서 론
해수 중으로 유입된 중금속은 수중에 유리된 상태로 존재하 거나 저질에 흡착되어 오랜 기간 잔류하게 되고, 여러 가지 경 로를 통해 각종 생물에 영향을 미친다(Arai et al., 2002). 일반 적으로 수중에 존재하는 구리(Cu)는 노출 시간과 노출 농도에 비례하여 갑각류 및 어류 등의 사망, 성장 및 생화학적 변동에 영향을 미친다(Marr et al., 1996; Wong et al., 1999; Mcgeer et al., 2000; Sherba et al., 2000; Flik et al., 2002; Minghetti et al., 2008).
복족류의 순환계는 개방혈관계로 혈구세포가 모든 조직 사 이를 순환하면서 상처치유, 패각의 재생, 영양분의 소화, 분비 물의 배설 등 다양한 기능을 수행하며, 이물질 침입에 대한 생 체방어의 기능이 있다(Cheng, 1981). Hemolymph 내의 혈구 들은 phagocytosis를 포함하여 다양한 형태의 면역 기능이 있 으며, 중금속과 같은 오염 물질에 의해 영향을 받으며(Exon and Koller, 1986), 체내에 흡수된 금속이온은 hemolymph 의 기능적인 변화를 야기시킨다(Weifang et al., 2009). 한편 hemolymph내의 phenoloxidase는 Cu를 함유하는 산화효소의 하나로 병원성 미생물 및 이물질이 체내로 들어왔을 때 이에 필 요한 멜라닌의 합성과 상처 부위의 치유에 중요한 역할을 담당
하며(Johnson et al., 2003), 생물체내에 구리와 같은 외인성 물 질이 다량으로 노출될 경우 방어반응으로 superoxide anion, hydrogen peroxide와 같은 활성산소(reactive oxygen species, ROS)를 제거하고 체외로 배설하기 위해 생체 변환을 일으킨 다(Barry et al., 1995). ROS에 대한 방어기작으로 작용하는 항 산화효소에는 superoxide dismutase (SOD), catalase (CAT), gultathione peroxidase, gultation S-transferase 등이 있으며 이들은 세포내에 여러 소기관, 세포질 및 핵 등 다양한 곳에 존재한다(Lopes et al., 2001). SOD는 두 가지의 이성질 효소 (Isoenzyme)를 가지는 항산화효소로서 free radical을 H2O2와 O2로 아세틸화시킨다. 또한 다른 항산화효소와 같이 작용하여
radical의 직접적인 독성과 이들이 금속이온과 상호작용하여
유독성의 hydroxy radical이 증가하는 것을 방지하는 역할을 하며, 이러한 과정에서 체내의 농도가 증가될 수 있다(Winston and Guilio, 1991; Parihar et al., 1997). CAT는 2H2O2를 2H2O 와 O2로 환원하는 효소로서 과산화물이 세포막에 활성이나 효 소활성을 억제하는 것을 막는 역할을 한다(Chance et al., 1979;
Wendel and Feuerstein, 1981).
따라서 본 연구는 연안해역에서 구리농도의 증가에 따라(Lee and Kim, 2000) 전복에 미치는 영향을 파악하기 위하여 hemo- lymph내의 무기, 유기 및 효소활성, hepatopancreas의 항산화 효소의 변화를 검토하였다.
*Corresponding author: [email protected]
154
구리(Cu) 노출에 따른 전복(Haliotis discus hannai)의 hemolymph 및 hepatopancreas 내의 생화학적 변화
박희주 . 강주찬 *
Biochemical Changes in the Hemolymph and Hepatopancreas of Abalone Haliotis discus hannai Exposed to Copper
Department of Aquatic Life Medicine, Pukyong National University, Busan 608-737, Korea
This study the effects of Cu exposure on biochemical factors in the hemolymph and hepatopancreas of the abalone Haliotis discus hannai. Abalone were exposed to 0, 5, 10, 20 and 40 µg/L Cu for 4 weeks. The calcium concentrations in hemolymph were decreased significantly on exposure to 20 and 40 µg/L Cu after 2 weeks. The aspartate aminotrans- ferase (AST), alanine aminotransferase (ALT) and alkaline phosphatase (ALP) activities in abalone hemolymph were markedly elevated after exposure to 40 µg/L Cu for 4 weeks. The hepatopancreas superoxide dismutase (SOD) and catalase (CAT) activities were also significantly increased by exposure to 20 µg/L Cu for 4 weeks. These biochemical factors may represent a convenient method of monitoring heavy metal pollution in coastal areas. From these results, we conclude that a high copper concentration (40 µg/L) in water may curtail hemolymph homeostasis and anti-oxidative reactions in abalone.
