4) 에서와 마찬가지로 유의할만한 발현양상을 보이지 않. PKC γ. 았으나, P14에 아주 낮은 발현이 관찰되었으며, P21에. PKC γ는 전신 조직절편에서 mRNA 발현을 관찰. 는 비교적 높은 발현이 관찰되었으며, 성숙한 흰쥐 심. 한 배자기에서 뇌에서 발현하는 것은 확인할 수 있었으. 장에서는 약간 감소하는 것을 확인할 수 있었다.. 나 심장에서는 전혀 발현하지 않았으며, 출생후에도 심. Fig. 1. Localization of PKC isoforms (α, β, γ, δ, ε and ζ) mRNA in the prenatal (E14, E16, E18) rat hearts with in situ hybridization histochemistry. Arrow indicates heart.. Fig. 2. Localization of PKC isoforms (α, β, γ, δ, ε and ζ) mRNA in the postnatal (P0, P7, P14, P21, adult) rat heart with in situ hybridization histochemistry.. 1344. Korean Circulation J 1998;28(8):1341-1349.
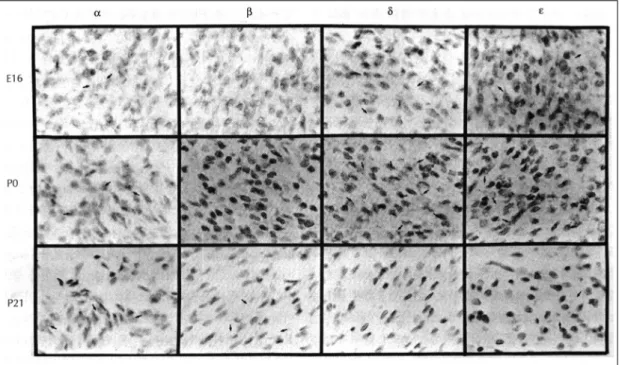
(5) Fig. 3. Cellular localization of PKC isoforms (α, β, δ, ε) mRNA in the developing (E16, P0 and P21) rat hearts with in situ hybridization histochemistry. Arrows indicates the silver grains on the cardiomyocytes.. 장에서의 발현을 관찰할 수 없었다.. PKC δ 관찰을 시작한 E14에서부터 발현을 보였으며, 그 이 후 점차 발현이 증가하여 P0에는 높은 발현을 보인후 급격히 발현이 감소하여, P14에서부터는 발현을 관찰 할 수 없었다.. PKC ε 관찰을 시작한 E14에서부터 발현을 보였으며, 그 이. Table 1. The mRNA expression of PKC isoforms in the developing and adult heart E14. α. β. γ. δ. ε. ζ. +. -. -. +. +. -. E16. +. -. -. +. +. E18. ++. -. -. ++. ++. -. P0. ++. -. -. +++. +++. -. P7. +++. -. -. +. ++. -. P14. ++. +/-. -. -. +. -. P21. ++. ++. -. -. +. -. Adt. ++. +. -. -. +/. -. 후 점차 발현이 증가하여 P0에는 높은 발현을 보인후 점차 발현의 감소를 보여, 성숙한 흰쥐 심장에서는 아 주 낮은 발현만을 확인할 수 있었다.. 슬라이드 자가방사능법(slide autoradiography)으로 관 찰한 PKC 동종효소(α, β, δ, ε) mRNA 발현양상Fig. 3) (Table 1). PKC ζ. 필름 자가방사능법으로 심장에서 발현을 확인할. PKC ζ는 PKC γ와 마찬가지로 전신 조직절편에. 수 있었던 PKC 동종효소(α, β, δ, ε) mRNA의. 서 mRNA 발현을 관찰한 배자기에서 뇌에서 발현하. 세포 수준에서의 발현양상을 E16, P0, P21에 슬라. 는 것은 확인할 수 있었으나 심장에서는 전혀 발현하. 이드 자가방사능법으로 관찰한 결과, 필름 자가방사. 지 않았으며, 출생후에도 심장에서의 발현을 관찰할. 능법에서 보였던 결과와 일치하는 것을 확인할 수. 수 없었다.. 있었다.. 1345.
