48 책임저자:엄애선, 133-791, 서울시 성동구 행당동 17번지
한양대학교 생활과학대학 식품영양학과 Tel: 02-2220-1203, Fax: 02-2292-1226 E-mail: [email protected]
접수일:2009년 3월 18일, 게재승인일:2009년 3월 25일
Correspondence to:Ae-Son Om
Department of Food and Nutrition, College of Human Ecology, Hanyang University, 17, Haengdang-dong, Seongdong-gu, Seoul 133-791, Korea Tel: +82-2-2220-1203, Fax: +82-2-2292-1226
E-mail: [email protected]
Genistein이 MG-63 조골세포의 증식과 세포외 기질형성에 미치는 영향
한양대학교 식품영양학과, 1국립농업과학원 유해물질과
심재영ㆍ김인혜ㆍ백민경1ㆍ엄애선
Proliferation and Extra Cellular Matrix Formation Effects of Genistein on Human Osteoblast-like MG-63 Cell Line
Jae-Young Shim, In-Hye Kim, Min-Kyung Paik1 and Ae-Son Om
Department of Food and Nutrition, College of Human Ecology, Hanyang University, Seoul 133-791,
1National Academy of Agricultural Science, RDA, Suwon 441-707, Korea
We investigated the proliferation and extracellular matrix formation effects of genistein on MG-63 cell, human osteoblast like cell. The cells were treated with 1μM, 10μM, 100μM of genistein for 12 h, 24 h, 36 h, 48 h in order to determine the proliferation rate. The growth rate in control group increased up to 36 hours and then decreased at 48 h. Meanwhile, its rate increased rapidly up to at 24 h and then declined at 48 h by treating genistein. The effective time and concentrations of genistein for MG-63 cell was a 12 h and 10μM∼100μM, respectively. ALP and OC mRNA, initial bone proliferation marker, were strongly expressed by increasing concentration of genistein. The expression of Matrix metal- loproteinase (MMP)-13, a collagenase involved in extracellular matrix degradation, and tissue inhibitor of metalloproteinase (TIMP)-1 elevated at 10μM of genistein. Based on the results, genistein may affect the proliferation of osteoblast on MG-63 cell in initial time and increased the mineral deposition in the extra cellular matrix. Thus, genistein can play role in preventing bone loss during the initial osteoblast proliferation step and in promoting bone formation by inhibiting bone resorption which resulted from stimulation of the initial osteoblast proliferation due to the increased expression of ALP, OC, MMP-13 and TIMP-1. (Cancer Prev Res 14, 48-53, 2009)
Key Words: Genistein, Osteoblast, OC, MG-63 cell, ALP, MMP-13, TIMP-1
서 론
뼈는 태어나서 죽을 때까지 조골세포(osteoblast)와 파 골세포(osteoclast)에 의해 새로운 뼈가 형성되고 오래된 뼈는 분해되는 동적인 대사조직으로1) 조골세포와 파골 세포의 적절한 활성에 의해서 진행된다.2) 골다공증은 조 골세포에서의 골형성(bone formation) 관련 국소인자의 발
현 저하 및 파골세포의 골흡수(bone resorption) 작용의 상 대적인 증가에 의해 골밀도가 낮아지는 질환이다.3) 여성 의 경우 폐경 이후 에스트로겐 호르몬 분비의 감소로 남 성에 비해 골다공증의 발병률이 현저히 높으나 남성의 경우도 노화과정이 진행되며 골다공증의 발병률이 증가 한다.4) 그러나 최근 약물남용, 무리한 체중감량, 갑상선 비대증, 신장병, 당뇨병 등과 같은 만성질환의 증가, 이 른 폐경기, 젊은 여성들의 질환으로 인한 난소절제 등의
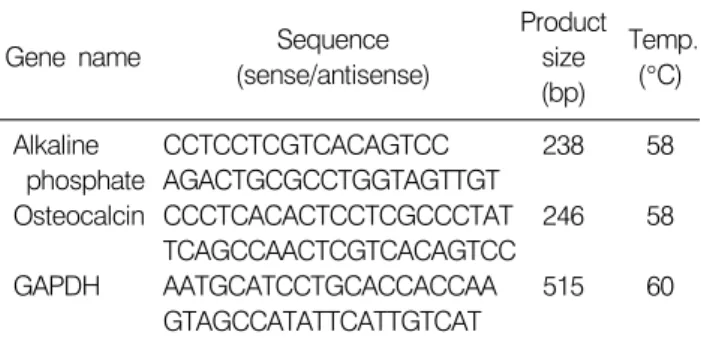
Table 1. Gene-specific primer for RT-PCR
Gene name Sequence
(sense/antisense)
Product size (bp)
Temp.
