p38 Mitogen-Activated Protein Kinase의 억제를 통한 진세노사이드 Rg3의 Thromboxane A
2억제 효과
권 혁 우 극동대학교 임상병리학과
Inhibitory Effects of 20(S)-Ginsenoside Rg3 on Thromboxane A 2
Generation via p38 Mitogen-Activated Protein Kinase
Hyuk-Woo Kwon
Department of Biomedical Laboratory Science, Far East University
ABSTRACT Thromboxane A
2plays a central role in hemostasis and thrombus formation, which stimulates other platelets as an agonist leading intracellular signaling cascades. In this study, we tried to explain the inhibitory mechanism of 20(S)-ginsenoside Rg3 (G-Rg3) on thromboxane A
2production by assaying the activities of TXA
2production-asso- ciated microsomal enzymes and associated signaling molecules. Thrombin-stimulated human platelet aggregation was dose-dependently decreased by G-Rg3 and attenuated thromboxane A
2production via down regulation of microsomal cyclooxygenase-1 and thromboxane A
2synthase activities. In addition, arachidonic acid release, which is a precursor of TXA
2was decreased by G-Rg3 through dephosphorylation of cytosolic phospholipase A
2and p38 mitogen-activated protein kinase. Overall, the results revealed the specific signaling pathway of TXA
2production via G-Rg3. Therefore, G-Rg3 is a beneficial novel compound inhibiting TXA
2production, which may prevent platelet aggregation-mediated thrombotic disease.
Key words: 20(S)-ginsenoside Rg3, thromboxane A
2, arachidonic acid, cyclooxygenase-1, thromboxane A
2synthase
Received 3 July 2018; Accepted 27 August 2018
Corresponding author: Hyuk-Woo Kwon, Department of Biomed- ical Laboratory Science, Far East University, Chungbuk 27601, Korea
E-mail: [email protected], Phone: +82-43-880-3081
서 론
혈액은 각 장기와 조직에 영양분 및 산소를 공급하고 노폐 물을 조직으로 운반하는 생존에 절대적인 기능을 담당한다.
이와 같은 기능을 수행하기 위하여 혈액은 항상 원활하게 순환되어야 한다. 따라서 혈관이 파열될 경우 손상된 부위로 인한 혈액 손실을 최소화하고 정상적인 순환을 유지하기 위 해서는 신속한 지혈 반응이 필요하다. 이처럼 혈소판 응집반 응은 지혈을 위한 필수적인 과정이지만, 동시에 고지혈증과 함께 혈전증, 동맥경화증, 뇌졸중, 심근경색, 죽상동맥경화 증과 같은 심혈관계 질환을 유발할 수 있는 요인이 되기도 한다. 그러므로 혈소판 응집을 저해시킬 수 있는 다양한 약 물의 탐색은 심혈관계 질환의 예방을 위한 중요한 요소라 할 수 있다(1,2). 혈관의 손상 부위에 동원된 혈소판은 다양 한 생체 agonists(collagen, adenosine diphosphate, throm- bin)에 의해 활성화되며, 활성화된 혈소판 막의 phosphati- dylinositol 4,5-bisphosphate는 phospholipase C에 의해
서 inositol-1,4,5-triphosphate(IP3)와 diacylglycerol로 가수분해되고, 생성된 IP3는 혈소판의 endothelial retic- ulum의 Ca2+ channel을 열어 세포질 내부의 Ca2+ 농도를 증가시킨다. 혈소판 내 증가한 Ca2+은 myosin light chain 을 인산화하여 혈소판의 shape change와 granule release 를 유도하고 최종적으로 혈소판의 응집반응을 일으킨다(3, 4). 이러한 혈소판의 활성화 과정 중 thromboxane A2 (TXA2)의 전구체인 arachidonic acid는 cytosolic phos- pholipase A2(cPLA2)에 의해 가수분해되어 세포질 내부로 유입되고 이후 합성효소인 cyclooxygenase-1(COX-1)과 TXA2 synthase(TXAS)에 의해 TXA2로 전환된 후 혈소판 외부로 분비된다(5,6). 이때 생성된 TXA2는 주변의 다른 혈 소판 막 수용체에 결합하여 혈소판 응집을 더욱 촉진하는 강력한 agonist로 작용한다고 알려져 있다(7).
