인간 췌장암세포주인 PANC-1에서 1,2,3,4,6-Penta-O-galloyl- β-D-glucose의 CD44 저해를 통한 항암 효과
⁃ 연구노트 ⁃
배민서1․김은영1․김윤희1,2,3
1대구대학교 식품영양학과
2대구대학교 항노화연구소
3대구대학교 산업기술연구소
Anti-Pancreatic Cancer Effect of 1,2,3,4,6-Penta-O-galloyl- β-D-glucose in PANC-1 through Inhibition of CD44
Min-Seo Bae1, Eun-Young Kim1, and Yoon Hee Kim1,2,3
1Department of Food and Nutrition, 2Research Institute of Anti-Aging, and
3Institute of Industrial and Technology, Daegu University
ABSTRACT Pancreatic cancer is currently the fifth leading cause of cancer-related mortality in South Korea in 2017.
1,2,3,4,6-Penta-O-galloyl-β-D-glucose (PGG) is a gallotannin isolated from plants such as Galla Rhois. PGG has an anti-allergic effect, an anti-inflammatory effect and an inhibitory effect in colon cancer. However, the effect of PGG on pancreatic cancer cells remains unclear. Thus, this study evaluated the effect of PGG in pancreatic cancer, and its molecular mechanisms were evaluated. PGG inhibited cell proliferation and induced cytotoxicity. Further, PGG suppressed clonogenic activity, adhesion to fibronectin and migration. PGG inhibited the protein expression of the cluster of differentiation 44 (CD44), which is a marker of pancreatic cancer stem cells. PGG also suppressed the expression of Nanog, Oct-4, and Sox-2, which are factors for cancer stem cells and they are down stream of the CD44 signaling pathway. This data suggests that PGG is a potentially effective functional compound as therapy for pancreatic cancer.
Key words: PGG, pancreatic cancer, CD44, cancer stem cell
Received 25 August 2019; Accepted 13 November 2019 Corresponding author: Yoon Hee Kim, Department of Food and Nutrition, College of Engineering, Daegu University, Gyeongsan, Gyeongbuk 38453, Korea
E-mail: [email protected], Phone: +82-53-850-6834 Author information: Min-Seo Bae (Graduate student), Eun-Young Kim (Student), Yoon Hee Kim (Professor)
서 론
국가 암 정보센터의 통계자료에 따르면 2016년 췌장암 발 생자수는 인구 10만 명당 13명으로 국내에서 9번째로 많이 발생하였다(National Cancer Information Center, 2019).
2017년 암으로 사망한 사람 중 전체의 22.8%는 폐암으로 사망하였으며, 다음으로 간암(13.6%), 대장암(11.1%), 위암 (10.2%), 췌장암(7.3%) 순으로 췌장암으로 인한 사망자수 는 5번째로 많았다(National Cancer Information Center, 2019). 또한 5년 상대생존율은 위암 76%, 대장암 75.9%, 간암 34.6%, 폐암 28.2%, 유방암 92.7%, 자궁경부암 79.8
%, 전립선암 93.9%인 것에 비해 췌장암은 11.4%로 5년 상 대생존율이 다른 암에 비해 현저히 낮았다(Korean Statis- tical Information Service, 2019). 췌장암은 초기에 자각증
상이 없거나 비특이적이기 때문에 초기진단이 어려워, 50~
60%의 환자에서 진단 시 원격전이나 혈관침윤 등이 동반되 는 경우가 많다(Sreedharan 등, 2019; Kleeff 등, 2016;
Huguet 등, 2014). 이로 인해 췌장암은 가장 효과적인 치료 방법인 수술을 시행할 수 없는 경우가 많다. 수술을 시행한 후나 수술을 할 수 없는 경우 췌장암 치료를 위해 많이 사용 하는 gemcitabine이나 5-fluorouracil(FU)과 같은 항암요 법이나 방사선요법 등을 병행해야 하나 이에 대한 저항성이 높아 치료방법이 제한적이기 때문에 5년 생존율이 다른 암 종에 비해 낮다(Kleeff 등, 2016; Jeune 등, 2019; Lanfre- dini 등, 2019). 따라서 췌장암의 생존율 증진에 도움을 줄 수 있는 새로운 물질에 대한 탐색이 필요하다.
Cluster of differentiation 44(CD44)는 포유류 세포막에 존재하는 당단백질로 췌장암에서 과발현하며(Mattheola- bakis 등, 2015; Takada 등, 1994; Immervoll 등, 2011), 막수용체인 CD44를 통한 신호전달은 췌장암에서 암세포 생 존, 증식, 이동, 침윤 및 다중약물내성(multidrug resist- ance)을 일으킨다(Itano 등, 2008; Sironen 등, 2011). 또한 CD44는 췌장암 줄기세포의 대표적인 표지자로 췌장암의 전 이, 재발, 약물내성에 관여한다(Fitzgerald와 McCubrey,
Fig. 1. Chemical structure of 1,2,3,4,6-penta-O-galloyl-β-D-glu- cose (PGG).