(6) PKC α는 E16에 약한 발현을 보였으나 P0에는 비. 세포내 Ca++ 농도를 증가시킨다. 세포반응 후기에는. 교적 높게 발현하였으며, P21에서도 P0와 유사한 정도. phosphatidyl choline이 phospholipase D에 의해 가. 의 발현을 보였다. PKC β는 E16과 P0에는 유의할. 수분해되어 DAG가 형성되고, phospholipase A2에 의. 만한 발현을 보이지 않았으나 P21에 비교적 높게 발현. 해 free cis unsaturated fatty acids와 lysoph-. 하였다. PKC δ는 E16에 약한 발현을 보였으나 P0에. osphatidyl choline이 형성되어 DAG에 의존하는 PKC. 는 높게 발현하였으며, P21에서는 유의할 만한 발현을. 의 활성화를 강화하며, DAG은 PKC의 Ca++에 대한. 관찰할 수 없었다. PKC ε는 E16에 약한 발현을 보였. 친화력를 증가시킨다. 즉 세포내에서 PKC의 활성화는. 으나 P0에는 높게 발현하였으며, P21에서는 아주 낮은. 처음에는 세포내 Ca++ 농도의 증가와 inositol pho-. 발현만을 관찰할 수 있었다.. spholipid의 가수분해로 생성된 DAG에 의해 일어나고, 그후 PKC의 활성화는 세포내 Ca++ 농도가 증가되지. 고. 안. 않더라도 DAG과 free cis unsaturated fatty acids에 의해 지속적으로 유지되게 된다.22) 심장에서 활성화된. Protein kinase는 Takai 등18)이 뇌에서 C2+와 인. PKC는 cardiac action potential plateau와 repolar-. 지질에 의존하는 protein kinase를 처음 발표하면서. ization phase에 참여하는 칼슘 및 potassium 채널을. 알려지기 시작했고, Kuo 등19)은 심장을 비롯한 여러. 조절하고,1) 근섬유기질단백(myofilament protein su-. 조직에서 이 kinase의 활성을 증명하였고, 이후 Ka-. bstrate)을 인산화하여 칼슘에 대한 민감도를 증가시. toh 등20)이 심장에서의 protein kinase의 역할 및 기. 켜 심장의 수축력에 영향을 미친다.23) 또한, PKC는 세포증식에 관여하는 c-myc, c-fos,. 능을 자세히 보고하였다. PKC는 다양한 세포에서 신경전달 및 세포외유출. c-jun등의 protooncogene의 발현을 유발하고 배자기. (exocytosis), 세포 이동, 근육 수축, 세포의 성장등을. 유전자들(skeletal α-actin, β-myosin heavy chain,. 포함한 생리적 작용에 중요한 역할을 담당하는 단백 인. atrial natriuretic factor)과 수축단백 유전자들(my-. 산화 효소(protein phosphorylating enzyme)로서 심. osin light-chain 2, cardiac α-actin)의 발현을 증가. 장에서는 autonomic agonist 들에 반응하여 calcium. 시킨다.1,5,6,24) 따라서 심근세포에서 α1 수용체의 자극. 과 potassium등의 ion channel을 조절하고, 심장의 수. 은 이들 유전자의 발현에 따른 단백질합성을 증가시킴. 축력 및 심근비대에 관여하는 것으로 알려져 있다.8,9). 으로써 심근 비대를 유발한다.25) 본 연구에서 태생기부. 그러나 심장은 막수용체복합의 구성요소와 세포내 신. 터 성인기까지 심장 발달 단계에 따라 여러 PKC동종효. 호전달 과정이 연령에 따라 변화하기 때문에 발달단계. 