37
<원례보저>
최근 국내 분리 고병원성 infectious bursal disease virus의 segment A 유전자 특성
오현석·이진화·권혁무·성환우*
강원대학교 수의과대학 (게재승인: 2010년 9월 30일)
Sequence analysis of segment A gene of a very virulent infectious bursal disease virus recently isolated in Korea
Hyun Seok Oh, Jin Hwa Lee, Hyuk Moo Kwon, Haan Woo Sung*
College of Veterinary Medicine, Kangwon National University, Chunchon 200-701, Korea
(Accepted: September 30, 2010)
Abstract : Infectious bursal disease virus (IBDV) is a member of the
Avibirnavirusgenus of the
Birnaviridaefamily which genome consists of two segments (A and B) of double stranded RNA. Segment A gene of KNU08010 isolate, which was isolated from a 15-day-old chicken flock in 2008, was sequenced and compared with other IBDV isolates including SH/92 strain, the first Korean very virulent (vv) IBDV isolate.
The amino acid sequences of segment A gene showed that KNU08010 had 99.2% homology with SH92 strain. KNU08010 isolate had specific amino acids A222, I242, I256, I294 and S299 which are highly conserved among vvIBDV strains. Phylogenetic analysis based on the nucleotide sequences of variable region of the VP2 gene of 18 IBDV strains revealed that KNU08010 was grouped with vvIBDVs and was closely related to Korean vvIBDVs isolated from wild birds.
Keywords : segment A gene, sequence, very virulent IBDV
서 론
전염성
F
낭병(Infectious bursal disease, IBD) virus
(IBDV)
는감수성조류에감염하여면역능력저하와,
증체저하및폐사를유발한다
[9]. IBDV
는혈청형(serotype) I
과II
로구분되는데혈청형II
는닭과칠면조에감수성 이있으나병원성은나타나지않고,
혈청형I
바이러스가주로닭에서병원성을나타낸다
[14, 21].
혈청형I
바이러스에는기존바이러스
(classical IBDV)
뿐만아니 라기존바이러스와 항원성이변화된 항원성변이형(antigenic variant subtype)
과병원성이변화된병원성변 이형(pathogenic variant subtype)
으로구분될수있다[9].
병원성변이형중치사율이매우높은
very virulent(vv)
IBDV
는1980
년대후반유럽에서처음발생보고된이후여러나라에서발생되고있으며국내에서도
1992
년 처음발생된사실이보고된바있다[1, 8, 23, 26].
IBDV
는Birnaviridae family
에 속하는 바이러스로RNA
는두개의분절(segment)
로구성되어있으며큰분 절인segment A
는2
개의open reading frame(ORF)
를가지고있고이중
large ORF
는polyprotein
으로번역된후viral protease
에의해VP2(37 kD), VP4(32 kD), VP3(28 kD)
로가공된다[6, 12, 13, 22]. VP2
는숙주의중화항체를유도하는항원결정부위를갖고있는구조단백질로
이중아미노산
206~350
에해당하는부위는다양한변이가관찰되어
hypervariable region(HVR)
이라고부른다[3, 5, 10, 15].
또한,
아미노산326~332
에존재하는serine rich heptapeptide(SWSASGS)
는모든병원성바이러스(virulent strain)
에서보존되어있고병원성이약한바이*Corresponding author
Tel: +82-33-250-8680, Fax: +82-33-244-2367 E-mail: [email protected]
러스에는
serine
기가적은것이특징이다[11]. HVR
에 나타나는A222, I242, I256, I294, S299
아미노산은지역과국가에관계없이대부분의
vvIBDV
에서보존되어있으며
[15],
국내vvIBDV
에서도 확인된바있다[18].
VP3
는중화항체와는관련없는group-specific antigen
으로혈청형에관계없이염기서열이유사하며
, polyprotein
을가공하는
protease
인VP4
는VP3
와마찬가지로염기 서열이대부분잘보존되어있다[3, 7, 25].
본연구는기존의
IBD
사례와는다르게2
주령전후 의닭에서높은치사율을보이는사례에서분리된바이 러스의유전자특성을파악하여기존바이러스와의차 이점을구명하고자실시하였다.
재료 및 방법
IBDV의분리및증폭
IBDV
분리및증폭은specific pathogen free(SPF)
종란에접종하는방법으로실시하였다
.