Key words : Cu, Abalone, Hemolymph, Hepatopancreas
Hee Ju Park and Ju Chan Kang*
부경대학교 수산생명의학과
재료 및 방법 실험동물
본 실험에 사용한 전복(Haliotis discus hannai)은 제주도 소 재의 양식장으로부터 분양받아 PVC(52×36×30 cm) 수조에 서 10일간 순치하였고, 시험에는 외관상 건강한 개체(각장 50.44±3.0 mm, 전중 14.74±2.5 g)를 사용하였다.
실험조건
모든 실험은 항온실(20±1℃)에서 순치 시에 사용한 수조에서 동일공간을 사용하여 실시하였으며, 매주 구간별로 6미의 전 복을 분석에 사용하였다. 실험에 사용한 해수의 수질은 Table 1 과 같다. 시험용액은 구리표준용액(copper standard solution) 1,000 mg/L을 이용하여 예비실험을 바탕으로 5, 10, 20, 및 40 µg/L로 설정하였다. 실험은 환수식 방법에 의해 매일 새로운 용액으로 교환하였고, 수질분석을 실시하였다. 먹이는 매일 미역(Undaria pinnatifida)을 충분한 양으로 공급하며 4주 동 안 실험을 실시하였다. 생화학적 분석은 시험시작 후 2주 및 4 주에 실시하였다.
Hemolymph 분석
Hemolymph는 hemocyte의 엉김을 방지하기 위해 EDTA (50 mM EDTA in PBS, pH 7.6)를 도포된 주사기로 심장 옆의 혈관에서 채취 한 뒤, 1,200×g에서 10분간 원심분리 후 상층액 에서 채취하여 -75℃에 보관하였다. Phenoloxidase의 활성은 Ashida and Soderhall(1984)을 이용하였다. 0.2M Tris-HCl buffer (pH 7.1)에 hemolymph와 10% sodium dodecyl sulfate 를 넣고 잘 혼합한 뒤, 15 mM L-DOPA (10 mM CaCl2 in 9,5 M HCl)를 넣은 뒤 즉시 파장 490 nm에서 흡광도를 측정하 였다. 흡광도 측정 후 28℃ 배양기에서 20분간 반응 시킨 후, 차가운 증류수를 넣어 반응을 정지시켰다. Hemolymph의 무
기성분은 마그네슘(Mg)과 칼슘(Ca)을 측정하였다. Mg의 정 량은 Xylidyl blue method, Ca은 OCPC (o-cresolphthalein- complexon)법으로 비색 정량하였다. 유기성분은 총 단백질 (total protein)과 glucose를 측정하였다. 총 단백질은 Biuret 법, glucose는 GOD법으로 측정하였다. 효소는 aspartate aminotransferase (AST), alanine amiotransferase (ALT) 및 alkaline phosphatase (ALP)를 분석하였다. AST와 ALT는 Retiman-Frankel법, ALP는 Kind-king법을 이용하여 측정하 였다. 각 측정에는 임상용 키트(Asan Pham. Co., Korea)를 사 용하였다.
Hepatopancreas 분석
항산화효소는 간췌장을 적출하여 SOD (superoxide dis- mutase) 및 CAT (catalase)를 각각 분석하였다. 간췌장 부위 를 적출 한 후 약 0.1g을 0.9% NaCl에 3회 세척한 다음 KCl (1.17%)을 함유한 100 mM phosphate buffer (pH 7.4)를 첨가 하여 teflon-glass homogenizer (099C K4424, Glas-Col)로 균 질화하였다. 균질화 된 시료는 1,000 rpm, 4℃, 15 min의 조건 에서 원심 분리하여(Mikro 22R, Hettich zentrifugen Ltd, Ger- many), 지방 및 침전물을 제거하였다. 균질액은 다시 13,000 rpm, 4℃, 20 min의 조건에서 원심분리 한 후 상층액을 -75℃ 에 보관하면서 항산화 효소 측정용 시료로 사용하였다.