(°C) Alkaline
phosphate Osteocalcin GAPDH
CCTCCTCGTCACAGTCC AGACTGCGCCTGGTAGTTGT CCCTCACACTCCTCGCCCTAT TCAGCCAACTCGTCACAGTCC AATGCATCCTGCACCACCAA GTAGCCATATTCATTGTCAT
238 246 515
58 58 60 요인들에 의한 칼슘과 비타민 D의 흡수 장애 및 부갑상
선 호르몬과 에스트로젠의 불균형 등으로 젊은 남녀성 인 뿐만 아니라 성장기 어린이까지 골다공증의 발병 위 험이 높아지고 있다.5,6)
골형성은 multipotential stem cell의 골형성 세포로의 변 환, 조골세포로의 분화, 골기질 형성과 광물화를 요구하 는 복잡한 생물학적 과정이다. 골은 강화된 특수 결합조 직으로서 33%가 유기 기질로 되어 있으며, 이중 28%는 제 1형 교원성 단백질이고 나머지 5%는 비교원성 단백 질로 되어있다.7) 교원성 단백질은 비교원성 단백질에 의 해 광물화가 일어날 수 있는 주형을 제공하여 골형성의 초기에 작용하며 alkaline phosphate (ALP)는 조골세포의 분화와 골기질의 성숙에 중요한 역할을 하며 osteocalcin (OC)는 거의 독점적으로 조골세포에 의하여 생성되며 활성이 활발한 조골세포의 특이 표지자로 알려져 있는 골기질 단백질이다.8) 또한 조골세포에서의 MMP는 파골 세포가 활성이 일어나기 전 골의 석회화를 제거하는 초 기 단계에서 골재흡수를 조절하는 역할을 한다.9) 이러한 골대사는 인종, 연령, 성별, 내분비호르몬, 신 체활동 등 유전적, 생리적 및 환경적 요인, 단백질, 미네 랄, 비타민, 식이 섬유소 등과 같은 영양적인 요인에 의 해 직간접적으로 영향을 받는다.10) 이에 골다공증의 예 방 및 치료를 위한 대표적인 대체요법으로 식물성 에스 트로젠의 경구 투여 또는 이를 다량으로 함유하고 있는 식품의 섭취가 시도되고 있다.11) 이소플라본(isoflavone)은 식물성 에스트로젠의 대표적인 성분으로 aglycone인 ge- nistein, daidzein, glycitin과 이들에 당이 결합된 배당체 등 지금까지 12종류가 밝혀져 있는데 이중 genistein은 에스 트로젠 효과뿐만 아니라 항에스트로젠 효과를 동시에 가지고 있다. 따라서, 최근 에스트로젠 보충요법이 여러 가지 부작용을 유발한다는 보고가 계속되고 있으므로 genistein은 골다공증 예방을 위한 대체물질로 각광받고 있다.12) 그러나 genistein의 직접적인 골형성 유도에 관한 국내 연구자료의 진행은 활발하지 않다. 이에 조골세포 에서 genistein의 골 형성 능과 기질 형성 능을 본 연구에 서 규명하고자 하였다.
재료 및 방법 1. MG-63 세포의 배양 및 증식 측정
MG-63 human osteoblastic cell은 한국 세포주 은행(서울 대학교 의과대학)에서 분양받아 본 연구실에서 계대 배 앙하여 사용하였다. MG-63세포는 10% fetal bovine serum (FBS, GIBCO, NE, USA)를 함유한 Dulbecco's modified Ea-
gle's medium (DMEM)에서 37°C CO2-incubator (Forma, mo- del 311 S/N29035, USA)를 이용하여 배양하였다. MG-63 세포를 96 well plate에 2×104으로 분주하고 genistein을 1, 10, 100μM의 농도에서 12, 24, 36, 48시간 동안 배양하였 다. 배양된 세포에 MTT (20μL/well) 시약을 첨가하고 2시 간 동안 37°C에서 배양한 후 MTT가 formazan으로 분해되 는 양을 ELISA reader를 이용하여 540 nm에서 흡광도를 측정하여 결정하였다. 각각의 처리 군을 6회 반복측정하 여 결정하였다. Genistein에 대한 세포 증식효과는 반복 실험의 평균값을 취한 후 미처리된 DMEM 용액에 배양 한 대조군에 대한 백분율로 표시하였다.