소가 시기에 따라 다르게 발현되는 양상은 심근 비대를. 에 따라서 autonomic agonists들에 대한 반응이 다양. 일으키는 유전자들과 연관이 있으리라 생각된다.. 하게 나타나고, 10여종으로 이루어져있는 PKC 동종효. 본 연구에서 PKC-α는 배자기 14일부터 성숙기에. 소들이 신경호르몬등의 autonomic agonist들에의해 서. 이르기까지 전반적인 발현양상을 보였는데 이는 Im-. 로 다른 반응을 보임으로써 이들 효소들이 발달단계에. munoblotting을 이용한 여러 연구8,12-15)에서 PKC-. 따른 심장기능에 있어서 개개의 특정한 역할을 수행할. α가 흰쥐의 신생아기 및 성숙 심장에서, 또한 배양된. 것으로 생각되어지고 있다.21). 신생아기 심근세포와 성숙 심근세포에서 존재한다는. PKC의 활성화는 호르몬, 신경전달물질, 성장인자등. 보고와 일치하는 소견이며 Kohout 와 Rogers16)가. 에 의해 세포막 인지질이 가수분해되거나 세포막 칼슘. Nothern blotting을 이용하여 신생아기 심근세포와 성. 채널이 열려 세포내 Ca++ 농도가 증가됨으로써 이루. 숙 심근세포에서 PKC-α mRNA가 존재한다는 보고. 어진다. 이들 agonist들에 의한 세포반응 초기에는 세. 와도 일치하는 소견을 보였다.. ++. 농도가 증가되거나,. PKC-β는 다소 논란이 있는데 Wetsel 등12)은 성. phospholipase C(PLC)가 활성화되어 phosphatidyl. 숙 적출 심장에서 immunoblotting과 면역조직화학법. inositol 4, 5-bisphosphate(PIP2)로부터 inositol 1, 4,. 등을 이용하여 PKC-β의 존재를 확인하였고 Rybin. 5-trisphosphate(IP3)와 DAG를 생성하는데 IP 3는. 등14)은 immunoblotting을 이용하여 성숙 심장뿐 아니. 포막 칼슘채널이 열려 세포내 Ca. 1346. Korean Circulation J 1998;28(8):1341-1349.
(7) 라 신생아기 심장에서 PKC-β의 존재를 확인할 수 있 13). 보여 성숙기에는 그 발현이 미약하였다. PKC-ε은 전. 었으나 immunoblotting을 이용한 Bogoyevitch 등 은. 반적으로 발달단계에 따라 유의한 발현을 보였으나 신. 성숙 적출 심장에서, Nothern blotting을 이용한 Ko-. 생아기에 가장 풍부하게 존재하였다. Angela 등8)도. 16). hout와 Rogers. 는 배양된 신생아기 심근세포와 배. PKC-α, β, δ, ε, ζ등이 신생아기 심장에서 발현. 양된 성숙 심근세포에서 각각 PKC-β와 PKC-β. 되고 PKC-α, δ, ζ는 빠르게, ε은 다른 동종효소들. mRNA의 존재를 확인할 수 없었다고 한다. 본 실험. 에 비해 천천히 감소함을 보고하였다.. 에서는 배자기 및 출생후 14일까지 즉, 신생아기에. 한편, 심장은 발달 단계에 따라서 autonomic ag-. 는 PKC-β mRNA의 발현을 관찰할 수 없었고, 출. onist에 대한 반응이 다양하게 나타나는데 이러한 차이. 생후 21일과 성숙기에만 발현을 확인할 수 있었다.. 는 막 수용체 복합(membrane receptor complex)의. 보고간에 서로 다른 차이가 있으나 PKC-α mRNA. 구성요소와 세포내 신호전달 과정이 연령에 따라 변화. 가 출생후 21일과 성숙기에만 발현한 본 실험의 결. 하기 때문이다.14,27) 또한, PKC 동종효소들이 신경호르. 과는 PKC-β mRNA가 다른 동종효소들과는 달리. 