SOD는 SOD Assay Kit-WST (Dojindo Co., Japan)를 이 용하였다. 분리 보관중인 간췌장 상층액을 5의 배수로 희석하 여 각 희석배수에 따른 inhibition rate를 구한 다음 spectropho- tometer를 이용하여 450 nm에서 inhibition curve를 작성하고, inhibition curve에서 활성이 50% 억제되는 농도를 찾아 계산 하였다. CAT활성도 측정은 H2O2를 기질로 사용하여 spectro- photometer에 의해 240 nm 파장에서 H2O2가 환원되어 감소하 는 흡광도로서 효소 활성도의 단위는 1분간에 1 mg의 단백질 이 반응하여 환원시킨 H2O2를 nmol로 나타내었다. 단백질 함 량은 Bradford법을 이용한 Kit (Biorad. Co. Ltd., USA)를 사 용했으며, 표준 단백질로서 BSA (bovine serum albumin)를 사용하여 spectrophotometer에서 540 nm로 측정하였다.
유의성 검정
실험 결과의 통계 처리는 SPSS 통계프로그램(version 12.0k) 을 이용하여 ANOVA test를 실시한 후, 사후 검정은 다중 비교 로 Duncan test를 사용하여 각 처리구 사이에 유의성(P<0.05) 을 조사하였다(Zar, 1996).
결 과 Hemolymph의 생화학적 변동
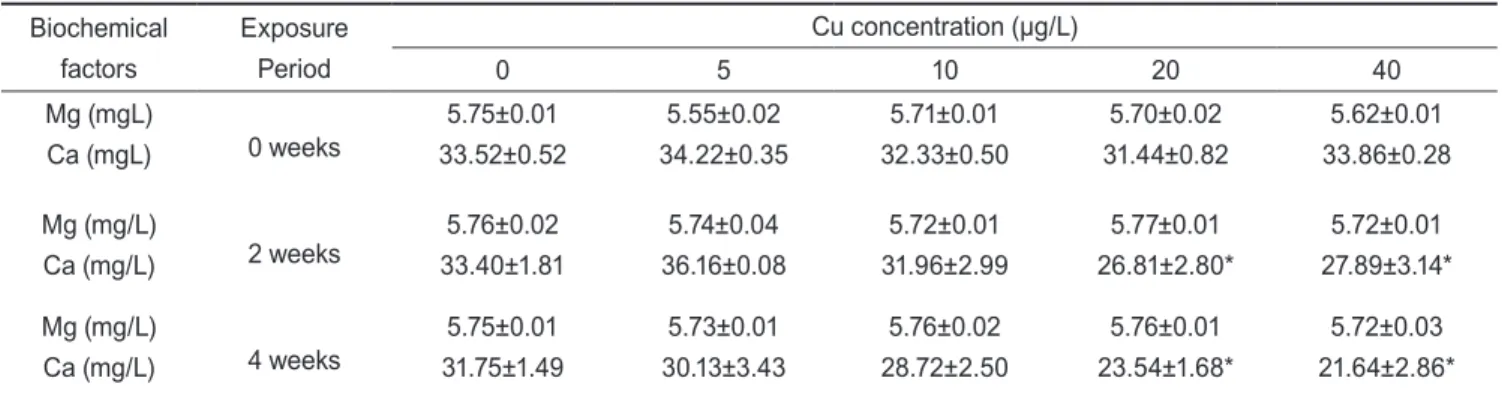
Cu에 노출된 전복의 hemolymph의 무기성분인 마그네슘 Table 1. The chemical components of seawater used in the
experiments
Item Value
Temperature (℃) 20.0±1.0
pH 8.1±0.2
Salinity (psu) 33.5±0.3
Dissolved oxygen (mg/L) 7.1± 0.2 Chemical oxygen demand (mg/L) 1.2± 0.1
Ammonia (µg/L) 9.5±0.7
Nitrite (µg/L) 5.3±0.3
Nitrate (µg/L) 14.4±1.0
Cu (µg/L) 0.2±0.14
(Mg)과 칼슘(Ca) 농도의 변화는 Table 2에 나타냈다. Mg농도 는 전 실험기간 중에 노출농도에 따라 유의한 차이를 나타내지 않았으나, Ca농도는 2주 및 4주째에 20 µg/L이상의 농도에서 유의적인 감소를 나타냈다(P<0.05). Hemolymph의 유기성분 인 총 단백질과 glucose 농도의 변화는 Table 3에 나타냈다.