2. Reverse transcription polymerase chain re- action (RT-PCR)을 통한 골형성 표지자 분석
동일한 조건에서 배양된 MG-63 세포에 TRI reagent (BIO- RAD, CA, USA)을 가한 후 4°C, 12,000 rpm에서 15분간 원심 분리를 시행하였고, 상층액에 0.2 mL의 chloroform을 가 한 후 4°C, 12,000 rpm에서 15분간 원심분리하였다. 이 시편의 상층액에 0.6 mL의 isopropanol을 가하여 12,000 rpm에서 10분간 원심분리하고 추출된 RNA은 75% ethanol 로 세척하였다. 추출된 RNA를 정량화한 후 oligo dT primer와 AMV reverse transcriptase를 이용하여 1μg의 RNA에서 cDNA를 합성하였다. 이 cDNA를 사용하여 관찰대상 유전자(Table 1)를 polymer chain reaction 방법으로 증폭하였다. 증폭된 산물은 1.5% agarose gel을 이용하여 전기영동하고 ethidium bromide로 염색한 후 UV하에서 확인하였다.
3. Von Kossa 염색을 통한 골기질 형성 분석
동일한 조건에서 배양된 MG-63 세포를 PBS로 세척하 고 4% formalin으로 10분간 고정하였다. 증류수로 세척한 후 5% AgNO3을 처리하고 30분간 암실에 보관하였다. 과 잉의 AgNO3를 증류수를 수회 세척한 후 NaHCO3/ for- malin buffer를 7분간 반응시키고 5% Na2S2O3으로 여분의
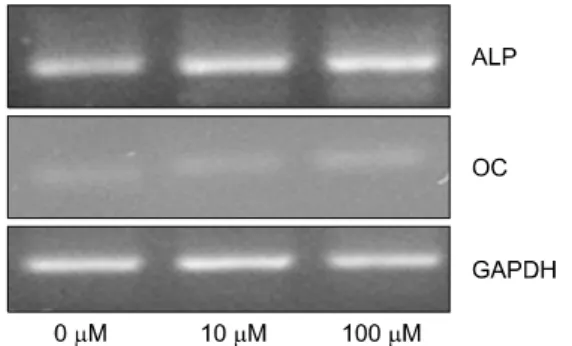
Fig. 2. Effects of genistein on ALP (alkaline phosphatase) mRNA level, OC (osteocalcin) mRNA level in MG-63 cell. OC and ALP were measured by RT-PCR.
Fig. 1. Proliferation on MG-63 cell by time course and dose dependant. Cell viability of MG-63 cells cultured with genistein 1μM, 10μM and 100μM from 12 hours to 48 hours. The number of cells was measured at MTT assay. a and b means within a raw with no common superscripts differ significantly (p<0.05). Data expressed as the mean±SD (n=6).
AgNO3를 중성화시킨 후 색의 변화를 관찰하였다.
4. Western blot analysis을 통한 세포외 기질 분해 분석
동일한 조건에서 배양된 MG-63 세포를 1 mL의 RIPA buffer (20 mM Tris-HCl, pH 8, 150 mM NaCl2, 10 mM NaPO4, 10% glycerol, 100 Na3VO4, 100 M ammonium molybabdate, 1% NP-40, 0.1% SDS)에 4°C 상태로 세로를 풀어놓고, 초음 파 파쇄기를 이용하여 낮은 강도로 5초 동안 10회 반복 하여 세포벽을 파괴시킨 후 균질액을 15,000 g에서 20분 간 원심분리하여 상층액을 단백질 추출액으로 준비하였 다. 준비된 단백질을 Bio-Rad protein assay로 정량화한 후 6X sample buffer에 혼합하고 95°C에서 끓인 다음 10%
polyacrylamide/SDS gel에서 전기영동시킨 후 nitrocellulose membrane에 흡착시켰다. 1차 항체 MMP-1, 13, TIMP-1 (Santa Cruz Biotech, CA, USA)는 5% skim milk, 0.1% Tween 20을 함유한 PBS에 희석시켜서 실온에서 2시간 동안 반 응시킨 후 0.01% Tween 20을 함유한 PBS로 5분간 5회씩 세척하였다. Blocking solution으로 희석시킨 이차항체(pe- roxidase conjugated anti-IgG)를 실온에서 1시간 동안 반응 시킨 후 0.01% Tween 20을 함유한 PBS로 5분간 5회씩 세척하였다. 발색은 ECL kit로 확인하였으며 강도는 Gel- Documation (EDAS290, Kodack Co., Japan)를 사용하여 정 량화하였다.