2014). 따라서 췌장암세포에서 CD44를 억제하는 것은 췌 장암 치료를 위한 하나의 치료 전략일 수 있다.
1,2,3,4,6-Penta-O-galloyl-β-D-glucose(PGG, Fig. 1) 는 오배자(五倍子, Galla Rhois)라고 불리는 옻나무과의 낙 엽소교목에서 추출・분리한 gallotannin 성분으로(Kwon 등, 2013), 항알레르기 효과(Kim 등, 2015; Kim 등, 2016), 항 염증 효과(Lee 등, 2017), 대장암 억제 효과(Kawk 등, 2018) 등이 알려져 있다.
본 연구에서는 PGG의 췌장암세포주인 PANC-1에 있어 서 CD44를 통한 PGG의 항암 효과 기전에 대해 검토하였다.
재료 및 방법
실험 재료
세포배양에 사용되는 fetal bovine serum(FBS), pen- icillin, streptomycin, Dulbecco’s modified Eagle’s me- dium(DMEM), Antibiotic Antimycotic solution, 0.5% Tryp- sin-EDTA는 HyClone(Logan, UT, USA)에서 구매하였 다. CytoTox96® Non-Radioactive Cytotoxicity Assay kit과 CellTiter96®Aqueous One Solution Cell Prolifer- ation Assay kit은 Promega(Madison, WI, USA)에서 구입 하여 사용하였다. Crystal violet과 glutaraldehyde 용액은 Sigma-Aldrich Co.(St. Louis, MO, USA)로부터 구매하였 다. CytoSelectTM 48-well Cell Adhesion Assay kit(Fi- bronectin, Colorimetric kit)과 CytoSelectTM 24-well Cell Migration Assay kit(8 μm, Colorimetric kit)은 CellBio- labs Inc.(San Diego, CA, USA)에서 구매하여 사용하였다.
Western blot 분석에 사용한 일차 항체 anti-CD44는 Abcam (Cambridge, UK)에서, anti-Nanog, anti-Oct-4와 anti- Sox-2는 Santa Cruz Biotechnology Inc.(Dallas, TX, USA) 에서, anti-β-actin은 Novus Biologicals(Centennial, CO, USA)에서 구매하였다. 이차 항체인 anti-rabbit IgG-HRP, anti-mouse IgG-HRP는 Cell Signaling Technology(Dan- vers, MA, USA)에서 구매하여 사용하였다. TRIzol® re- agent는 Abcam(Carlsbad, CA, USA)에서 구매하였다. RNA
PrimeScriptTM RT Reagent kit은 Takara(Tokyo, Japan) 에서 구매하였다. LightCycler® 480 SYBR Green 1 Master Mix는 Roche Diagnostics(Mannheim, Germany)에서 구매 하였다. Immobilon® transfer membrane(0.45 μm)은 Mil- lipore(Bedford, MA, USA)에서 구매하였다.
PGG 분리
본 실험에 사용된 PGG는 오배자로부터 Kwon 등(2013) 에 의해 서술된 방법으로 추출・분리한 시료를 실험에 사용 하였고, 순도는 98% 이상이었다.
세포주 및 세포배양
PANC-1 세포는 한국세포주은행(Korea Cell Line Bank, Seoul, Korea)에서 분양받아 실험에 사용하였다. 세포는 10% FBS, 1% Antibiotic Antimycotic solution을 함유하 는 DMEM 배지에서 배양하였다. 세포는 37°C, 5% CO2 조 건하에서 2~3일에 한 번씩 계대배양하였다.
췌장암세포의 세포증식 및 독성 측정
PGG가 PANC-1의 세포증식 및 세포독성에 미치는 영향 을 검토하기 위하여 PANC-1 세포를 96-well plate에 1×
104 cells/200 μL/well의 세포수가 되도록 분주하여 37°C, 5% CO2 배양기에서 24시간 배양하였다. PGG를 농도별(0, 1, 2.5, 5, 10 μM)로 처리한 후 48~96시간 동안 배양하였다.
배양 후 CellTiter96® Aqueous One Solution Assay of Cell Proliferation(Promega)을 이용해 세포증식능을 mi- croplate reader(Sunrise・Basic Tecan, Tecan Austria, Salzburg, Austria)를 사용하여 측정하였다. 또한, CytoTox 96® Non-Radioactive Cytotoxicity Assay(Promega)을 이용해 PGG가 세포에 독성을 나타내는지를 microplate reader(Sunrise・Basic Tecan)를 사용하여 측정하였다. 세 포증식은 490 nm에서 측정된 흡광도 값으로, 세포독성은 양성대조군의 최대 lactate dehydrogenase(LDH) 방출에 대한 실험군의 LDH 방출 비율로 계산하였고 계산식은 아래 와 같다.