Cu노출에 따른 총 단백질과 glucose농도는 실험기간동안 노출 농도에 따라 약간의 변동은 있었으나 유의한 차이는 나타나지 않았다. Hemolymp의 aspartate aminotransferase (AST), ala- nine aminotransferase (ALT) 및 alkaline phosphatase (ALP) 활성의 변화는 Figs. 1-3에 나타냈다. AST와 ALT활성은 노출 4주째에 40 µg/L에서 유의한 증가를 보였으며(P<0.05), ALP 활성은 노출 4주째에 20 µg/L이상의 농도에서 유의한 증가를 나타냈다(P<0.05).
Hepatopancreas의 산화효소 활성
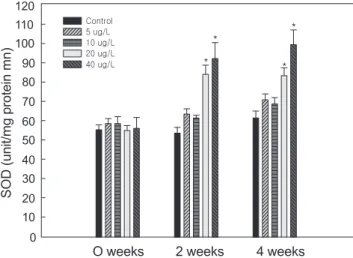
Cu에 노출된 전복의 hepatopancreas의 superoxide dis- mutase (SOD) 활성변화는 Fig. 4에 나타냈다. SOD활성은 Cu노출에 따라 증가하는 경향을 나타냈으며, 2주째부터는 20
Table 2. Changes of inorganic matter in hemolymph of abalone Haliotis discus hannai exposed to various Cu concentra- tions for 4 weeks
Biochemical factors
Exposure Period
Cu concentration (µg/L)
0 5 10 20 40
Mg (mgL)
Ca (mgL) 0 weeks 5.75±0.01
33.52±0.52
5.55±0.02 34.22±0.35
5.71±0.01 32.33±0.50
5.70±0.02 31.44±0.82
5.62±0.01 33.86±0.28 Mg (mg/L)
Ca (mg/L) 2 weeks 5.76±0.02
33.40±1.81
5.74±0.04 36.16±0.08
5.72±0.01 31.96±2.99
5.77±0.01 26.81±2.80*
5.72±0.01 27.89±3.14*
Mg (mg/L)
Ca (mg/L) 4 weeks 5.75±0.01
31.75±1.49
5.73±0.01 30.13±3.43
5.76±0.02 28.72±2.50
5.76±0.01 23.54±1.68*
5.72±0.03 21.64±2.86*
Values are mean±S.E. (n=7). *Values with superscript are significantly different (P<0.05) as determined by Duncan's test.
Fig. 1. Changes of aspartate aminotransferase (AST) activity in hemolymph of abalone Haliotis discus hannai exposed to various Cu concentrations for 4 weeks. Values are mean±S.E.
(n=7). *Values with superscript are significantly different (P<0.05) as determined by Duncan's test.
Table 3. Changes of organic matter in hemolymph of abalone Haliotis discus hannai exposed to various Cu concentrations for 4 weeks
Biochemical factors
Exposure Period
Cu concentration (µg/L)
0 5 10 20 40
TP (g/dL)
GL (g/dL) 0 weeks 0.720±0.030 15.47±0.30
0.710±0.060 15.31±0.80
0.714±0.054 15.40±0.57
0.729±0.062 15.50±0.51
0.718±0.077 15.51±0.72 TP (g/dL)
GL (g/dL) 2 weeks 0.725±0.049 15.08±0.73
0.714±0.017 15.40±0.97
0.695±0.016 15.42±0.64
0.698±0.024 15.59±1.01
0.720±0.016 15.40±0.32 TP (g/dL)
GL (g/dL) 4 weeks 0.725±0.010 15.97±0.20
0.715±0.016 15.51±0.37
0.712±0.030 15.46±0.83
0.709±0.025 14.40±1.13
0.689±0.011 14.13±0.19 Values are mean±S.E. (n=7). *Values with superscript are significantly different (P<0.05) as determined by Duncan's test.