5. 통계분석
자료 분석은 SPSS (SPSS Inc. CA, USA) program을 이용 하여 통계분석을 시행하였다. 모든 변수의 평균과 표준 편차를 구하였으며, 군별 차이는 One-way ANOVA의 Duncan’s multiple range test를 이용하여 분석하였다.
결과 및 고찰
1. Genistien에 의한 MG-63 세포 증식 측정
MG-63 세포에서 genistein의 생존 및 증식에 미치는 효 과를 알아보기 위해 MTT assay를 실시한 결과는 Fig. 1에 나타내었다. 여러 차례 시행한 예비실험 결과에 의해 세 포 독성이 나타나기 시작하는 고농도를 제외하고 시료 처리 농도(1μM, 10μM, 100μM)를 선택하였으며, 증식 변화를 관찰할 수 있는 시간(12시간, 24시간, 36시간, 48 시간)을 선택하였다. MG-63 세포의 증식률은 미처리된 배양액만 처리한 대조군에 대한 비율로 나타내었다(Fig.
1). Genistein을 처리할 경우 12시간에 모든 농도군의 증 식율이 대조군에 비하여 유의적(p<0.05)으로 증가하였 으며 24시간에는 10μM에서, 36시간에는 100μM에서 유 의적(p<0.05)으로 증가하였으나 48시간에는 유의적으로 감소하였다. 따라서, 이후 실험에서는 10μM, 100μM의 농도와 12시간의 반응시간을 선택하였으며 본 실험에서 사용한 genistein의 농도와 반응 시간은 genistein이 조골세 포에서의 분화능 효과를 실험하고자 하였던 다른 연구13) 와도 유사하였다.
2. Genistien에 의한 MG-63 세포의 mRNA ALP 변화
MG-63 세포에 10μM, 100μM의 genistein을 12시간 동 안 처리하여 ALP의 발현 양상을 RT-PCR로 측정한 결과 는 Fig. 2와 같다. ALP는 거의 모든 조직에 존재하지만
Fig. 3. Effects of genistein on mineralization in MG-63 cell. Cell was stained by Von Kossa Stating Solution.
Fig. 4. Effects of genistein on MMP (matrix metallopro- teinase)-1,13 and TIMP (tissue inhibitor of metllaproteinase-1) protein expression in MG-63 cell. MMP and TIMP were measured by Western Blotting.
특히 골조직에서는 골 성장이 활발히 일어날 때 그 활성 이 증가한다.14) ALP는 비교원성 단백질로 유기인산 에스 테르를 가수분해하여 골형성과 재생이 일어나는 부위의 국소적 인산 이온의 농도를 증가시킨다.15) 그러므로 조 골세포 활성을 알아보는 표지자로 MG-63 세포에서의 ALP 활성을 측정함으로써 genistein이 조골세포의 활성에 미치는 영향을 알아보았다. 본 실험에서는 genistein의 처 리에 의해 ALP의 발현이 증가되었으며 농도 10μM에서 가장 높았다. 이는 초기 세포의 증식단계에서 여러 층을 형성하는 골결절 형성의 초기에 도움을 줄 것으로 예상 되며 증가된 ALP 합성은 세포외기질에 calcium phosphate 를 침착시켜 골형성의 핵 역할을 함으로써16) 석회화를 유도하여 조골세포의 분화와 성숙에 주요한 기능을 할 것으로 판단된다.
3. Genistien에 의한 MG-63 세포의 mRNA OC의 변 화
MG-63 세포에 10μM, 100μM의 genistein을 12시간 동 안 처리하여 OC의 발현 양상을 RT-PCR로 측정한 결과 는 Fig. 2와 같다. OC는 골형성의 생화학적 표지자인 비 타민 K 의존성 단백질로써 조골세포에서 합성된다. OC 는 비교원성 단백질로 혈중 농도가 골형성 여부를 확인 할 수 있는 요소이기도 하다.17) 본 연구에서 OC는 ge- nistein의 농도가 증가할수록 발현이 증가하는 것으로 관 찰되었으나 ALP에 비교해서는 크게 증가하지 않았다.