Mitogen-activated protein kinases(MAPKs)는 세포의 신호전달계에 작용하는 인산화 효소로서 extracellular sig- nal-regulated kinase(ERK1/2), p38 MAPK, c-Jun N- terminal kinase(JNK)로 분류되며, 지혈과 혈전 생성에서 의 MAPKs의 역할은 오랫동안 연구되어 왔다(8). MAPKs 중 ERK2, JNK1과 p38은 인체 혈소판에서 검출되었으며 트롬빈에 의해 인산화되어 활성을 가지는 것으로 보고되었 다(9-11). cPLA2는 TXA2 생산을 위한 효소로서 트롬빈에
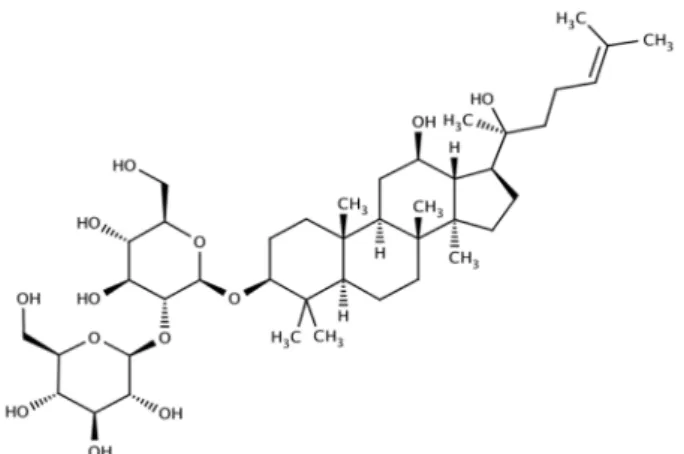
Fig. 1. Chemical structure of 20(S)-ginsenoside Rg3.
의한 자극 시 세포질 내 cPLA2는 세포 내 Ca2+의 존재 하에 혈소판 막으로 이동하여 arachidonic acid를 유리시킨다 (12). 이때 cPLA2의 완전한 촉매 활성을 위해 p38 MAPK는 cPLA2의 Ser505 위치를 인산화시킨다(13).
인삼(
Panax ginseng
Meyer)은 전통 동양 의학에서 다양 하게 사용되어 왔으며, ginsenoside Rg3(G-Rg3)는 colla- gen 유도 응집반응, TXA2 생성, ATP 방출 및 Ca2+의 동원 에 대한 억제 효과가 있는 것으로 보고되었다(14,15). 또한 Rg3의 유도체인 dihydroxyginsenoside Rg3는 ERK2와 p38 MAPK를 감소시켰고(16), ginsenoside-Rg3가 풍부 한 홍삼 추출물은 collagen으로 유발한 rat 혈소판 응집반응 에서 억제 효과를 보였다(17). 또한, 우리는 이전 실험에서 트롬빈에 의해 유도된 인체 혈소판 응집반응에서 G-Rg3 (20S, 20R)의 억제 효과를 확인하였지만(18), G-Rg3의 저 해 메커니즘은 완전히 설명되지 못했다. 따라서 본 연구에서 는 G-Rg3(20S)가 TXA2 생성에 관여한다고 알려진 효소인 COX-1, TXAS, cPLA2와 관련 신호전달 분자인 p38 MAPK 의 인산화를 어떻게 조절하며 혈소판 응집을 억제에 관여하 는지 규명하고자 하였다.재료 및 방법
재료
20(S)-Ginsenoside Rg3(G-Rg3, Fig. 1)는 엠보연구소 (Daejon, Korea)에서 구입하였다. G-Rg3는 Korean red ginseng에서 추출되었으며(순도 98% 이상) 분자량은 785.025 g/mol이다. Thrombin은 Chrono-Log 사(Ha- vertown, PA, USA)에서, prostaglandin H2, TXB2 en- zyme immunoassay(EIA) kits, Lactate dehydrogenase cytotoxicity assay kit과 COX fluorescent activity assay kit은 Cayman Chemical 사(Ann Arbor, MI, USA)로부터 구입하였다. 그 밖의 시약들은 Sigma-Aldrich 사(St. Louis, MO, USA)에서 구입하였고, Serotonin EIA kit은 Labor Diagnostika Nord 사(Nordhorn, Germany)에서, ATP assay kit은 Biomedical Research Service Center(Buf-
falo, NY, USA)에서 구입하였다. Western blotting용 anti- body들과 lysis buffer는 Cell Signaling(Beverly, MA, USA)에서 구입하였고, polyvinylidene difluoride(PVDF) membrane과 enhanced chemiluminescence solution(ECL) 은 GE Healthcare(Buckinghamshire, UK)에서 구입하였다.