세포독성(%) = 실험군의 LDH 방출(OD) 양성대조군의 최대 LDH 방출(OD)×100
췌장암세포의 군집 형성능 측정
PGG가 췌장암세포의 군집 형성에 미치는 영향을 검토하 기 위해 PANC-1 세포를 6-well plate에 1×103 cells/2 mL/well의 세포수가 되도록 분주하여 37°C, 5% CO2 배양 기에서 24시간 배양하였다. PGG를 농도별(0, 1, 2.5, 5, 10 μM)로 처리한 후 3일마다 PGG를 함유한 새로운 배지로 교 환하며 총 17일 동안 배양하였다. 배양 후 PBS로 1회 세척 한 다음 6% glutaraldehyde와 5% crystal violet을 1:1로 혼합한 용액으로 세포를 염색하고 건조하여 사진 촬영을 하 였다.
췌장암세포의 부착 및 이동 측정
PGG가 췌장암세포의 fibronectin에 대한 세포 부착능력 에 미치는 영향을 검토하기 위해 CytoSelectTM 48-well Cell Adhesion Assay kit(Fibronectin, Colorimetric kit) 을 사용하였다. PGG를 농도별로 처리한 PANC-1 세포를 1×105 cells/500 μL/well의 세포수가 되도록 fibronectin 이 코팅된 48-well plate에 분주하여 37°C, 5% CO2 배양기 에서 90분간 배양하였다. 배양 후 부착되지 않은 세포는 PBS로 세척하고 kit에 포함된 staining solution으로 세포 핵을 염색시키고 extraction solution으로 세포를 용해하여 fibronectin에 부착된 세포들을 microplate reader(Sunrise
・Basic Tecan)를 사용하여 560 nm에서 흡광도를 측정하 였다.
PGG가 췌장암세포의 이동능력에 미치는 영향을 검토하 기 위해 CytoSelectTM 24-well Cell Migration Assay kit (8 μm pore size, Colorimetric Kit)을 사용하였다. Kit의 구성품인 polycarbonate membrane inserts(8 μm pore size), 즉 upper chamber에 serum free media로 PGG를 농도별로 처리한 PANC-1 세포를 5×104 cells/250 μL/
well의 세포수가 되도록 분주하였다. 10% FBS를 함유한 DMEM 배지를 plate의 lower well에 500 μL씩 넣었다.
37°C, 5% CO2 배양기에서 48시간 동안 배양하였다. Insert 안의 배지를 제거한 후 lower well로 이동되지 않고 inserts 에 남은 세포를 젖은 면봉으로 제거하였다. 그 후 lower well 쪽으로 이동한 세포는 Cell Stain Solution으로 10분간 염색한 다음 extraction solution으로 염색된 세포의 염색용 액을 추출하여 microplate reader(Sunrise・Basic Tecan) 를 사용하여 560 nm에서 흡광도를 측정하였다.
Western blot 분석
PGG가 항암 효과를 나타내는 기전을 밝히기 위해 PANC- 1 세포를 6-well plate에 1×105 cells/3 mL/well의 세포수 가 되도록 분주하고, 37°C, 5% CO2 배양기에서 24시간 배 양하였다. PGG를 농도별로 처리한 후 72시간 배양하였다.
세포를 PBS로 세척한 후 cell lysis buffer를 이용해 세포용 해물을 얻었다. 단백질 농도는 bovine serum albumin (BSA)을 표준화한 Bio-Rad Protein Assay kit(Bio-Rad, Hercules, CA, USA)을 사용하여 정량하였다. 단백질 농도 가 같도록 조정한 후 2×sodium dodecyl sulfate(SDS) sample buffer를 가하고 100°C에서 5분간 끓였다. Mini- PROTEAN system(Bio-Rad)을 이용하여 10% SDS poly- acrylamide gel을 만들어 각 well에 10 μg에 해당하는 단백 질이 되도록 각 시료를 loading 하고 전기영동을 시행하였 으며 이어서 Immobilon® transfer membranes 0.45 μm (Millipore)에 transfer 시켰다. Membrane을 1×blocking buffer(Biofact Biofactory, Daejeon, Korea)로 실온에서 1시간 동안 blocking 한 후 일차항체 anti-CD44(1:3,000,
Cell Signaling Technology), anti-Nanog(1:1,000, Santa Cruz Biotechnology Inc.), anti-Oct-4(1:1,000, Santa Cruz Biotechnology Inc.), anti-Sox-2(1:1,000, Santa Cruz Biotechnology Inc.), anti-β-actin(1:10,000, Novus Biologicals)을 각각 처리하여 4°C에서 하룻밤 배양하였다.