0weeks 2weeks 4weeks
AST (Karmen Unit)
0 10 20 30 40 50 60 70
Control 5 ug/L 10 ug/L 20 ug/L 40 ug/L
* 60
50 40 30 20 10 0
AST (Karmen Unit)
Oweeks 2weeks 4weeks
고 찰
Cu와 같은 미량 금속은 연체동물의 체내에 흡수되어 정상적 인 생명활동의 유지와 생리적 반응의 항상성 유지에 이용되기 도 하지만, 필요 이상의 농도에 노출될 경우 세포 내부의 구조 적 이상을 유발하거나 생체 기능의 장애를 일으켜 생리적 활동 에 부정적인 영향을 미치기도 한다(Kawai, 1959; Sutherland µg/L이상의 농도에서 유의하게 증가하여 4주후에 40 µg/L농
도에서 최대 활성을 나타냈다(P<0.05). Hepatopancreas의 catalase (CAT)의 활성변화는 Fig. 5에 나타냈다. CAT활성은 Cu의 노출기간과 노출 농도에 따라 보편적으로 상승하는 경향 이 나타났으나, 유의한 활성의 증가는 2주 4주째에 40 µg/L 농 도에서 확인되었다(P<0.05).
Fig. 5. Changes of catalase (CAT) activity in hepatopancreas of abalone Haliotis discus hannai exposed to various Cu con- centrations for 4 weeks. Values are mean±S.E. (n=7). *Values with superscript are significantly different (P<0.05) as deter- mined by Duncan’s test.
0 weeks 2 weeks 4 weeks
CAT ( nmol/mg protein sec)
0.0 0.5 1.0 1.5 2.0 2.5 3.0
Control 5 ug/L 10 ug/L 20 ug/L 40 ug/L
* *
CAT ( nmol/mg protein sec)
3.0 2.5 2.0 1.5 1.0 0.5
0.0 O weeks 2 weeks 4 weeks
ATL (Karmen Unit)
0 weeks 2 weeks 4 weeks
ALT (Karmen Unit)
0 10 20 30 40 50
Control 5 ug/L 10 ug/L 20 ug/L 40 ug/L
*
Fig. 2. Changes of aline aminotransferase (ALT) activity in hemolymph of abalone Haliotis discus hannai exposed to various Cu concentrations for 4 weeks. Values are mean±S.E.
(n=7). *Values with superscript are significantly different (P<0.05) as determined by Duncan's test.
50 40 30 20 10
0 O weeks 2 weeks 4 weeks
Fig. 4. Changes of superoxide dismutase (SOD) activity in hepatopancreas of abalone Haliotis discus hannai exposed to various Cu concentrations for 4 weeks. Values are mean±S.E.
(n=7). *Values with superscript are significantly different (P<0.05) as determined by Duncan’s test.
O weeks0 weeks 2 weeks2 weeks 4 weeks4 weeks
SOD (unit/mg protein mn)
0 10 20 30 40 50 60 70 80 90 100 110 120
Control 5 ug/L 10 ug/L 20 ug/L 40 ug/L
*
*
* *
SOD (unit/mg protein mn)
120 110 100 90 80 70 60 50 40 30 20 10 0
Fig. 3. Changes of alkaline phosphatase (ALP) activity in hemolymph of abalone Haliotis discus hannai exposed to various Cu concentrations for 4 weeks. Values are mean±S.E.
(n=7). *Values with superscript are significantly different (P<0.05) as determined by Duncan's test.
0 weeks 2 weeks 4 weeks
ALP (Unit/L)
0.0 0.5 1.0 1.5 2.0 2.5 3.0
Control 5 ug/L 10 ug/L 20 ug/L 40 ug/L
* *
ALP (Unit/L)
3.0 2.5 2.0 1.5 1.0 0.5
0.0 O weeks 2 weeks 4 weeks
and major, 1981). 무척추동물의 개방혈관계는 선천성 면역체 계가 주요하며, pro-phenoloxidase를 매개로 하는 멜라닌 자극 계는 방어 수단으로 역할을 수행한다. Hemolymph 내에서 합 성된 pro-phenoloxidase는 체내의 세균 및 기생충과 같은 미생 물이나 기타 이물질에 의해 phenoloxidase로 활성화되며, 방 어 작용을 담당하는 phenoloxidase는 금속이나 유기용매에 의 해 감소한다고 알려져 있다. Cu, 아연, 칼슘과 같은 2가 양이 온을 형성하는 금속은 바지락 내의 sulpher기와 결합하여 phe- noloxidase 활성을 저해시키며(Rishan et al., 2005), 새우류에 있어 Cu는 phenoloxidase활성이 감소된다(Cheng and Wang, 2001). Cu에 노출된 전복의 phenoloxidase의 활성은 유의한 차이는 없었으나 노출농도가 증감함에 따라 감소하는 경향을 나타내어 전복에 있어서도 Cu는 phenoloxidase의 활성을 감소 시키는 인자로 작용할 수 있을 것으로 예상된다.