즉,
바이러스분리 용채취시료인F
낭조직을항생제가첨가된멸균PBS
로
20%(w/v)
장기유제액을만든뒤3,000 rpm
에서10
분간원심분리하고상층액을채취하여
0.45 µm membrane filter(Millipore, USA)
에여과한후10
일령SPF
종란의chorioallantoic membrane(CAM)
에0.2 mL
씩접종하였다. 5
일간배양후계태아에서의IBDV
특이병변을확인하 고CAM
을채취하여실험에사용하였다.
Viral RNA 추출및염기서열분석
바이러스
RNA
는TRIZOL(Invitrogen, USA)
를사용하여제조사의 설명에따라 추출하였다
. Segment A gene
염기서열을분석하기위하여Kwon
등[18]
이보 고한 것과 동일한primer 5
쌍을 이용하여reverse transcriptase (RT)-polymerase chain reaction(PCR)
을통 해segment A gene
을증폭하였다. IBD viral RNA
는이 중쇄(double strand) RNA
이기 때문에 추출한IBDV dsRNA
는90% demethyl sulfoxide
에부유시킨후98
oC
에서
5
분간반응시켜단일가닥으로denaturation
시킨후42
oC
에서60
분간RT
반응을수행하였다. PCR
은94
oC 5
분간denaturation, 51
oC 30
초간annealing, 72
oC 2
분간elongation
과정을35
회 반복 수행하였으며 최종elongation
은74
oC 15
분간실시하였다. RT
와PCR
반응에사용한시약은
Accupower RT/PCR Premix(Bioneer, USA)
를사용하였다.
PCR
로증폭된DNA
는1.5% agarose gel
에전기영동으로전개시킨후
IBDV
특이증폭산물을수거하였다.
수거된
gel
로부터QiAquick Gel Extraction Kit(Qiagen, USA)
을 이용하여 증폭DNA
를 정제하고Macrogen (Korea)
사에증폭DNA
와primer
를제공하여염기서열 을분석하였다.
분석된염기서열은MEGA4.0
과Bioedit software
를이용하여다른바이러스(Table 1)
와의관련성을분석하였다
.
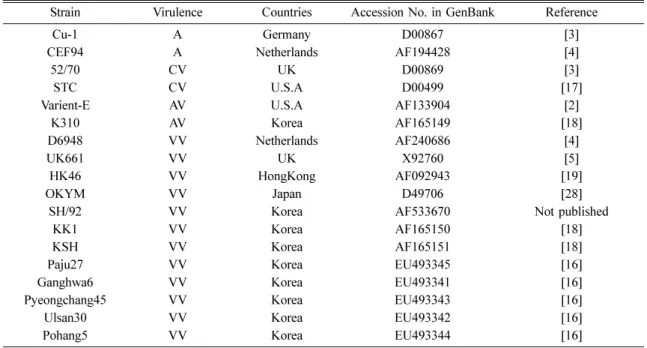
Table 1. IBDV strains used for alignment and comparison of homology of nucleotides and amino acids in this study
Strain Virulence Countries Accession No. in GenBank Reference
Cu-1 A Germany D00867 [3]
CEF94 A Netherlands AF194428 [4]
52/70 CV UK D00869 [3]
STC CV U.S.A D00499 [17]
Varient-E AV U.S.A AF133904 [2]
K310 AV Korea AF165149 [18]
D6948 VV Netherlands AF240686 [4]
UK661 VV UK X92760 [5]
HK46 VV HongKong AF092943 [19]
OKYM VV Japan D49706 [28]
SH/92 VV Korea AF533670 Not published
KK1 VV Korea AF165150 [18]
KSH VV Korea AF165151 [18]
Paju27 VV Korea EU493345 [16]
Ganghwa6 VV Korea EU493341 [16]
Pyeongchang45 VV Korea EU493343 [16]
Ulsan30 VV Korea EU493342 [16]
Pohang5 VV Korea EU493344 [16]
IBDV: infectious bursal disease virus, A: attenuated, CV: classical virulent, AV: antigenic variant, VV: very virulent.
결 과
발생사례및바이러스분리
충남당진소재한농장으로부터
IBDV KNU08010
주 를분리하였다.
바이러스가분리된농장은약15,800
수의토종닭이입추되어사육되던농장으로
14
일령부터 폐사가크게증가되기시작하여연구자의실험실로가 검물이의뢰된사례이었다. 15
일령때의뢰된 가검물은부검결과
F
낭의종대및출혈,
선위근위연접부위의출혈
,
피하의반상출혈등이특징으로전형적인IBD
감염사례로판단되었다
.