Tris-buffered saline containing 0.1% Tween-20(TBST, pH 8.0)로 세척한 다음 이차항체(HRP-conjugated anti- rabbit IgG, anti-mouse IgG, 1:5,000, Cell Signaling Technology)에 1시간 동안 실온에서 배양하였다. TBST로 세척하고 ECL solution(Thermo Scientific, Rockford, IL, USA) 기질로 반응시킨 후 LAS-3000 Lumino Image Ana- lyzer System(Fujifilm, Tokyo, Japan)을 이용하여 밴드를 확인하였다.
RNA 추출 및 cDNA 합성
PGG가 항암 효과를 나타내는 기전을 밝히기 위해 PANC -1 세포를 6-well plate에 1×105 cells/3 mL/well의 세포 수가 되도록 분주하고, 37°C, 5% CO2 배양기에서 24시간 배양하였다. PGG를 농도별로 처리한 후 72시간 배양하였 다. 세포배양액을 제거한 다음 PBS로 세척 후 TRIzol® re- agent를 이용하여 총 RNA를 추출하였다. RNA 정량 후 RNA PrimeScriptTM RT Reagent kit(Takara)을 사용하여 cDNA 를 합성하였다.
Quantitative real time-PCR 분석
CD44 mRNA 발현량을 검토하기 위해서 LightCycler® 480 SYBR Green 1 Master Mix(Roche Diagnostics)를 사용하여 LightCycler®96 system(Roche Diagnostics)으 로 측정하였다. PCR 반응의 특이도를 확인하기 위해서 증폭 할 때마다 melting curve 분석을 수행하였고 β-actin을 기 준으로 2-ΔCt로 계산하였으며 유전자의 발현량은 대조군을 1로 간주하여 상대적인 값을 측정하였다. Primer는 Bioneer (Daejeon, Korea)에서 제작하여 사용하였고 염기서열은 다 음과 같다: CD44 Forward 5’-GAGAGCTGGCCAAGTCT TCA-3’, Reverse 5’-GTCAAGCTGTGCTTCCAGAGT TA-3’, β-actin Forward 5’-CATCCGTAAAGACCTCT ATGCCAA-3’, Reverse 5’-ATGGAGCCACCGATCCAC A-3’.
통계분석
실험에서 얻어진 결과의 통계적 유의성은 GraphPad Prism Version 5.0 software(GraphPad, San Diego, CA, USA)를 이용해 분석하였다. 실험 결과는 mean±standard error of mean(SEM)으로 나타냈으며, 각 그룹 간의 통계적 유의성을 one-way ANOVA를 통한 Dunnett’s test 또는 two-way ANOVA를 통한 Bonferroni test를 통해서 검정 하였다. P 값은 P<0.05를 통계적 유의차가 있다고 판단했다.
A B
C PGG (μM) Gem (μM) 5-FU (μM)
Fig. 2. Effect of PGG on (A) cell proliferation, (B) cell cytotoxicity, and (C) colony formation of PANC-1. PANC-1 cells were treated with the indicated concentration of PGG, gemcitabine (Gem), or 5-FU for 48∼96 h or 17 days. Cell proliferation and cytotoxicity were determined by One Solution Assay kit or Non-Radioactive Cytotoxicity Assay kit, respectively. Colony formation assay was determined by crystal violet staining. The data are reported as the mean±SEM (n=4). Statistical significance is based on the difference compared with 0 μM PGG by (A, B) two-way ANOVA followed by Bonferroni test (*P<0.05, **P<0.01, ***P<0.001).
결과 및 고찰
PGG가 PANC-1의 세포증식, 독성 및 군집 형성에 미치는 영향
PGG가 PANC-1의 세포증식에 미치는 영향을 검토하기 위해 0, 1, 2.5, 5, 10 μM 농도로 PGG를 48, 72, 96시간 동안 처리한 결과, 72시간부터 1 μM의 PGG를 처리한 군에 서부터 농도 의존적으로 췌장암세포의 성장을 억제하였다 (Fig. 2A, P<0.05). 또한 PGG가 PANC-1의 세포독성에 미 치는 영향을 측정한 결과(Fig. 2B), PGG를 72시간 동안 처 리했을 때 1 μM의 PGG에서부터 농도 의존적으로 췌장암세 포에 대해 독성을 보였다(P<0.05). 암줄기세포의 특징 중 하나는 자기재생능력(self-renewal potential)으로, 췌장암 줄기세포에서도 이러한 자기재생능력이 높다(Barkeer 등, 2018). 이러한 암줄기세포의 자기재생능력을 측정하기 위 한 in vitro 실험법으로 serial colony-forming unit(CFU)을 사용한다(Clarke 등, 2006). 이에 PGG가 PANC-1의 군집 형성능력에 미치는 영향을 측정하기 위해 PANC-1 세포에 0, 1, 2.5, 5, 10 μM PGG를 17일 동안 처리하여 형성된 군집을 crystal violet으로 염색한 결과(Fig. 2C), 1 μM PGG 부터 PANC-1의 군집 형성을 현저히 감소하였다. 이상의 결과로부터 PGG는 췌장암세포의 증식을 억제하고 독성을 유도할 뿐만 아니라 암 줄기세포의 특징 중 하나인 군집 형 성을 억제하였다.