세척 혈소판 준비
Acid-citrate-dextrose solution(0.8% citric acid, 2.2
% sodium citrate, 2.45% glucose)으로 항응고 처리된 hu- man platelet-rich plasma(PRP)를 한국적십자 혈액원 (Changwon, Korea)으로부터 제공받았다. 미량의 적혈구 를 제거하기 위해 PRP를 125×
g
에서 10분간 원심분리 한 후, 1,300×g
에서 10분간 원심분리 하여 platelet pellets을 얻었다. 이것을 washing buffer(138 mM NaCl, 2.7 mM KCl, 12 mM NaHCO3, 0.36 mM NaH2PO4, 5.5 mM glu- cose, 1 mM EDTA, pH 6.5)로 두 번 세척하고. 세척된 혈소 판을 suspension buffer(138 mM NaCl, 2.7 mM KCl, 12 mM NaHCO3, 0.36 mM NaH2PO4, 0.49 mM MgCl2, 5.5 mM glucose, 0.25% gelatin, pH 6.9)로 재구성하여 Born 과 Hume(19)의 방법을 사용하여 최종 108/mL 농도가 되게 하였다. 위에 있는 모든 과정은 낮은 온도에서 일어날 수 있는 혈소판 응집을 피하기 위하여 25°C에서 수행하였다.이 실험은 The Korea National Institute for Bioethics Policy Public Institutional Review Board(Seoul, Korea) 의 승인을 받아 수행되었다(PIRB12-072).
혈소판 응집반응 측정
혈소판 응집반응은 Born(20)의 방법에 따라 시행되었다.
세척 혈소판(108/mL)에 여러 농도의 G-Rg3(50~300 μM) 를 첨가하여 37°C에서 3분간 전처리한 후, 0.05 U/mL thrombin으로 응집을 유도하고 5분간 측정하였다. 응집은 1,000 rpm stirring speed에서 aggregometer로 측정하였 고(Chrono-Log, Havertown, PA, USA), 응집능은 빛 투과 도의 증가한 정도로 산출하였다. Suspension buffer를 투과 도 0%의 기준값으로 사용하였고, G-Rg3는 dimethyl sulf- oxide(DMSO)에 녹여 0.1%의 최종농도로 사용하였다.
세포독성 평가
세척 혈소판(108/mL)에 여러 농도의 G-Rg3(50~300 μM) 를 첨가하여 37°C에서 5분간 전처리한 후, 12,000×
g
로 15 분간 원심분리 하여 세포 debris를 제거한 상층을 lactate dehydrogenase cytotoxicity assay kit(Cayman Chemical 사)으로 측정하였다. 0.1% Triton X-100으로 혈소판을 완 전히 용해한 값은 양성대조군으로서 100%로 기준을 정하고 G-Rg3의 값을 %로 제시하였다.TXB
2생성량 측정
세척 혈소판(108/mL)에 여러 농도의 G-Rg3(50~300 μM)
를 첨가하여 37°C에서 3분간 전처리한 후, 0.05 U/mL thrombin으로 응집을 유도하고 5분간 반응시켰다. Ice- cold 1 mM EDTA와 0.2 mM indomethacin을 첨가하여 반응을 정지시켰다. TXA2의 안정한 대사체인 TXB2의 양을 TXB2 EIA kit을 사용하여 synergy HT multi-model mi- croplate reader(BioTek Instruments, Winooski, VT, USA)로 측정하였다.
COX-1의 활성 측정
COX-1과 TXAS의 활성 측정은 Lee 등(21,22)의 방법에 따라 시행되었다. 세척 혈소판(108/mL)에 1% protease in- hibitor cocktail(Sigma-Aldrich 사)을 첨가하여 sonicator (Bandelin Electronic, Bandelin, Germany)로 100% 감도 에서 20초 동안 10번 sonication 하여 혈소판 용해물을 만 들었다. 이를 4°C에서 12,000×
g
로 15분간 원심분리 하여 세포 debris를 제거한 후 상층을 4°C에서 105,000×g
로 1시 간 원심분리 하여 microsomal fraction을 분리하였다(23).분리된 microsomal fraction을 bicinchoninic acid protein assay kit(Pierce Biotechnology, IL, USA)을 사용하여 단 백질을 정량한 후 1 mg microsomal fraction에 G-Rg3(50
~300 μM)를 첨가하여 37°C에서 30분간 전처리한 다음, COX-1 활성을 COX activity assay kit을 사용하여 syn- ergy HT multi-model microplate reader(BioTek Instru- ments)로 측정하였다.
TXAS의 활성 측정
COX-1 활성 측정과 마찬가지로 microsomal fraction 1 mg에 G-Rg3(50~300 μM)를 첨가하여 37°C에서 30분간 전처리한 후 prostaglandin H2를 첨가하여 37°C에서 1분간 효소반응을 일으키고 1 M citric acid로 정지시켰다. 그 후 1 N NaOH를 사용하여 중화시킨 다음 TXA2의 안정한 대사 체인 TXB2의 양을 TXB2 EIA kit을 사용하여 synergy HT multi-model microplate reader(BioTek Instruments)로 측정하였다.