폐사수는10
일간약2,200
수가폐사하여총
13.9%
의폐사가있었으며폐사양상은급격히폐사수가증가하였다가감소하는양상을보였던사례이 었다
.
부검후F
낭을 채취하여10
일령의SPF
종란의CAM
에접종후5
일간배양한결과계태아의부종,
출혈 및간의괴사반점등IBDV
특이병변이관찰되어IBDV
를확인할수있었다
.
분리된IBDV
는SPF
종란의CAM
에서증폭하여유전자염기서열분석에사용하였다
.
염기서열분석결과
분절
A
유전자전체염기3,039
개중분리주KNU08010
는
VP2
시작부위일부와VP3
종결부위의일부염기30
개를제외한총
3,009
개의염기서열을분석하였다.
분석한
3,009
개의염기서열을기준으로다른바이러스와의상동성을비교한결과
KNU08010
주는일본에서분리보고된
vvIBDV
인OKYM
주와국내에서분리된vvIBDV
인
SH/92
주와각각98.9%
및98.6%
로높은 상동성을보였으며국내분리주
KK1
과는96.0%
의상동성을보였다
(Table 2).
또한KNU08010
주는classical IBDV
로분류되고있는
STC
주와는염기상동성이95.1%
이었으며, avirulent strain
인Cu1
주와는94.7%
의상동성을보였고, antigenic variant
바이러스로알려진Var-E
주와K310
주와는각각
95.2%
와93.9%
로vvIBDV
보다는낮은상동 성을보였다.
염기서열을토대로 예상되는아미노산의상동성비교결과에서도
KNU08010
주는OKYM
주및SH92
주와는 각각99.6%
와99.2%
의높은상동성을보였으며
, KK1
주와는97.4%, STC
와는96.2%, Cu1
주와는96.7%, Var-E
주와K310
주와는 각각94.0%
와93.5%
의 아미노산상동성을보였다(Table 2).
KNU08010
주는모든병원성바이러스주에서보존되고 있는
HVR
의 아미노산326-332
부위의serine rich heptapeptide(SWSASGS)
를가지는특징을보였으며VP2
단백질
(
아미노산1-454)
중아미노산201-350
에해당하 는HVR
에서다른분리주와의차이가 뚜렷하였다(Fig.
1). KNU08010
주는HVR
부위에서vvIBDV
간에잘보존되어있는것으로알려진
5
개의아미노산(A222, I242, I256, I294, S299)
이모두존재하였다. KNU08010
주는polyprotein
전체에서다른vvIBDV
주(SH/92, OKYM)
에서만공통적으로관찰되는아미노산을조사한결과
14
개
(A222, I242, I256, A270, I294, S299, L451, I541, Y680, N685, S715, D751, V990, A1005)
가확인되었다.
이중7
개
(A222, I242, I256, A270, I294, S299, L451)
는VP2(polyprotein 1~454), 4
개(I541, Y680, N685, S715)
는VP4(polyprotein 455~723),
나머지3
개(D751, V990, A1005)
는VP3(polyprotein 724~1012)
에존재하였으며L451
은VP2-VP4
절단부위근처에위치하였다(Fig. 1).
VP2
에서나타나는특징적인7
개의아미노산중6
개(A222, I242, I256, A270, I294, S299)
는HVR
에서확인 되었다. KNU08010
주는HVR
에서다른분리주와의차 이가가장뚜렷하였으며antigenic variant
주인K310
주와는가장많은
21
개의아미노산차이가 있었다.
국내에 서분리된vvIBDV
인SH/92
및KK1
주와는HVR
에서는 거의동일하였으나SH/92
주와는D279N, KK1
주와는F266L
과M290V
에서차이를보였다.
국내분리주간에polyprotein
전체의아미노산을비교한결과KNU08010
주는
SH92
주와는4
개의차이만있었으나KK1
주와는비교적많은차이를보여
17
개의아미노산차이를보였다.