PGG가 PANC-1의 부착능 및 이동성에 미치는 영향 암줄기세포는 무한증식, 자가재생능력, 세포자살에 대한 저항성과 같은 특징 이외에도 세포 이동성이 증가한다(Rodri- guez-Aznar 등, 2019). 특히 췌장암이 치료하기 어려운 이 유는 췌장암 자체의 성장 때문만이 아니라 진단 시 50~60%
의 환자에게서 인근 장기로의 광범위한 전이가 동반되는 경 우가 많기 때문이다(Walter 등, 2019; Sreedharan 등, 2019;
Kleeff 등, 2016; Huguet 등, 2014). 이처럼 췌장암의 이동 성 증가가 암의 나쁜 예후와 관련이 있기 때문에(Walter 등, 2019) 암세포가 이동하기 위해 필요한 부착능과 이동성에 대해 검토하였다. Fibronectin은 세포외기질에 풍부한 단백 질로 세포막에 있는 integrin adhesion receptor에 결합하 여 세포의 부착, 이동 등과 같은 다양한 세포 활동을 중재한 다(Erdogan 등, 2017). 따라서 PGG가 fibronectin에 대한 PANC-1의 부착능에 미치는 영향을 측정한 결과(Fig. 3A), 1 μM PGG 처리에 의해 fibronectin에 대한 PANC-1의 부 착이 유의적으로 억제되었다(P<0.001). 다음으로 췌장암의 이동성에 미치는 영향을 검토하기 위해 transwell을 사용하 여 측정하였다. 구체적으로 본 연구에 사용한 kit은 poly- carbonate membrane(8 μm pore size)을 중심으로 mem- brane 위쪽에는 serum free media를 포함한 췌장암세포가 membrane 아래쪽에 10% FBS를 포함한 media 쪽으로 얼 마나 많이 이동하는지를 측정하였다. 이 방법으로 PGG가 PANC-1의 이동성에 미치는 영향을 측정한 결과(Fig. 3B),
A
B
Fig. 3. Effect of PGG on (A) cell adhesion and (B) cell migration of PANC-1. PANC-1 cells were treated with the indicated con- centration of PGG, gemcitabine (Gem), or 5-FU for 90 min or 48 h, respectively. Adhesion of PANC-1 against fibronection was evaluated by 48-well Cell Adhesion Assay kit. Migration of PANC-1 were determined by 24-well Cell Migration Assay kit (8 μm pore size). The data are reported as the mean±SEM (n=4). Statistical significance is based on the difference com- pared with 0 μM PGG by one-way ANOVA followed by Dun- nett’s test (*P<0.05, **P<0.01, ***P<0.001).
A
B
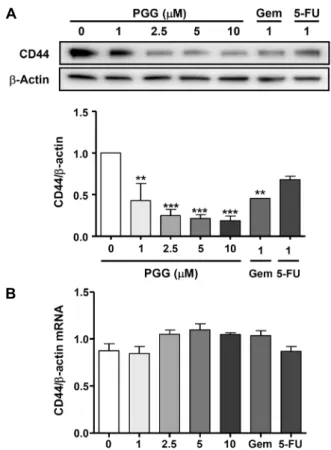
Fig. 4. PGG inhibits expression of (A) CD44 protein, not (B) CD44 mRNA. PANC-1 cells were treated with the indicated con- centration of PGG, gemcitabine (Gem), or 5-FU for 72 h. (A) Protein of whole cell lysates was resolved on 10% SDS-PAGE for CD44. β-Actin expression is shown as a lodging control.
Relative values over western blot band are indicated by the in- tensity of band of CD44 divided by density of band of β-actin.
(B) Total RNA was extracted and transcribed to cDNA. Quanti- tative PCR was performed with the incorporation of SYBR green into the double-strained PCR products. The mean relative expressions of the CD44 were calculated and differences were determined using the 2-ΔCt method. The data are reported as the mean±SEM (n=3). Statistical significance is based on the differ- ence compared with 0 μM PGG by one-way ANOVA followed by Dunnett’s test (**P<0.01, ***P<0.001).