Arachidonic acid 방출의 측정
세척 혈소판(108/mL)에 여러 농도의 G-Rg3(50~300 μM) 를 첨가하여 37°C에서 3분간 전처리한 후, 0.05 U/mL thrombin으로 응집을 유도하고 5분간 측정하였다. Ice-cold 1 mM EDTA와 0.2 mM indomethacin을 첨가하여 반응을 정지시켰다. 생성된 arachidonic acid의 양을 EIA kit (Cusabio, Wuhan, Hubei, China)을 사용하여 synergy HT multi-model microplate reader(BioTek Instruments)로 측정하였다.
Western blot을 이용한 cPLA
2와 p38 MAPK의 인산화 측정
세척 혈소판(108/mL)에 여러 농도의 G-Rg3(50~300 μM) 를 첨가하여 37°C에서 3분간 전처리한 후 0.05 U/mLthrombin으로 응집을 유도하고 5분간 반응시켰다. 동량의 lysis buffer를 첨가함으로써 반응을 정지시켰다. 그 후 혈소 판 용해물을 bicinchoninic acid protein assay kit(Pierce Biotechnology, Rockford, IL, USA)을 사용하여 단백질을 정량하였고 동량의 단백질(15 μg)을 분석에 사용하였다. 전 기영동은 6~8% SDS-PAGE를 사용하였고 PVDF mem- brane에 단백질을 transfer 하였으며, transfer 한 mem- brane은 ECL 시약을 반응시켜 발색하였다.
ATP와 serotonin 방출의 측정
ATP와 serotonin의 방출은 Park 등(24)의 방법에 따라 수행되었다. 세척 혈소판(108/mL)에 여러 농도의 G-Rg3 (50~300 μM)를 첨가하여 37°C에서 3분간 전처리한 후, 0.05 U/mL thrombin으로 응집을 유도하고 5분간 반응시켰 다. Ice-cold 1 mM EDTA로 반응을 정지한 후, 원심분리 하여 상층으로 방출된 ATP와 serotonin을 ATP assay kit 과 serotonin EIA kit을 사용하여 Synergy HT multi- model microplate reader(BioTek Instruments)로 측정하 였다.
통계분석
측정된 모든 실험 결과는 mean±SEM으로 처리하여 analysis of variance(ANOVA)로 분석하였다. 그룹 간의 평균에 유의적인 차이가 있을 경우 Newman-Keuls meth- od로 비교하여 각 그룹 간에 표기하였다.
P
<0.05일 때 유의 적인 의미가 있는 것으로 판단하였다.결 과
G-Rg3가 thrombin 유도 혈소판 응집과 세포독성에 미치 는 효과
혈소판을 최대치로 응집시키는 thrombin의 농도가 약 0.05 U/mL였기에, 본 실험에서는 0.05 U/mL thrombin으 로 응집을 유도하였다(18). 혈소판에 thrombin을 첨가하여 응집을 유도하였을 때 응집률이 91.0±1.0%로 나타났고, G-Rg3를 다양한 농도(50~300 μM)로 첨가하였을 때에 농 도 의존적으로 강하게 응집이 억제되는 결과를 확인하였다 (Fig. 2A). 이때 0.1% DMSO는 thrombin 자극에 영향을 미치지 않았다. 이는 G-Rg3가 강력한 항 혈소판 효과를 가 지고 있는 물질임을 보여주는 결과이다. 천연물의 세포 독성 을 평가하기 위하여 lactate dehydrogenase(LDH) leak- age를 수행하였다. 인체 혈소판에 G-Rg3(50~300 μM)를 처리하여 LDH leakage를 분석한 결과 G-Rg3는 세포독성 결과에서 유의성을 나타내지 않았다(Fig. 2B). 0.1% Triton- X100은 양성대조군으로 사용되었다.
G-Rg3가 TXA
2방출에 미치는 효과
다음으로 G-Rg3가 TXA2의 방출에 미치는 영향을 확인
A
B
Fig. 2. Effects of G-Rg3 on thrombin-induced platelet aggrega-
tion and cytotoxicity. (A) Effect of G-Rg3 on thrombin-induced human platelet aggregation. (B) Effect of G-Rg3 on cytotoxicity.Platelet aggregation and cytotoxicity were carried out as described in ‘Materials and Methods’ section. The data are expressed as the mean±standard deviation (n=4). *
P<0.05,
**P<0.01 versus the
thrombin-stimulated human platelets. NS, not significant versus without G-Rg3.하였다. Fig. 3에서 보이는 바와 같이 intact cell에서 1.2±
0.2 ng/108 platelets였던 TXA2 양이 thrombin에 의해 47.3
±1.9 ng/108 platelets로 강하게 증가하였고, G-Rg3(300 μM)는 thrombin에 의해 증가한 TXA2를 5.2±1.7 ng/108 platelets로 89.0% 억제하였다.