어류에 있어서의 혈액학적 지표들은 다양한 스트레스에 대해 반응한다(Jee et al., 2004). 전복의 hemolymph의 무기성분인 마그네슘(Mg)에서는 모든 실험구간에서 유의적인 변화가 관 찰되지 않았으며, 칼슘(Ca)에서는 20 µg/L의 농도에서부터 유 의적인 감소가 나타났다. 일반적으로 중금속에 노출된 어류에 서 상피세포의 막 투과성 변화 및 장관의 흡수 장애로 인한 칼 슘 농도의 감소가 보고되었다(Koyama and Itazawa, 1977). 그리고 Cu의 아가미 축적은 아가미의 세포막의 Ca2+-APTase 의 효소활성을 억제하는 것으로 보고되었다(Viarengo et al., 1996). 따라서 전복에 있어서도 다량의 Cu에 의해 칼슘대사가 저해된 것으로 판단된다. 결합조직을 구성하는 Hemolymph 의 유기성분인 총 단백질 및 glucose 농도는 일반적으로 대 사 과정에서 필수적인 요소로서 스트레스에 의한 체내의 방 어 작용에서 사용되어 지기도 한다(Abdel-Moati, 1992). 전 복의 hemolymph의 총 단백질과 glucose농도의 유의한 변화 는 관찰되지 않았지만, 고농도의 노출에서는 감소하는 경향 을 나타냈다. 이는 다양한 방어 작용을 일으키는 과정에서 발 생한 총 단백질 및 glucose 농도의 소모가 하나의 원인으로 생 각할 수 있다. AST와 ALT는 해당 효소를 이루는 필수아미노 산과 탄수화물 대사에도 중요한 역할을 한다. 평상시에는 혈중 에 미량이 존재하고 있으나, 간이나 췌장 등 장기의 조직적 손 상이나 이상발생시 혈중으로의 유리되어 증가하는 것으로 알 려져 있다(Blasco and Puppo,1999). 전복의 AST와 ALT는 노 출 4주째에 40 µg/L의 Cu농도에서 유의적인 증가를 나타내 어 Blasco and Puppo (1999)의 연구에서 보고된 바지락, R.
philippinarum의 결과와 유사한 결과를 나타내고 있다. ALP 는 금속이온에 민감하게 반응하여 금속 중독의 척도로 이용되 고 있다(Bogé et al., 1988). 전복의 ALP 활성은 Cu 20 µg/L 이상의 농도에서 4주째에 유의한 증가가 확인되어 이 같은 결 과는 앞의 두 효소와 마찬가지로 Cu의 영향에 대한 생체의 방 어 기작으로 생각된다.
척추동물이 가지는 간의 기능을 하는 것으로 알려진 무척추동
물의 간췌장(hepatopancreas)은 다양한 효소작용의 근원이 된 다(George and Norman, 1973). 다양한 원인에 의해 발생되는 활성산소는 노화의 원인이기도 하며, 조직의 생리적 반응을 저 해하기도 한다(Goldberg and Stern, 1977; Simon et al., 1981;
Moody and Hassan, 1982). 과다한 Cu 스트레스로 인해 발생 된 활성산소는 SOD에 의해 과산화수소로 변환되어 해독화가 진행된다(Nebot et al., 1993). Cu에 노출된 담치(M. edulis)는 O2-가 증가되며(Pipe et al., 1999), 전복의 SOD는 수온 자극 과 같은 외부 스트레스에 의해서도 급격하게 활성이 증가된다 (Kim et al. 2005). 전복의 SOD활성은 2주째부터 20 µg/L이 상의 Cu농도에서 유의한 증가가 관찰되었다. 이 같은 결과는 Cu 노출에 의한 조직 손상을 최소화 하려는 전복의 방어행동에 의해 진행되는 생리적작용으로 추정된다. SOD와 더불어 CAT 의 활성은 SOD에 의해 전환된 과산화수소(H2O2)를 다시 산소 (O2)와 물(H2O)로 전환시킨다(Forman and Fridovich, 1973). 본 실험에서 역시 SOD의 활성이 증가하면서 발생된 과산화수 소를 CAT가 분해하는 모습을 보여주고 있다. SOD의 활성 이 후에 천천히 CAT의 활성이 증가하고 있으며, Cu 20 µg/L이상 2주째부터 유의성은 없으나 증가폭이 커짐을 확인할 수 있었 다. 이는 SOD에서 CAT로 이어지는 단계적인 산화스트레스의 저하 시스템으로 가정할 수 있다.