몬에 대해 서로 다른 반응을 보임으로써 심장의 기능에. 성숙기에서 더욱 중요한 역할을 담당할 것임을 암시. 있어서 각각 서로 다른 특정한 역할을 수행할 것으로 생. 하고 있다.. 각되어지고 있다. 이러한 특성으로 인해 그간 심장에서. PKC-γ는 신경조직에만 국한하여 분포한다고 알려 22). 져 왔는데,. 본 실험에서도 심장의 발달과정중 어떤. 시기에도 PKC-γ mRNA발현을 관찰할 수 없었다.. 규명하기 위한 필수적인 선행조건이 되어왔다.13) Insitu hybridization은 Nothern blotting과 함께 cDNA. PKC-δ는 immunoblotting을 이용한 Bogoyev13,26). itch 등. PKC 동종효소의 분포를 알아보는 것은 그들의 역할을. probe를 이용하여 PKC 동종효소의 조직내 분포를 알아. 은 성숙 적출 심장과 배양 심근세포에서. 보는 방법으로 특정한 PKC 동종효소의 mRNA가 조직. 는 관찰할 수 없었으나, 배양된 신생아기 심근세포에. 내에 상대적으로 많이 분포할수록 단백이 많이 존재할. 서는 PKC-δ가 존재함으로써 배자기 및 출생 직후. 것이라는 명백한 가정에 근거를 두고 있다.. 에는 PKC-δ mRNA가 유의한 발현을 보이다가 출. 본 실험에서도 이들과 아주 유사한 결과를 보였는데,. 생후 7일부터는 급격히 감소함을 보인 본 실험의 결. PKC-δ와 ε은 출생후 7일부터 발현이 감소하기 시. 과와 유사함을 보여준다. 그러나 이와는 다르게. 작하였으며 특히 PKC-δ는 급격히 감소하였고,또한. Wetsel 등12)은 immunoblotting과 면역조직화학법. PKC-ε도 급격히 감소하였으며 PKC-α는 출생후. 을 이용하여 성숙 적출심장에서 PKC-δ의 발현을. 14일부터 감소하기 시작하였으나 전반적으로 성인기에. 보고한 바 있다.. 이르기까지 유의한 발현을 관찰하였다. PKC-α, δ,. PKC-ε은 배자기 및 출생후부터 성숙한 흰쥐 심장. ε는 발달단계에 따라 그 발현이 감소함을 보임으로써. 에 이르기까지 전반적으로 유의할만한 발현을 보였는. 이들이 빠르게 성장하는 시기에 중요한 역할을 담당하. 데 출생후 7일부터 점차 발현강도가 감소하는 양상을. 리라 생각되어지며 또한 심장이 발달 단계에 따라서. 보였다. 다른 보고들8,13-15)에서도 PKC-ε가 신생아. autonomic agonist에 대한 반응이 다양하게 나타나는. 기 및 성숙한 심근세포에서 폭넓게 존재한다는데 의견. 데 PKC 동종효소의 변화가 관련되어 있을 가능성을. 이 일치하고 있다.. 시사한다.. PKC-ζ는 다른 여러 보고들과 달리 심장에서는 어. 각 연구마다 약간의 서로 다른 차이를 보이는데 이는. 14). PKC 동종효소가 다른 여러 조직(특히 신경조직동)에. 는 PKC 동종효소의 발현이 발달단계에 따라 변화가. 비해 심장에 분포하는 농도가 낮고, 실험동물의 strain. 있는지 알아보기 위해 태아기 14일과 출생후 2일에서. 의 차이, 서로 다른 실험방법 및 기술적 차이가 존재하. 15일까지의 신생아기와 성숙기 흰쥐 심장의 단백 추출. 기 때문이라 생각되어진다. 많은 연구들이 심장에서. 물로부터 immunoblotting을 이용하여 조사하였는데. PKC를 활성화시키는 phorbol ester등을 이용하여. PKC-α와 δ는 태아기와 신생아기에는 유의한 발현. PKC의 역할을 규명하기 위해 시행되어 왔으나, 다양. 을 보인 반면, 생후 2주째부터 급격히 감소하는 경향을. 한 PKC 동종효소에 대한 기능에 대하여 아직 명확하. 떠한 발현도 관찰할 수 없었다. Rybin과 Steinberg. 1347.