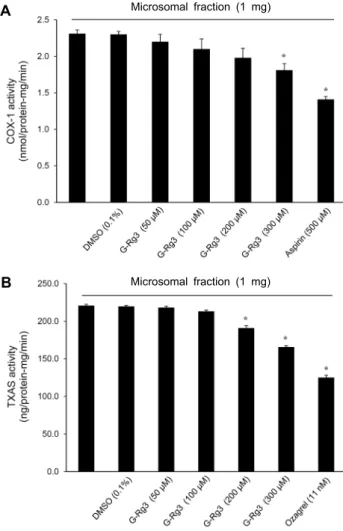
G-Rg3가 COX-1와 TXAS 활성에 미치는 효과
TXA2 생성은 혈소판 막에서 분해되어 나오는 arach- idonic acid로부터 COX-1과 TXAS의 효소작용에 의해 만 들어진다. 따라서 이들에 미치는 G-Rg3의 효과를 함께 살 펴보았다. 그 결과 Fig. 4A 및 4B에서 보여주는 바와 같이 G-Rg3가 COX-1과 TXAS 활성을 억제하는 것을 확인할 수 있었다. Aspirin과 ozagrel은 양성대조군으로 사용되었 다. 각각의 사용농도는 Lee 등(25)의 방법을 참조하여 수행 하였다.
Fig. 3. Effects of G-Rg3 on TXA
2 production. Measurement of TXA2 were performed as described in ‘Materials and Methods’section. The data are expressed as the mean±standard deviation (n=4). *
P<0.05,
**P<0.01 versus the thrombin-stimulated human
platelets.A
B
Microsomal fraction (1 mg)
Microsomal fraction (1 mg)
Fig. 4. Effects of G-Rg3 on COX-1 and TXAS activity. (A)
Effects of G-Rg3 on COX-1 activity. (B) Effects of G-Rg3 on TXAS activity. Measurements of COX-1 and TXAS were per- formed as described in ‘Materials and Methods’ section. The data are expressed as the mean±standard deviation (n=4). *P<0.05
versus microsomal fraction.Fig. 5. Effects of G-Rg3 on arachidonic acid release. Arachidon-
ic acid release was determined as described in ‘Materials and Methods’ section. The data are expressed as the mean±standard deviation (n=4). *P<0.05 versus the thrombin-stimulated human
platelets.A
B
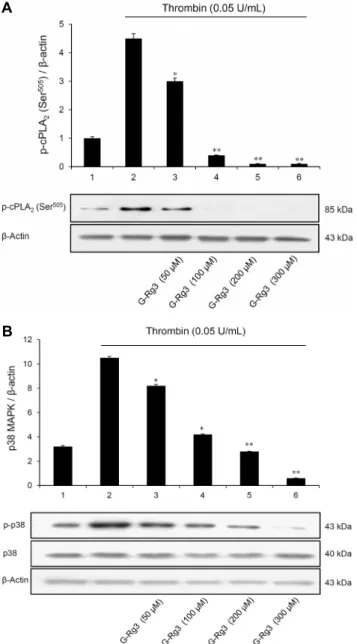
Fig. 6. Effects of G-Rg3 on cPLA
2- and p38 MAPK-phosphor- ylation. (A) Effect of G-Rg3 on cPLA2 phosphorylation. (B) Effects of G-Rg3 on p38 MAPK phosphorylation. Western blot was determined as described in ‘Materials and Methods’ section.The data are expressed as the mean±standard deviation (n=4).