1 μM의 PGG 처리군부터 PANC-1의 이동이 유의적으로 억제되었다(P<0.05). 이상의 결과로부터 PGG는 암세포의 주요 특징인 부착능력과 이동성을 억제하였다.
PGG가 CD44 발현 및 CD44 신호전달 체계에 미치는 영향 CD44는 암줄기세포의 표지자로 췌장암에서 과발현하며, stem-like cell receptor로 작용하여 췌장암의 진행, 전이, 약물내성, 나쁜 예후와 관련된다(Wei 등, 2011; Mattheo- labakis 등, 2015). 따라서 암줄기세포 특이적인 표지자인 CD44의 단백질 발현 억제는 췌장암 치료의 방법 중 하나일 수 있다(Yan 등, 2016). 이에 PGG가 CD44의 단백질 발현 에 미치는 영향을 측정한 결과(Fig. 4A), 1 μM PGG 처리에 의해 CD44의 단백질 발현이 유의적으로 감소하였다(P<
0.01). 췌장암의 항암치료에 많이 사용되는 gemcitabine이 나 5-FU는 PGG보다 효과가 미비하였다. 하지만 PGG는 CD44 mRNA의 발현에는 영향을 미치지 않았다(Fig. 4B).
이상의 결과로부터 PGG는 CD44의 단백질 분해를 촉진하는 것으로 생각된다. Yang 등(2019)의 연구에 의하면 PANC- 1 세포에서 PGG가 insulin 수용체를 통한 신호전달을 억제 함으로써 췌장암세포를 이식한 동물의 간과 근육조직에서
해당과정, 포도당 신생합성, 단백질 분해와 지질 분해를 억 제하여 암악액질을 개선하였다고 보고하였으나 CD44 발현 에 대한 연구는 본 연구가 처음으로 그 의미가 있다고 하겠 다. 한편 Nuñez-Sánchez 등(2017)에 의하면 대장암 환자 의 정상 조직에 비해 암 조직에서 CD44가 과발현하였으며, gallotannin이 포함된 석류 추출물을 2주 동안 하루 900 mg 씩 섭취한 결과 대장암 조직에서 CD44의 발현이 감소한다 고 하였다. 또한 Jia 등(2017)의 연구에 따르면 난소암세포 주에서 Phyllanthus niruri L., P. emblica L.과 P. urinaria L. 등에 함유되어 있는 gallotannin인 corilagin이 CD44의 단백질 발현을 농도 의존적으로 감소한다고 하였다.
CD44를 통한 신호전달체계는 암의 부착, 이동, 증식 및 분화와 같은 다양한 역할을 하는 것으로 알려져 있다(Yan 등, 2016). Li 등(2014)에 의하면 췌장암세포주에 CD44 항
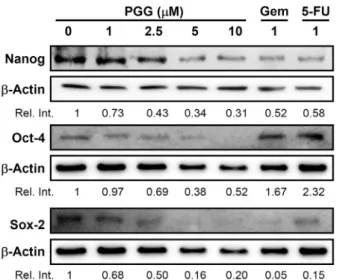
Fig. 5. PGG suppresses CD44 mediated signaling pathway.
PANC-1 cells were treated with the indicated concentration of PGG, gemcitabine (Gem), or 5-FU for 72 h. Protein of whole cell lysates was resolved on 10% SDS-PAGE for Nanog, Oct-4, and Sox-2. β-Actin expression is shown as a lodging control.
Relative values over western blot band are indicated by the in- tensity of band of target protein divided by density of band of β-actin.
체 처리 시 Nanog와 Sox2와 같은 자가재생 관련 유전자를 억제한다고 하였다(Li 등, 2014). 또한 Wang 등(2014)에 의하면 Oct-4와 Nanog는 유방암 환자에게서 나쁜 예후에 관련되며, Oct-4와 Nanog의 억제는 암줄기세포의 이동을 억제한다고 하였다(Wang 등, 2014). 이에 CD44 하위 신호 전달 경로이며 줄기세포의 표지자(Rodriguez-Aznar 등, 2019)인 Nanog, Oct-4 및 Sox-2 발현에 미치는 PGG의 영향을 검토하였다. PGG를 PANC-1에 72시간 처리했을 때 Nanog, Oct-4, Sox-2의 단백질 발현을 억제하였다 (Fig. 5). 이상의 결과로부터 PGG는 CD44의 발현을 억제하 고 그 하위 신호전달경로인 Nanog-Oct-4-Sox-2의 발현 을 억제하여 췌장암의 증식, 부착 및 이동 등을 억제하는 것으로 생각된다.