(8) 게 규명되지 않은 실정이다. 앞으로 그들 동종효소들의. 중심 단어:Protein kinase C・심장・발달・in situ hy-. 정확한 분포와 더불어 심장내에서 그들 개개의 역할을. bridization.. 규명하는 것이 매우 중요한 일이라 생각된다.. 요. 약. ■ 감사문 본 논문은 고려대학교 의과대학 생명과학연구소의 연구비 보 조로 이루어졌습니다.. REFERENCES. 서 론: PKC는 다양한 세포에서 성장에 관여하며, 심장에서. 1) Puceat M, Brown JH. Protein kinase C in the heart. In.. 각각의 동종효소들이 agonist에 대해 각기 다른 반응을 보이며, 발달과정에서 agonist에 대한 반응의 변화가. 2). 알려져 있다. 이번 연구에서는 심장 발달과정에서 PKC isozymes(α, β, γ, δ, ε, ζ) mRNA의 발현양상 을 in situ hybridization 조직화학법으로 관찰하여 각. 3). 각의 PKC 동종효소들의 심장 발달과정에서의 역할을 예측하고자 하였다.. 재료 및 방법: 심장 발달 과정에서 PKC isoforms(α, β, γ, δ, ε, ζ) mRNA의 발현 양상을 in situ hybridization histo-chemistry로 관찰하였다. 배자기 14일(E14),. 4) 5). E16, E18은 흰쥐 배자 시상절편은 사용하였으며, 출생 직후(P0), P14, P21,성숙한 흰쥐는 심장 절편을 사용하 였다.. 6). 결 과: PKC α는 배자기 14일 부터 심장에서 발현을 관찰할 수 있었으며, 그 이후 점차 증가하여 출생후 7일에 가장. 7). 높게 발현한 후 약간 감소하였다. PKC β는 P14부터 발 현이 관찰되었으며, P21에 비교적 높게 발현한 후 성숙 한 흰쥐에서 약간 감소하는 양상을 보였다. PKC δ는. 8). E14에 발현을 보이기 시작하여, P0에 가장 높게 발현한 후 급격히 감소하여 P14부터는 발현을 관찰할 수 없었 다. PKC ε은 E14에 발현하기 시작하여 P0에 가장 높. 9). 게 발현하였으며 그 이후 발현이 서서히 감소하여 성숙. 10). 한 흰쥐 심장에서는 발현을 관찰할 수 없었다. PKC γ와. 11). ζ는 심장 발달과정에서 전혀 발현을 보이지 않았다.. 결 론:. 12). 이상의 결과에서 PKC 동종효소들이 심장 발달과정에 서 각기 다른 역할을 할 것을 예상할 수 있었으며, 심장. 13). 이 왕성하게 성장하는 출생 전후에 높은 발현을 보이는 PKC α, δ, ε은 심장 성장에 중요한 역할을 할 것을 예상할 수 있으며, 발달과정 후기에 발현을 보이는 PKC β는 심장 성숙에 중요한 역할을 할 것으로 예상된다. 1348. 14). Kuo JE, editors. Protein Kinase C. Oxford, UK: Oxford University Press;1994. p.249-68. Allen BG, Katz S. Isolation and characterization of the calcium- and phospholipid-dependent protein kinase (protein kinase C) subtypes from bovine heart. Biochemistry 1991;30:4334-43. Capogrossi MC, Kaku T, Filburn CR, Pelto DJ, Hansford RG, Spurgeon HA, et al. Phorbol ester and dioctanoyl glycerol stimulate membrane association of protein kinase C and have a negative inotrophic effect mediated by changes in cytosolic Ca2+ in adult rat cardiac myocytes. Circ Res 1990;66:1143-55. Puceat M, Hilal-Dandan R, Bruton LL, Brown JH. Neurohormonal regulation of PKC isozymes in isolated cardiac cells. Biophys J 1993;64:A76. Sei CA, Irons CE, Sprenkle AB, McDonough PM, Brown JH, Glembotski CC. The α-adrenergic stimua-ion of atrial natriuretic factor expression in cardiac myocytes requires calcium influx, protein kinase C and calmodulin regulated pathways. J Biol Chem 1991;266:15910-6. Dinnmon PM, Iwaki K, Henderson SA, Sen A, Chien KR. Phorbol ester induce immediate early-genes and activate cardiac gene transcription in neonatal rat myocardial cells. J Moles Cell Cardiol 1990;22:901-10. Otani H, Mitsuyoshi H, Xun-Tsing Z, Omori K, Inagaki C. Different patterns of protein kinase C redistribution mediated by α1-adrenoreceptor stimulation and phobol ester in rat isolated ventricular papillary muscle. Br J Pharmacol 1992;107:22-6. Angela C, Bogoyevitch MA, Fuller SJ, Lazou A, Parker PJ, Sugden PH. Expression of protein kinase C isoforms during cardiac development. Am J Physiol 1995;269: H1087-97. Azzi A, Boscoboinik D, Hensey C. The protein kinase C family. Eur J Biochem 1992;208:547-57. Dekker LV, Parker PJ. Protein kinase C- a question of specificity Trends Biochem Sci 1994;19:73-7. Nishizuka Y. Studies and perspectives of protein kinase C. Science 1986;233:305-11. Wetsel WC, Khan WA, Merchenbthaler I, Rivera H, Halpern AE, Phung HM, et al. Tissue and cellular distribution of the extended family of protein kinase C isoenzymes J Cell Biol 1992;117:121-33. Bogoyevitch MA, Paker PJ, Sugden PH. Characterization of protein kinase C isotype expression in adult rat heart. Circ Res 1993;72:757-67. Rybin VO, Steinberg SF. Protein kinase C isoform expression and regulation in the developing rat heart. Circ Res 1994;74:299-309.. Korean Circulation J 1998;28(8):1341-1349.