야생조류를포함한
IBDV 18
개주를대상으로HVR
의Table 2. Nucleotide and deduced amino acid sequence similarities of segment A gene of KNU08010 with other IBDV strains
KNU08010 OKYM SH/92 KK1 Var-E STC Cu1 K310
KNU08010 98.9
*98.6 96.0 95.2 95.1 94.7 93.9
OKYM 99.6
†99.0 96.4 95.1 95.6 95.4 94.1
SH/92 99.2 98.9 96.4 95.2 95.6 95.5 94.3
KK1 97.4 96.8 96.4 95.6 96.4 97.3 96.3
Var-E 94.0 93.7 93.7 93.0 96.7 96.8 95.7
STC 96.2 95.9 95.9 95.0 93.9 97.7 96.3
Cu1 96.7 96.4 96.4 96.1 95.0 97.5 97.7
K310 93.5 93.2 93.1 93.2 92.1 94.3 95.7
*
Percent homology of nucleotide sequences.
†Percent homology of amino acid sequences.
염기서열을기준으로
phylogenetic tree
를작성하여근연관계를분석하였다
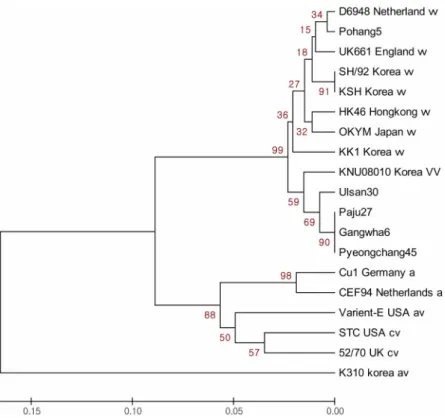
(Fig. 2).
분리주KNU08010
주는국내외보고된
vvIBDV
와동일한계열로분류되었고,
특히야생조류에서 분리된
vvIBDV
인Pohang5, Ulsan30,
Paju27, Gangwha6, Pyeongchang45
와가장가까운근연관계를 나타냈다
.
반면, classical IBDV(STC, 52/70), avirulent IBDV(Cu1, CEF94), antigenic variant strain(Var- E)
과는다른계열로분류되었다.
Fig. 1. Deduced amino acid sequence translated from open reading frame 2 in segment A gene of infectious bursal disease virus (IBDV) strains. Differences from the consensus sequence are shown. Identical amino acids are marked as a period (.). Hyphens indicate positions where amino acids are not confirmed in KNU08010. Hydrophilic regions, which represent areas critical for antigenicity, are boxed. The serine-rich region is underlined.
Fig. 2 . Phylogenetic tree based on the nucleotide sequences of variable region (nucleotides 571-1277) of the VP2 gene
of 18 IBDV strains denoted from Table 1. a: attenuated, cv: classical virulent, av: antigenic variant, vv: very virulent.
고 찰
vvIBDV
는폐사율이높을뿐만아니라기존의classical IBDV
와는다르게폐사를유발하는감수성일령범위가 넓은특징이있다.
즉, vvIBDV
는3
주령미만이나14
주령전후의닭에 감염하여도 폐사를 유발할수있어
classical IBDV
와는 차이를 보인다.
본 실험에서KNU08010
주가분리된농장에서는폐사가2
주령부터나타나기시작하였으며폐사한병아리의부검결과
F
낭의 종대및출혈,
선위근위연접부위의출혈,
피하의반상 출혈등IBD
의임상병변이나타나는것으로보아3
주령미만의병아리에감염될경우임상병변이거의나타 나지않던
classical IBDV
감염과는차이를보였다.
KNU08010
주는모든병원성바이러스주에서보존되고 있는
HVR
의 아미노산326-332
부위의serine rich heptapeptide(SWSASGS)
를가지는특징을보였다.
또한VP2
의HVR
부위에서vvIBDV
간에잘보존되어있는것으로 알려진
5
개의 아미노산(A222, I242, I256, I294, S299)
이모두존재하였다. Jackwood
와Sommer-Wagner [15]
는4
개대륙18
개국에서분리된vvIBDV
를대상으로
HVR
부위에서의아미노산특징을조사한결과대부 분의vvIBDV
는위의5
개아미노산이잘보존되고있음 을확인한바있다.
본실험에서분리된KNU08010
주도이부위
5
개의아미노산이모두잘보존된것으로보아vvIBDV
로추정되었다.
KNU08010
주는polyprotein
전체에서다른vvIBDV
주(SH/92, OKYM)
에서만공통적으로관찰되는아미노산을조사한결과
14
개(A222, I242, I256, A270, I294, S299, L451, I541, Y680, N685, S715, D751, V990, A1005)
가확인되었다
.