요 약
PGG는 오배자(五倍子, Galla Rhois)의 gallotannin으로 항 알레르기 효과, 항염증 효과, 대장암 억제 효과 등을 가지는 것으로 알려져 있다. 췌장암은 5년 상대생존율이 11.4%로 다른 암에 비해 현저히 낮으며 수술을 할 수 없는 경우가 많고 항암요법에도 저항성이 높아 치료방법이 제한적이다.
CD44는 막에 존재하는 당단백질로 암줄기세포의 표지자이 고 췌장암세포에서도 과발현하는 것으로 알려져 PGG를 통 한 CD44의 발현 억제는 췌장암의 치료 효과를 기대할 수 있다. 따라서 본 연구에서는 PGG가 췌장암세포주인 PANC- 1 세포에 대한 영향을 조사하고 CD44를 통한 관련 메커니즘 에 대해 평가하였다. PGG는 PANC-1의 증식을 억제하고
암세포독성을 유도하였으며 군집 형성을 억제하였다. 또한 막 부착분자인 fibronectin에 대한 PANC-1의 부착능 및 transwell을 통한 이동능이 억제되었다. 이러한 PGG의 췌 장암 억제 효과에 대한 분자적 기전을 확인한 결과 췌장암 줄기세포의 표지자인 CD44, Nanog, Sox-2 및 Oct-4의 단백질 발현을 억제하였다. 이러한 결과로부터 gallotannin 의 하나인 PGG가 췌장암 줄기세포 표지자인 CD44의 발현 억제 효과를 가지는 것으로 나타나, 향후 췌장암을 치료하는 데 유용한 물질로 사용될 가능성이 있을 것으로 생각된다.
감사의 글
이 논문은 한국연구재단(NRF-2016R1D1A1B03930131) 및 대구대학교 연구 장학기금 지원에 의한 것이며 이에 감사 드립니다.
REFERENCES
Barkeer S, Chugh S, Karmakar S, Kaushik G, Rauth S, Rachagani S, et al. Novel role of O-glycosyltransferases GALNT3 and B3GNT3 in the self-renewal of pancreatic cancer stem cells.
BMC Cancer. 2018. 18:1157. doi: 10.1186/s12885-018-5074-2.
Clarke MF, Dick JE, Dirks PB, Eaves CJ, Jamieson CH, Jones DL, et al. Cancer stem cells-perspectives on current status and future directions: AACR Workshop on cancer stem cells.
Cancer Res. 2006. 66:9339-9344.
Erdogan B, Ao M, White LM, Means AL, Brewer BM, Yang L, et al. Cancer-associated fibroblasts promote directional can- cer cell migration by aligning fibronectin. J Cell Biol. 2017.
216:3799-3816.
Fitzgerald TL, McCubrey JA. Pancreatic cancer stem cells: asso- ciation with cell surface markers, prognosis, resistance, meta- stasis and treatment. Adv Biol Regul. 2014. 56:45-50.
Huguet F, Mukherjee S, Javle M. Locally advanced pancreatic cancer: the role of definitive chemoradiotherapy. Clin Oncol.
2014. 26:560-568.
Immervoll H, Hoem D, Steffensen OJ, Miletic H, Molven A.
Visualization of CD44 and CD133 in normal pancreas and pancreatic ductal adenocarcinomas: non-overlapping mem- brane expression in cell populations positive for both markers.
J Histochem Cytochem. 2011. 59:441-455.
Itano N, Zhuo L, Kimata K. Impact of the hyaluronan-rich tumor microenvironment on cancer initiation and progression. Can- cer Sci. 2008. 99:1720-1725.
Jeune F, Coriat R, Prat F, Dousset B, Vaillant JC, Gaujoux S.
Pancreatic cancer surgical management. Presse Med. 2019.
48:e147-e158.
Jia L, Zhou J, Zhao H, Jin H, Lv M, Zhao N, et al. Corilagin sensitizes epithelial ovarian cancer to chemotherapy by in- hibiting Snail-glycolysis pathways. Oncol Rep. 2017. 38:2464- 2470.
Kawk SH, Kang YR, Kim YH. 1,2,3,4,6-Penta-O-galloyl-β-d- glucose suppresses colon cancer through induction of tumor suppressor. Bioorg Med Chem Lett. 2018. 28:2117-2123.
Kim YH, Choi YR, Kim JY, Kwak SH. Anti-allergic effect of 1,2,3,4,6-penta-O-galloyl-β-D-glucose on RBL-2H3 cells. J Ko- rean Soc Food Sci Nutr. 2016. 45:613-618.
Kim YH, Yang X, Yamashita S, Kumazoe M, Huang Y, Naka- hara K, et al. 1,2,3,4,6-Penta-O-galloyl-β-D-glucopyranose in-
creases a population of T regulatory cells and inhibits IgE pro- duction in ovalbumin-sensitized mice. Int Immunopharmacol.