*
P<0.05,
**P<0.01 versus the thrombin-stimulated human plate-
lets.G-Rg3가 arachidonic acid의 분비에 미치는 효과
G-Rg3가 억제하는 TXA2의 생성을 정확히 분석하기 위 하여 arachidonic acid 방출반응을 분석하였다. 그 결과 Fig. 5에서 보이는 바와 같이 thrombin으로 자극한 인체혈 소판은 arachidonic acid release의 양을 2,100.8±28.2 ng/108 platelets로 강하게 증가하였고, G-Rg3(50~300 μM) 는 농도 의존적인 억제 효과를 나타냈다. G-Rg3(300 μM) 는 arachidonic acid release를 1,044.2±15.6 ng/108 pla- telets로 50.3% 억제하였다.G-Rg3가 cPLA
2와 p38 MAPK 인산화에 미치는 효과
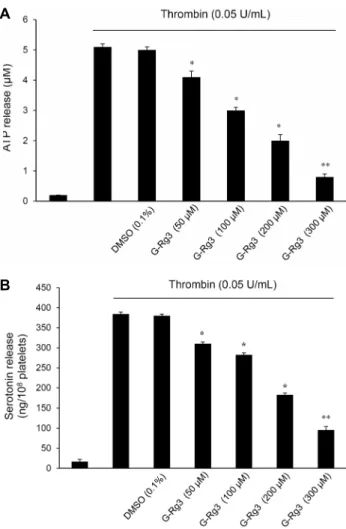
cPLA2는 arachidonic acid 방출의 중요한 매개체로 세포 질에서 혈소판 막으로 이동하여 혈소판 membrane의 ara- chidonic acid를 분해시킨다. 이때 MAPK 중 하나인 p38 MAPK는 cPLA2의 Ser505 위치를 인산화시키는 것으로 알 려져 있으며(13), p38 MAPK 또한 인산화를 통해서 혈소판 내에서 활성을 갖는 것으로 보고되었다(10). 따라서 G-Rg3 가 억제하는 arachidonic acid 방출반응이 상위 신호전달 분자인 p38 MAPK와 cPLA2의 인산화에 미치는 영향을 확 인하였다. Fig. 6A에서 보이는 바와 같이 thrombin(0.05 U/mL)으로 자극한 인체혈소판은 cPLA2의 Ser505 위치가 인산화되는 것을 확인할 수 있었으며(line 2), 이러한 인산화 는 G-Rg3(50~300 μM)에 의해 농도 의존적인 억제 양상을 나타내었다. cPLA2의 상위 신호전달 분자인 p38 MAPK 또 한 thrombin(0.05 U/mL)에 강한 인산화 양상을 보였지만, G-Rg3(50~300 μM)에 의해 농도 의존적인 억제 양상을 나 타내었다(Fig. 6B).G-Rg3가 ATP와 serotonin의 방출에 미치는 효과
G-Rg3가 혈소판 내 과립방출에 미치는 효과를 확인하기위하여 ATP release와 serotonin release를 분석하였다.
ATP release는 thrombin(0.05 U/mL)에 의해 5.1 μM로 증가하였지만, G-Rg3(50~300 μM)에 의해 농도 의존적으 로 억제되었다(Fig. 7A). Serotonin release 또한 throm- bin(0.05 U/mL)에 의해 384.4±4.9 ng/108 platelets로 증 가하였지만, G-Rg3(50~300 μM)에 의해 농도 의존적으로 억제되었다(Fig. 7B). G-Rg3(300 μM)의 ATP release와 serotonin release의 억제율은 각각 84.3%와 75.2%를 나 타내었다.
A
B
Fig. 7. Effects of G-Rg3 on ATP and serotonin release. (A) Ef-
fects of G-Rg3 on ATP release in thrombin-activated platelets.(B) Effects of G-Rg3 on serotonin release in thrombin-activated platelets. Determination of ATP and serotonin release was car- ried out as described in ‘Materials and Methods’ section. The data are expressed as mean±standard deviation (n=4). *
P<0.05,
**
P<0.01 versus the thrombin-stimulated human platelets.
고 찰
G-Rg3는 오래전부터 분리되어 다양한 생리활성이 보고 되어 왔으며 혈소판에 대해서도 부분적인 작용들이 보고된 바가 있지만(14,15), 인체혈소판에 미치는 효과에 대한 연 구는 미흡하였기에 본 연구에서는 이를 확실히 규명하고자 하였다. 그 결과 thrombin이 유도한 혈소판 응집에 있어서 G-Rg3는 농도 의존적으로 응집반응을 억제하고 있음을 보 여주었고(Fig. 2A), 혈소판에서 합성되어 분비되는 agonist 인 TXA2를 억제하는 결과를 나타내었다(Fig. 3). Thrombin 이 유도하는 혈소판의 활성화 과정에서 thrombin이 혈소판 막에 있는 수용체를 활성화시키면 막에 위치한 phosphati- dylinositol 4,5-bisphosphate를 분해하여 diacylglycer- ol(DG)을 방출시키고, DG-와 monoacylglycerol(MG)-li- pase의 효소작용으로 arachidonic acid가 되며, 분비된 arachidonic acid는 COX-1과 thromboxane A synthase
(TXAS)의 효소작용에 의해 TXA2로 전환되어 다른 혈소판 을 응집하는 강력한 agonist로 작용한다(5-7). 앞선 연구에 서 G-Rg3는 rat 혈소판의 TXA2 생성을 억제하는 효과가 보고되었다(15). 그러나 G-Rg3가 어떤 과정을 통해 TXA2
의 생성을 억제하였는지에 대한 연구 결과는 불명확하다.
본 연구에서는 G-Rg3가 TXA2의 생성에 미치는 영향을 살펴 보았다. 아울러 TXA2의 생성에 관여하는 COX-1 및 TXAS 의 활성과 arachidonic acid 분비 조절 효소인 cPLA2의 인 산화와 상위 신호전달 분자인 p38 MAPK에 G-Rg3가 관여 하는지 확인하였다.