OC는 골, 상아질 및 석회화된 연골에서 발현되는 매우 특이적인 유전자로서, 골에서는 성숙된 조골세포에서만 발현되는 후기 조골세포 인자이다.18) OC의 발현은 골이 석회화되어 결절을 형성하는 시기와 일치하며, 이는 골 기질의 석회화를 촉진시키는 요인으로 작용한다.19)
Uchinama의 연구20)에서 genistein은 OC의 발현에 영향을 주지 않았으나 본 연구실에서는 처리된 genistein의 농도 가 증가할수록 발현이 증가되었다.
4. Von Kossa 염색을 통한 골기질 형성 분석
무기질이 침착된 기질을 갈색으로 표현되게 하는 Von Kossa 염색을 통하여 골기질 형성정도를 측정한 결과는 Fig. 3과 같다. Genistein의 농도가 증가함에 따라 무기질 의 침착이 증가하는 것이 관찰되었다. 이러한 결과는 이 미 언급된 ALP와 OC의 발현에 대한 일반적인 사실과도 일치하여 조골세포의 활성이 증가되었다는 것을 볼 수 있다.
5. Genistien에 의한 MG-63 세포의 세포 외 기질분 해의 변화
MG-63 세포에 10μM, 100μM의 genistein을 12시간 동 안 처리하여 MMP-1,13과 TIMP-1의 단백질 발현 양상을 Western Blotting을 이용하여 측정한 결과는 Fig. 4와 같 다. MMP-13와 TIMP-1은 10μM 농도의 genistein 에서 높 게 발현되었다. 일부 MMP는 조골세포에서 발현되기도
하며 in vitro에서 골 분해 중 MMP-1, 2, 9, 13과 TIMP-1, 2의 발현이 관찰된다는 연구도 있다. 또한 MMP-13은 조 골세포의 분화와 무기질화하는 동안 증가하며 Cbfs1에 의해 조절된다.21) 본 연구에서는 genistein에 의해 조골세 포의 MMP-13과 TIMP-1이 증가되어 조골세포가 분화하 여 무기질화하였다고 볼 수 있으며 이는 이미 언급한 골 기질 형성과도 일치한다.
결 론
본 연구에서는 조골세포에서 genistein의 골형성능과 기질 형성능의 기전을 규명하고자 하였다. MG-63 세포 에 genistein을 1μM, 10μM, 100μM의 농도 별로 처리하 여 각각 12시간, 24시간, 36시간, 48시간에 증식 변화를 측정하였다. 대조군의 성장은 36시간까지 증가하다 48 시간에 감소하였다. 반면 genistein을 투여한 경우 24시간 까지 급속하게 증식율이 증가하다 48시간에 감소하였 다. MG-63 세포에 10μM, 100μM의 genistein을 12시간 동안 처리하여 ALP와 OC의 발현 양상을 RT-PCR로 관찰 한 결과 초기 조골세포 분화 지표인 ALP의 경우 genistein 의 농도가 증가할수록 발현이 증가되었으며, 후기 조골 세포 분화 지표인 OC의 경우 ALP에 비해서는 약하지만 발현이 증가되었다. 세포 외 기질의 무기질 침착 변화를 측정한 결과 genistein의 농도 증가에 따라 무기질 침착이 증가하였다. MMP-1, 13과 TIMP의 단백질 발현 양상을 Western Blotting을 이용하여 변화를 측정한 결과 세포기 질 분해효소인 MMP-13, TIMP-1은 genistein 10μM에서 발현이 증가하였다. 따라서 human osteoblast-like cell인 MG-63 cell에 genistein 12시간 동안 10μM과 100μM을 처 리한 결과 genistein이 MG-63 cell의 초기 조골세포 분화능 에 효과를 보이는 것으로 생각되며, 세포 기질의 무기질 화를 촉진하여 골손실을 예방하는 것으로 생각된다.
감사의 글
본 연구는 한국학술진흥재단의 기초과학지원사업 (MOEHRD, Basic Research Promotion Fund, R04-2004-000- 10237-0)에 의해 이루어졌습니다.
참 고 문 헌
1) 최혜미, 김정희, 이주희, 김초일, 송경희, 장경자, 민혜 선, 임경숙, 변기원, 송은승, 여의주, 이홍미, 김경원, 김 희선, 김창임, 윤은영, 김현아. 21세기 영양학. 파주시
교문사. pp 334-347, 2000.