이상의 결과와 논의로부터 구리에 대한 전복의 hemolymph 내의 생리적 변동 및 hepatopancreas의 항산화효소의 활성의 변동은 연안 중금속 오염에 대한 모니터링의 수단으로 이용될 수 있을 것으로 생각된다.
사 사
본 연구는 국립수산과학원 R&D과제‘생태독성 평가기법을 이용한 어장 건강성 평가 연구’의 지원에 의해 수행되었습니 다.
참고문헌
Abdel-Moati M. 1992. Biochemical and physiologycal responses of Mytilus edulis to Hg and Pb in the coastal waters of Alex- andria region. Proceeding of the FAD-UNEP-IOC workshop on biological effects of pollutants in marine organisms. Ath- ens, Greece 69, 61-62.
Arai T, Maeda M, Yamakawa H, Kamatani A and Miyazaki N.
2002. Growth effect on the uptake and elimination of trace metals in the abalones Haliotis. Fisheries Sci 68, 1094-1098.
Ashida M, Soderhall K. 1984. The prophenoloxidase activating system in crayfish. Comp Biochem Physiol 77, B21-6.
Barry H and John MCG. 1995. The definition and measurement of antioxidants in biological systems Free Radical Biology and Medicine, 125-126.
Blasco J and Puppo J. 1999. Effect of heavy metals (Cu, Cd and Pb) on aspartate and alanine aminotransferase in Ruditapes
philippinarum (Mollusca: Bivalvia) Comparative Biochem- istry and Physiology Part C: Pharmacology, Toxicology and Endocrinology, 253-263.
Bogé G, Bussiere D and Pérès G. 1988. Effets du chrome hexava- lent sur les fonctions de la bordure en brosse de l'intestin de la truite arc en ciel (Salmo gairdneri R.) Water Research, 441- 447.
Chance B, Sice H and Boveris A. 1979. Hydroperoxide metabo- lism in mamamalian organs. Physiil Rev, 527-605.
Cheng TC. 1981. Bivalves. In: Inveterbrate Blood Cell. Ratcliffe NA and Rowley AF, eds. Academic Press. London, 233-300.
Cheng W and Wang CH. 2001. The susceptibility of the giant freshwater prawn Macrobrachium rosenbergii to Lactococ- cus parvieae and its resistance under copper sulfate stress.
Dis Aquat Organ 47, 137-144.
Exon JH and Koller LD. 1986. Immunotoxicity for cadmium: In:
Handbook of Experimental Pharmacology. Cadmium. Foul- kes EC, ed. Berlin Heidelberg, Springer-Verlag. 339-350.
Forman HJ and Fridovich I. 1973. Superoxide dismutase: A com- parison of rate constant. Arch Biochem Biophys 158, 396.
George H and Norman L. 1973. Enzymatic activity in hepato- pancreas of Nassarius obsoletus. Comp Biochem Phys B, 153-157.
Goldberg B. and Stern A. 1977. The role of the superoxide anion as a toxic species in the erythrocyte. Arch Biochem Biophys, 218-225.
Johnson JK, Rocheleau TA, Hillyer JF, Chen CC, Li J and Chris- tensen BM. 2003. A otential role for phenylalanine hydroxy- lase in mosquito immune responses. Inspect. Biochem Mol Biol 33, 345-354.
Jee JH, Kim SG and Kang JC. 2004. Effects of phenanthrene on growth and basic physiological functions of the olive floun- der, Paralichthys olivaceus. J Exp Mar Biol Ecol 304, 123- Kawai K. 1959. The cytochrome system in marine lamellibranch 136.
tissues. Biol Bull 117, 125-132.
Kim JH, Mok JS and Park HY. 2005. Trace Metal Contents in Seaweeds from Korean Coastal Area. J. Korean Soc Food Sci Nutr 34, 1041-1051.
Koyama J. and Itazawa Y. 1977. Effects of oral administration of cadmuim on fish: I. Analutical results of the blood and bones.
Bull Jap Soc Sci Fish 43, 523-526.
Lee IS and Kim EJ. 2000. Distribution of Heavy Metals in Sedi- ments , Seawater and Oysters (Crassostrea gigas) in the Jin- hae Bay. J Ecology 23, 59-64.