2015. 26:30-36.
Kleeff J, Korc M, Apte M, La Vecchia C, Johnson CD, Biankin AV, et al. Pancreatic cancer. Nat Rev Dis Primers. 2016. 2:
16022.
Korean Statistical Information Service. [cited 2019 Jan 15].
Available from: http://kosis.kr/statHtml/statHtml.do?orgId=117
&tblId=DT_117N_A00021&vw_cd=MT_ZTITLE&list_id=1 01_11744&seqNo=&lang_mode=ko&language=kor&obj_var _id=&itm_id=&conn_path=MT_ZTITLE
Kwon OJ, Bae JS, Lee HY, Hwang JY, Lee EW, Ito H, et al.
Pancreatic lipase inhibitory gallotannins from Galla Rhois with inhibitory effects on adipocyte differentiation in 3T3-L1 cells.
Molecules. 2013. 18:10629-10638.
Lanfredini S, Thapa A, O’Neill E. RAS in pancreatic cancer.
Biochem Soc Trans. 2019. doi: 10.1042/BST20170521.
Lee HW, Kang YR, Bae MS, Kim YH. Anti-inflammatory ef- fects of 1,2,3,4,6-penta-O-galloyl-β-d-glucose in LPS-stimu- lated macrophages. J Korean Soc Food Sci Nutr. 2017. 46:
409-416.
Li L, Hao X, Qin J, Tang W, He F, Smith A, et al. Antibody against CD44s inhibits pancreatic tumor initiation and post- radiation recurrence in mice. Gastroenterology. 2014. 146:1108- 1118.
Mattheolabakis G, Milane L, Singh A, Amiji MM. Hyaluronic acid targeting of CD44 for cancer therapy: from receptor biol- ogy to nanomedicine. J Drug Target. 2015. 23:605-618.
National Cancer Information Center. [cited 2019 Jan 2]. Avail- able form: https://www.cancer.go.kr/lay1/S1T639C641/contents.
do
Nuñez-Sánchez MA, González-Sarrías A, García-Villalba R, Monedero-Saiz T, García-Talavera NV, Gómez-Sánchez MB, et al. Gene expression changes in colon tissues from colorectal cancer patients following the intake of an ellagitannin-con- taining pomegranate extract: a randomized clinical trial. J Nutr Biochem. 2017. 42:126-133.
Rodriguez-Aznar E, Wiesmüller L, Sainz B Jr, Hermann PC.
EMT and stemness-key players in pancreatic cancer stem cells.
Cancers (Basel). 2019. 11:1136. doi: 10.3390/cancers11081136.
Sironen RK, Tammi M, Tammi R, Auvinen PK, Anttila M, Kosma VM. Hyaluronan in human malignancies. Exp Cell Res. 2011. 317:383-391.
Sreedharan L, Kumar B, Jewell A, Banim P, Koulouris A, Hart AR. Bridging clinic: The initial medical management of pa- tients with newly diagnosed pancreatic cancer. Frontline Gas- troenterol. 2019. 10:261-268.
Takada M, Yamamoto M, Saitoh Y. The significance of CD44 in human pancreatic cancer: Ⅱ. The role of CD44 in human pancreatic adenocarcinoma invasion. Pancreas. 1994. 9:753- 757.
Walter K, Tiwary K, Trajkovic-Arsic M, Hidalgo-Sastre A, Dierichs L, Liffers ST, et al. MEK inhibition targets cancer stem cells and impedes migration of pancreatic cancer cells in vitro and in vivo. Stem Cells Int. 2019. https://doi.org/10.
1155/2019/8475389
Wang D, Lu P, Zhang H, Luo M, Zhang X, Wei X, et al. Oct-4 and Nanog promote the epithelial-mesenchymal transition of breast cancer stem cells and are associated with poor prog- nosis in breast cancer patients. Oncotarget. 2014. 5:10803- 10815.
Wei HJ, Yin T, Zhu Z, Shi PF, Tian Y, Wang CY. Expression of CD44, CD24 and ESA in pancreatic adenocarcinoma cell lines varies with local microenvironment. Hepatobiliary Pan- creat Dis Int. 2011. 10:428-434.
Yan Y, Li Z, Kong X, Jia Z, Zuo X, Gagea M, et al. KLF4- mediated suppression of CD44 signaling negatively impacts pancreatic cancer stemness and metastasis. Cancer Res. 2016.
76:2419-2431.
Yang J, Wang F, Chen X, Qiu S, Cui L, Hu L. β-Pentagalloyl- glucose sabotages pancreatic cancer cells and ameliorates ca- chexia in tumor-bearing mice. Am J Chin Med. 2019. 47:675- 689.