2) Seibel MJ. Biochemical markers of bone turnover part I:
biochemistry and variability. Clin Biochem Rev 26, 97-122, 2005.
3) Mohan S, Baylink DJ. Bone growth factors. Clin Orthop Relat Res 263, 30-48, 1991.
4) Duda RJ, O’Brien JF, Kslxmsnn JA. Concurnet assays of circulating bone-gla protein and alkaline phosphate: Effect of sex, age, and metabolic bone disease. J Clin Enfocr Meta 66, 951-957, 1988.
5) 김경희. 성장기 어린이의 골밀도에 영향을 미치는 요인 에 관한 연구. 한양대학교 석사학위논문. 1999.
6) Paik MK, Lee HO, Chung HS, Yang SO, Om AS. Genistein may prevent cadmium-induced bone loss in ovariectomized rats. JMF 6, 337-343, 2003.
7) Urist MR. Bone formation by autoinduction. Science 150, 893-899, 1965.
8) Robey PG, Biano P, Termine JD. The cellular biology and molecular biochemistry of bone formation. In: Coe FL, Favus MJ, eds. Disorders of Mineral Metabolism. New York, Raven Press, pp 241-263, 1992.
9) Filanti C, Dickson GR, Di Martino D, Ulivi V, Sanuineti C, Romano P, Palerno C, Manduca P. The expression of me- talloproteinase-2,9, and -14 and of tissue inhibitors-1 and -2 is developmentally modulated during osteogenesis in vitro, the mature osteoblastic phenotype expressing metalloprotei- nase-14. J Bone Miner 15, 2154-2168, 2000.
10) Williams MH. Nutrition for health and fitness. McGraw Hill pp 611-630, 2000.
11) Anderson JJB, Garner SC. Phytoestrogens and bone. Baillieres Best Tract Res Clin Endocrinol Metab 12, 543-557, 1998.
12) Cooke PS, Selvaraj V, Yellayi S. Genistein, Estrogen Re- ceptors, and the Acquired Immune Response. J Nutr 136, 704-708, 2006.
13) Zhang Y, Jin H, Xu Z, Nan W. Role of c-jun in the effect of genistein on neonatal rat calvaria osteoblast proliferation and differentiation. Wei Sheng Yan Jin 37, 308-310, 2008.
14) Broskey AL. Non-collagen matrix proteins and their role in mineralization. Bone Miner 6, 111-123, 1992.
15) Bellow CG, Aubin JE, Heersche JNM. Initiation and progression of mineralization of bone nodules formed in vitro:
The role of alkaline phosphate and organic phosphate. Bone and Mineral 14, 27-40, 1991.
16) Wang J, Shang F, Jiang R, Liu L, Wnas S, Hou J, Huan M, Mei Q. Nitric oxide-donating genistein prodrug: design, synthesis, and bioactivity on MCT3T3-E1 cells. J Pahrmacol Sci 104, 82-89, 2007.
17) Safadi FF, Xu J, Smock SL. Expression of connective tissue growth factor in bone: its role in osteoblast proliferation and differentiation in vitro and bone formation in vivo. J Cell Physiol 196, 51-60, 2003.
18) Lawton DM, Andrew JG, Marsh DR. Expression of the gene encoding the matrix gla protein by mature osteoblasts in
human fracture non-uniuns. Mol Pathol 52, 92-101, 1999.
19) Hauschka PV, Lian JB, Cole DEC, Gundberg CM. Osteo- scalcin and matrix gla protein: vitamin K-dependent proteins in bone. Physiol Rev 69, 990-1047, 1989.
20) Uchinama S, Yamagucho M. Anaboic of B-cryptozanthin in osteoblastic MC3T3-E1 cells is enhanced with 17β-estradiol, genistein, or zinc sulfate in vitro: the unique effect with zinc
on Runx2 and α1 (I) collagen mRNA expressions. Mol Cell Biochem 307, 209-219, 2008.
21) Uchida M, Shima M, Shimosha T, Fujieda A, Obara K, Suzuki H, Nagai Y, Ikeda T, Yamato H, Kawaguchi H. Regulation of matrix metalloproteinases (MMPs) and tissue inhibitors of metalloproteinases (TIMPs) by bone resorptive factors in osteroblastic cells. J Cell Physiol 185, 207-114, 2000.