그 결과 thrombin이 촉진시킨 cPLA2와 p38 MAPK의 인 산화는 G-Rg3에 의하여 농도 의존적으로 강하게 감소하였 고(Fig. 5A, 5B), arachidonic acid의 생성량도 G-Rg3에 의해 감소하는 것을 확인할 수 있었다(Fig. 4). 또한 TXA2
생성에 관여하는 효소인 COX-1과 TXAS의 활성은 G-Rg3 에 의해 억제되었고, 최종 대사산물인 TXA2가 G-Rg3에 의 해 농도 의존적으로 억제되는 결과를 나타냈다. 이 밖에도 p38 MAPK는 혈소판의 과립방출에 기여하는 것으로 보고 되었으며(26), G-Rg3가 억제한 ATP와 serotonin의 방출 반응은 G-Rg3가 억제한 p38 MAPK의 인산화에 의한 결과 로 보인다. 결론적으로 G-Rg3는 cPLA2와 p38 MAPK, arachidonic acid release, 그리고 TXA2의 합성효소인 COX-1과 TXAS를 억제함을 밝힘으로써 G-Rg3가 억제하 는 TXA2의 신호전달 기작을 규명하였다. 따라서 G-Rg3는 혈소판 응집으로 인한 심혈관계 질환에 있어서 치료 및 예방 약물로 유용한 가치가 있다고 여겨진다.
요 약
G-Rg3는 인삼의 성분으로서 많은 효과가 밝혀졌지만, 혈소 판에서의 억제 기작에 대한 연구는 미흡하다. 본 연구에서는 G-Rg3가 혈소판에서 합성되어 분비되는 TXA2의 생산에 관여한다고 알려진 효소인 COX-1, TXAS와 신호전달 분자 인 p38 MAPK, cPLA2의 인산화를 어떻게 조절하며 혈소판 응집을 억제시키는지 규명하고자 하였다. 그 결과 G-Rg3는 thrombin이 유도한 혈소판 응집을 강력하게 억제하였고, p38 MAPK, cPLA2의 인산화와 COX-1, TXAS의 활성을 감소시키며 arachidonic acid 분비와 TXA2의 합성을 농도 의존적으로 억제하였다. 또한 G-Rg3는 thrombin이 촉진시 킨 p38의 인산화를 억제함으로써 과립방출의 지표인 ATP 와 serotonin의 방출을 억제하였다. 따라서 G-Rg3는 인체 혈소판에서 TXA2의 생성을 억제함으로써 혈소판 응집억제 를 나타내는 치료 및 예방약물로서 유용한 가치가 있다고 여겨진다.
감사의 글
본 연구는 한국연구재단의 연구개발비(NRF-2011-0012
143 to Hwa-Jin Park)로 수행되어 이에 감사드립니다.
REFERENCES
1. Jackson SP. 2011. Arterial thrombosis-insidious, unpre- dictable and deadly. Nat Med 17: 1423-1436.
2. Schwartz SM, Heimark RL, Majesky MW. 1990. Develop- mental mechanisms underlying pathology of arteries. Physiol
Rev 70: 1177-1209.
3. Payrastre B, Missy K, Trumel C, Bodin S, Plantavid M, Chap H. 2000. The integrin αIIb/β3 in human platelet signal transduction. Biochem Pharmacol 60: 1069-1074.
4. Phillips DR, Nannizzi-Alaimo L, Prasad KS. 2001. Beta3 tyrosine phosphorylation in αIIbβ3 (platelet membrane GP IIb-IIIa) outside-in integrin signaling. Thromb Haemost 86:
246-258.
5. Needleman P, Moncada S, Bunting S, Vane JR, Hamberg M, Samuelsson B. 1976. Identification of an enzyme in pla- telet microsomes which generates thromboxane A2 from prostaglandin endoperoxides. Nature 261: 558-560.
6. Patrono C. 1994. Aspirin as an antiplatelet drug. N Engl
J Med 330: 1287-1294.
7. Cheng Y, Austin SC, Rocca B, Koller BH, Coffman TM, Grosser T, Lawson JA, FitzGerald GA. 2002. Role of pros- tacyclin in the cardiovascular response to thromboxane A2.
Science 296: 539-541.
8. Adam F, Kauskot A, Rosa JP, Bryckaert M. 2008. Mitogen- activated protein kinases in hemostasis and thrombosis. J
Thromb Haemost 6: 2007-2016.
9. Nadal-Wollbold F, Pawlowski M, Lévy-Toledano S, Berrou E, Rosa JP, Bryckaert M. 2002. Platelet ERK2 activation by thrombin is dependent on calcium and conventional protein kinases C but not Raf-1 or B-Raf. FEBS Lett 531: 475-482.