Lopes PA, Pinheiro T, Mathias MC, Collares-Pereira MJ and Vie- gas-Crespo, AM. 2001. Response of antioxidant enzymee in freshwater fish population (Leuciscus alburnoides complex) to inorganic pollutants exposure. Sci Total Environ 280, 153- 163.
Marr JCA, Lipton J, Cacela D, Hansen JA, Bergman HL, Meyer JS. and Hogstrand C. 1996. Realtionships between copper exposure duration, tissue copper concentration, and rainbow
trout growth. Aquat Toxicol 36, 17-30.
Minghetti M, Leavera MJ, Carpeneb E and Georgea SG. 2008.
Copper transporter 1, metallothionein and glutathione reduc- tase genes are differentially expressed in tissues of sea bream (Sparus aurata) after exposure to dietary or waterborne cop- per. Toxicol Appl Pharm Volume 147 Issue 4, 450-459.
Mcgeer JC, Szebedinszky C, Mcdonaldm DG and Wood CM.
2000. Effects of chronic sublethal exposure to waterborne Cu, Cd or Zn in rainbow trout. 1: Iono-regulatory disturbance and metabolic costs. Aquat Toxicol 50, 231-243.
Moody CS and Hassan HM. 1982. Mutagenicity of oxygen free radicals. Proc Natl Acad SCI 79, 2855-2859.
Nebot C, Moutet M, Huet P, Xu JZ, Yadan JC. and Chaudiere J. 1993. Spectophotometric assay of superoxide dismutase activity basedon the activited autoxidation of a tetracyclic catechol. Anal Biochem 214, 442-451.
Parihar MS, Javeri T, Hemnani T, Dubey AK and Parkash P.
1997. Response of superoxide dismutase, glutathion peroxi- dase and reduced glutathion antioxidant defenses in gills of the freshwater catfish (Heteropneustes fossilis) to short-term elevated temperature. J Therm Biol 22, 151-156.
Pipe RK, Coles JA, Carissan FMM and Ramanathan K. 1999.
Copper induced immunomodulation in the marine mussel, Mytilus edulis. Aquqt Toxicol 46, 43-54.
Rishan CS, Wenjie L, Guangxing F, Tingjun, M. xianghong, Lin- gling Y and Liyan Z. 2005. Purification and characterization of phenoloxidase from clam, Ruditapes philippinarum. Fish Shell Immuno 18, 61-70.
Sherba M, Dunham DW and Harvey HH. 2000. Sublethal copper toxicity and food response in the freshwater crayfish, Cam- barus bartonii (Cambaridae, Decapoda, Crustacea). Ecotox- icol Environ Saf 46, 329-333.
Simon RH, Scoggin CH and Patterson D. 1981. Hydrogen per- oxide causes the fatal injury to human fibroblasts exposed to oxygen radicals. J Biol Chem 256, 7181-7186.
Sutherland J and Major CW. 1981. Internal heavy metal changes as a consequence of exposure of Mytilus edulis, the blue mussel, to elevated external copper (II) levels. Comp Bio- chem Physiol, 63-67.
Viarengo A, Pertic M, Mancinelli G, Burlundo B, Canesi JL and Orunesu M. 1996. In Vivo effects of copper on the calcium homeostasis mechanisms of mussel gill cell plasma mem- branes. Comp Biochem Phys B 113, 421-425.
Weifang W, Kangsen M, Wenbing Z, Qinghui A, Chunfeng Y, Huitao L and Zhiguo L. 2009. Effects of diertry copper on survival, growth and immune response of juvenile abalone, Haliotis discus hannai. Ino Aquaculture 297, 122-127.
Wendel A. and Feuerstein S. 1981. Drug-induced lipid peroxida- tion in mice-1. Modulation by monoxygenase activity, glu- tathione and selenium status. Biochem Pharmacol 30, 2513- 2520.
Winston GW and Di Giulio RT, 1991. Pro-oxidant and antioxi- dant mechanisms in aquatic organsm. Aquat Toxicol 19, 137-
161.
Wong PPK, Chu LM and Wong CK. 1999. Study of toxicity and bioaccumulation of copper in the silver sea bream Sparus sarba. Environ Int 25, 417-422.
Zar JH. 1996. Biostatistical Analysis. Prentice Hall, London, 662.
2011년 8월 12일 접수 2012년 1월 25일 수정 2012년 2월 6일 수리