이중7
개(A222, I242, I256, A270, I294, S299, L451)
는VP2, 3
개(I541, Y680, N685, S715)
는VP4,
나머지
3
개(D751, V990, A1005)
는VP3
에존재하였다.
IBDV
의병원성은VP2
단백질의변화뿐만아니라다른부위단백질변화와도 관련이있음을시사한바있다
[6, 20, 28, 29].
따라서KNU08010
주의VP3
혹은VP4
부위에서특징적으로나타나는이들아미노산이
vvIBDV
의병원성과관련이있는지여부는앞으로추가적인연 구가더필요할것으로생각되었다
.
Cystein
과proline
기는바이러스단백질의folding
에관 여한다고알려져있으며VP2
의아미노산222
와VP4
의아미노산
715
에위치하는proline
이vvIBDV
에서는각각alanine
과serine
으로대체되며이아미노산변이가VP2
와
VP4
의구조에변화를주어병원성변화가가능할것으로추정된바있다
[29]. KNU08010
주도이부위에서proline
이각각alanine
과serine
으로변이가되어있는것 으로나타났다.
KNU08010
주를포함하여vvIBDV
인SH92
와OKYM
주모두
VP2-VP4
절단부위근처에서L451
특성을가지는것으로나타났다
.
단백질절단부위의아미노산변화는감염세포에서바이러스증식성변화로병원성변 이가가능할수있기때문에
[29], KNU08010
주에서나타나는이러한특징도이바이러스의병원성과관련이 있을것으로추정되었다
.
HVR
의염기서열을기준으로phylogenetic tree
를비교한결과에서 분리주
KNU08010
주는기존의classical IBDV
와는 다른계열로 분류되며국내외에서 보고된vvIBDV
와동일한계열로분류되는것으로나타나분리주
KNU08010
주는vvIBDV
인것으로판단되었다.
국내 분리vvIBDV
간에polyprotein
전체의아미노산을비교 한결과KNU08010
주는SH92
주와HVR
부위에서매우유사하였을뿐만아니라다른부위에서도상동성이매우 높아
segment A
유전자중4
개의아미노산차이만있었 다.
반면KK1
주와는HVR
에서는 유사성이 높았으나VP4
나VP3
등 다른 부위에서는 많은 차이를 보여segment A
전체유전자에서총17
개의아미노산차이를보이는것으로나타나분리주
KNU08010
주는KK1
주보다는
SH92
주와더유사한것으로생각되었다.
KNU08010
주는HVR
부위의phylogenetic tree
분석결 과최근야생조류에서분리된바이러스[16]
와매우가까운근연관계를보이는것으로나타났다
. IBDV
는닭 뿐만아니라항체조사결과야생조류에서도일부양성으 로나타나야생조류도IBDV
감염이가능한것으로보고된바있으며
[24, 27],
국내에서는야생조류들로부터IBDV
가분리된바있다[16].
따라서닭농장에서발생한
KNU08100
주와국내야생조류에서분리된바이러스와
HVR
유전자에서매우근연관계가높은것으로보아이들질병전파와야생조류의관련성도배제할수없을 것으로생각되나관련성을보다정확히구명하기위해 서는
HVR
이외에도polyprotein
단백질전체뿐만아니라
segment B
유전자의비교도추가로더필요할것으로생각되었다
.
결 론
2
주령전후의닭에서높은치사율을보이는사례에서IBDV
를분리하여분절A
유전자를분석하였다.
분리주는일본및한국에서분리보고된
vvIBDV
와매우높은아미노산상동성을보였으며
HVR
부위에서vvIBDV
간에잘보존되어있는것으로알려진
5
개의아미노산이모두존재하는것으로보아
vvIBDV
로분류되었다.
또한
HVR
의염기서열을기준으로phylogenetic tree
를작 성하여다른바이러스들과근연관계를분석한결과분리주
KNU08010
주는국내외보고된vvIBDV
와동일한 계열로분류되었고,
특히야생조류에서분리된vvIBDV
들과매우가까운근연관계를나타냈다
.
감사의 글
본 연구는 과학재단 특정기초연구사업
(
과제번호: 2010-0000138)
의지원에의해이루어진것으로,
지원에감사드립니다
.
참고문헌
1.
권용국,
모인필,
성환우,
강문일,
고홍범,
이정길,
양창근