(9) 15) Puceat M, Hildal-Daldon R, Strulovici B, Bruton LL,. 21) Ono Y, Fujii T, Ogita K, Kikawa U, Igarashi K, Nis-. Brown JH. Differental regulation of protein kinase C isoforms in isolated neonatal and adult rat cardiomyocytes. J Biol Chem 1994;269:16938-44. Kohout TA, Rogers TB. Use of a PCR-based method to characterize protein kinase C isoform expression in cardiac cells. Am J Physiol 1993;264:C1350-59. Dasatnik MH, Buraggi G, Rosen DM. Localization of protein kinase C isoenzymes in cardiac myocytes. Exp Cell Res 1990;210:287-97. Takai Y, Kishimoto A, Iwasa Y, Kawahara Y, Mori T, Nishizuka Y. Calcium dependent activation of a multifunctional protein kinase by membrane phospholipids. J Biol Chem 1979;254:3692-5. Kuo JF, Andersson RGG, Wise BC, Mackerlova L, Salomonsson I, Brackett NL, et al. Calcium-dependent protein kinase: Widespread occurrence in various tissues and phyla of the animal kingdom and comparison of effects of phospholipid, calmodulin, and trifluoperazine. Proc Natl Acad Sci U.S.A. 1980;77:7039-43. Katoh N, Wrenn RW, Wise BC, Shoji M, Kuo JF. Substrate proteins for calmodulin-sensitive and phospholipidsensitive Cadependent protein kinases in heart, and inhibition of their phosphorilation by palmitoylcarnitine. Proc Natl Acad Sci U.S.A. 1981;78:4813-7.. hizuka Y. The structure, expression and properties of additional members of the protein kinase C family. J Biol Chem 1988;263:6927-32. Nishizuka Y. Intracellular signalling by hydrolysis of phospholipid and activation of protein kinase C. Science 1992;258:607-14. Brown JH, Martinson EA. Phosphoinositide-generated second messengers in cardiac signal transduction. Trends Cardiovasc Med 1992;2:209-14. Kaku T, Lakatta E, Filbnurn C. α-Adrenergic regulation of phosphoinositide metabolism and protein kinase C in isolated cardiac myocytes. Am J Physiol 1991;260:C635-42. Fuller SJ, Gaitanaki CJ, Sugden PH. Effects of catecholamines on protein synthesis in cardiac myocytes and perfused hearts isolated from adult rats. Biochem J 1990; 266:727-36. Bogoyevitch MA, Glennon PE, Andersson MB, Clerk A, Lazou A, Marhall CJ, et al. Endothelin-1 and fibroblast growth factors stimulate the mitogen-activated protein kinase signaling cascade in cardiac myocytes. J Biol Chem 1994;269:1110-9. Artman M. Developmental changes in myocardial contrctile responses to inotropic agents. Cardiovasc Res 1992; 26:3-13.. 16) 17) 18). 19). 20). 22) 23) 24) 25). 26). 27). 1349.
(10)
수치
관련 문서
The index is calculated with the latest 5-year auction data of 400 selected Classic, Modern, and Contemporary Chinese painting artists from major auction houses..
1 John Owen, Justification by Faith Alone, in The Works of John Owen, ed. John Bolt, trans. Scott Clark, "Do This and Live: Christ's Active Obedience as the
To evaluate the Effect of mutant RANKL on mRNA expressions in related with osteoclastogenesis,we investigated the expression of several osteoclast- specific genes both
Therefore, in this study, the conditions and problems of wind instrument programs of various junior high schools in the Gwangju region were analyzed
co-treatment with hispidulin and TGF-β up-regulated the protein of expression E-cadherin and occludin against TGF-β-induced in MCF-7 and HCC38 cells.. The
This study suggested diversity in the configuration of expression through expression of abstract images, the style of modern painting, which are expressed
This study observes the fictional realism used in The Magus and discusses how it is deployed in the short stories of The Ebony Tower differently.. Through this analysis,
Therefore, in this study, based on the media facade expression characteristics and expression techniques, evaluation factors through satisfaction analysis on