10. Kramer RM, Roberts EF, Strifler BA, Johnstone EM. 1995.
Thrombin induces activation of p38 MAP kinase in human platelets. J Biol Chem 270: 27395-27398.
11. Bugaud F, Nadal-Wollbold F, Lévy-Toledano S, Rosa JP, Bryckaert M. 1990. Regulation of c-jun-NH2 terminal kinase and extracellular-signal regulated kinase in human platelets.
Blood 94: 3800-3805.
12. McNicol A, Shibou TS. 1998. Translocation and phosphor- ylation of cytosolic phospholipase A2 in activated platelets.
Thromb Res 92: 19-26.
13. Kramer RM, Roberts EF, Um SL, Börsch-Haubold AG, Watson SP, Fisher MJ, Jakubowski JA. 1996. p38 mitogen- activated protein kinase phosphorylates cytosolic phospholi- pase A2 (cPLA2) in thrombin-stimulated platelets. Evidence that proline-directed phosphorylation is not required for mo- bilization of arachidonic acid by cPLA2. J Biol Chem 271:
27723-27729.
14. Matsuda H, Kubo M, Tani T, Arichi S, Kitagawa I. 1985.
Pharmacological study on Panax ginseng C.A. Meyer. V.
Effects of red ginseng on the experimental disseminated in- travascular coagulation. (4). On ginsenoside-Rg3, Rh1 and Rh2. Shoyakugaku Zasshi 39: 123-125.
15. Lee SR, Park JH, Kim ND, Choi KJ. 1997. Inhibitory effects of ginsenoside Rg3 on platelet aggregation and its mecha- nism of action. Korean J Ginseng Sci 21: 132-140.
16. Lee WM, Kim SD, Park MH, Cho JY, Park HJ, Seo GS, Rhee MH. 2008. Inhibitory mechanisms of dihydroginseno- side Rg3 in platelet aggregation: critical roles of ERK2 and cAMP. J Pharm Pharmacol 60: 1531-1536.
17. Jeong D, Irfan M, Kim SD, Kim S, Oh JH, Rhee MH. 2017.
Ginsenoside Rg3-enriched red ginseng extract inhibits plate- let activation and in vivo thrombus formation. J Ginseng
Res 41: 548-555.
18. Shin JH, Kwon HW, Cho HJ, Rhee MH, Park HJ. 2015.
Inhibitory effects of total saponin from Korean Red Ginseng on [Ca2+]i mobilization through phosphorylation of cyclic adenosine monophosphate-dependent protein kinase catalyt- ic subunit and inositol 1,4,5-trisphosphate receptor type Ⅰ in human platelets. J Ginseng Res 39: 354-364.
19. Born GVR, Hume M. 1967. Effects of the numbers and sizes of platelet aggregates on the optical density of plasma. Na-
ture 215: 1027-1029.
20. Born GVR. 1962. Quantitative investigations into the ag- gregation of blood platelets. J Physiol 162: 67-68.
21. Lee DH, Cho HJ, Kang HY, Rhee MH, Park HJ. 2012. Total saponin from Korean red ginseng inhibits thromboxane A2 production associated microsomal enzyme activity in platelets.
J Ginseng Res 36: 40-46.
22. Lee DH, Kim YJ, Kim HH, Cho HJ, Ryu JH, Rhee MH, Park HJ. 2013. Inhibitory effects of epigallocatechin-3-gal- late on microsomal cyclooxygenase-1 activity in platelets.
Biomol Ther 21: 54-59.
23. Barber AJ, Jamieson GA. 1970. Isolation and characterization of plasma membranes from human blood platelets. J Biol
Chem 245: 6357-6365.
24. Park JY, Ji HD, Jeon BR, Im EJ, Son YM, Lee JY, Lee DH, Lee YC, Hyun E, Jia Q, Hong M, Park HJ, Lee MH.
2013. Chlorin e6 prevents ADP-induced platelet aggregation by decreasing PI3K-Akt phosphorylation and promoting cAMP production. Evid-Based Compl Alt 2013: 569160.
25. Lee DH, Kim HH, Cho HJ, Yu YB, Kang HC, Kim JL, Lee JJ, Park HJ. 2014. Cordycepin-enriched WIB801C from
Cordyceps militaris inhibits collagen-induced [Ca
2+]i mobi- lization via cAMP-dependent phosphorylation of inositol 1, 4, 5-trisphosphate receptor in human platelets. Biomol Ther 22: 223-231.26. Flevaris P, Li Z, Zhang G, Zheng Y, Liu J, Du X. 2009.
Two distinct roles of mitogen-activated protein kinases in platelets and a novel Rac1-MAPK-dependent integrin out- side-in retractile signaling pathway. Blood 113: 893-901.