J. Exp. Biomed. Sci. 2013, 19(1): 9~16 pISSN : 1738-3226
Characterization of Matrix Metalloproteinase Expression in Triglyceride Treated THP-1 Macrophages
Yoonjung Cho1,#, Jaewon Lim1,#, Dong Hyun Lee1, Byung Chul Jung1, Min Ho Lee1, Dongju Jung2, Yoon Suk Kim1, Tae Ue Kim1,† and Ki-Jong Rhee1,†
1Department of Biomedical Laboratory Science, College of Health Sciences, Yonsei University, Wonju 220-710, Korea
2Institute for Integrated Cell-Material Sciences (WPI-iCeMS), Kyoto University, Kyoto 606-8501, Japan
Elevated blood triglyceride (TG) levels correlate with development of atherosclerosis suggesting that TG may promote the development of this disease. During atherosclerosis, TG is taken up by tissue macrophages which result in dramatic changes in various secreted factors. One such factor is the family of matrix metalloproteases (MMP) which are involved in tissue remodeling during both physiological and pathological processes. In this study, we examined the MMP expression profile in PMA-differentiated THP-1 macrophages treated with TG. We found that TG-treated THP-1 macrophages showed decreased expression of MMP-3, MMP-7, MMP-8 and MMP-9 in a time- and dose-dependent manner. In contrast, expression of MMP-1, MMP-2, and MMP-10 remained relatively unchanged after TG treatment. In addition, we found that expression of select MMPs was affected by various inhibitors of signaling pathways. In particular, expression of MMP-3 was slightly recovered by cRAF and PLC signaling pathway inhibitors. These data suggests a possible role of MMPs in macrophages during TG-induced atherosclerosis.
Key Words: Triglyceride, Matrix metalloprotease, THP-1 macrophages, Atherosclerosis
INTRODUCTION
Matrix metalloproteinases (MMPs) are zinc-dependent proteases that are synthesized in a variety of cells including monocytes, macrophages, endothelial cells, and smooth muscle cells (Schafers et al., 2010). MMPs are involved in a myriad of processes including blastogenesis, osteogenesis, wound healing, and angiogenesis. One prominent physio- logical role of MMP is degradation of the extracellular
matrix. However, abnormal overexpression or inappropriate MMP activation results in destruction of connective tissues resulting in a variety of inflammatory disorders (Amalinei et al., 2010). Moreover, MMPs are associated with cell proliferation, migration, and apoptosis. In immune cells, MMPs are involved in modulation of cytokines, cytokine receptors, or growth factors thereby regulating the immune response (Hojilla et al., 2003; Parks et al., 2004).
Currently, 23 kinds of MMPs are found in the human genome and they are categorized into five groups based on their substrate specificity and protein-domain-structure:
collagenases (MMP-1, MMP-8, MMP-13), gelatinases (MMP-2, MMP-9), stromelysins (MMP-3, MMP-10), matrilysins (MMP-7, MMP-26), and other MMPs (MMP- 12, MMP-19, MMP-20, MMP-27) (Nagase et al., 2006;
Fanjul-Fernández et al., 2010; Rivera et al., 2010). According to a recent report, MMPs expressed by monocytes and macrophages in particular are strongly associated with the development of atherosclerotic disorders (Newby, 2008).
Original Article
*Received: February 14, 2013 / Revised: March 15, 2013 Accepted: March 17, 2013
#These authors contributed equally to this work.
†Corresponding author: Ki-Jong Rhee. Department of Biomedical Laboratory Science, College of Health Sciences, Yonsei University, Wonju 220-710, Korea.
Tel: +82-33-760-2445, e-mail: [email protected]
†Corresponding author: Tae Ue Kim. Department of Biomedical Laboratory Science, College of Health Sciences, Yonsei University, Wonju, 220-710, Korea.
Tel: +82-33-760-2424, e-mail: [email protected]
○CThe Korean Society for Biomedical Laboratory Sciences. All rights reserved.
Human monocytes can express MMP-8, MMP-11, MMP- 17, MMP-23, and MMP-25 (Bar-Or et al., 2003). The MMP expression profile in monocytes changes depending on the cell cultivation time in vitro, stimulation from the extracellular matrix and influence from other cells (Amorino and Hoover, 1998; Matias-Roman et al., 2005). Many MMPs are also expressed in macrophages but the MMP expression profile is different from that of monocytes (Newby, 2008). Furthermore, studies using monocyte-like cell lines (eg, THP-1 and U937) showed that phorbol 12- myristate 13-acetate (PMA) treatment induced cell differ- entiation into macrophage-like cells which also dramatically changed the MMP expression profile (Amorino and Hoover, 1998; Newby, 2008). Collectively, these results all show that MMP expression by monocytes/macrophages differ based on the environmental cues, state of differentiation, and treatment by exogenous factors.
Atherosclerosis is a chronic inflammatory disease of arteries. This disease is mainly induced by oxidized low density lipoprotein (oxi-LDL) in the blood vessel and according to recent reports, elevated blood triglyceride (TG) levels is associated with the development of athero- sclerosis (Adams et al., 2006; Stalenhoef and de Graaf, 2008; Kannel and Vasan, 2009). The development of atherosclerosis is currently described by the reaction to injury hypothesis (Ross, 1993; Miyagawa and Hanafusa, 1999; Souza et al., 2009). In brief, permeability of the endothelial cell is increased due to injury. As a result, blood LDL passes through the endothelium and accumulates in the vessel tunica intima which then forms oxi-LDL (Shimokawa, 1999). Next, monocytes move to the tunica intima and differentiate into macrophages. The differentiated macro- phages ingest accumulated oxi-LDL in the tunica intima and become foam cells (Newby, 2008). In time, foam cells undergo apoptotic cell death and they are in turn removed by adjacent macrophages. Dead cells which are not removed by macrophages elicit necrosis of nearby cells which promote inflammatory reactions and finally atherosclerotic plaques (Thorp and Tabas, 2009; Tabas, 2009). Consistent with this hypothesis, TG promotes foam cell formation (Stalenhoef et al., 2008) which then increases the in- flammatory reaction of tissues due to necrosis rather than
apoptosis (Ley et al., 2011). Statistical analysis suggests that the risk of atherosclerosis is 2.6 fold higher in patients when blood TG levels are greater than 160 mg/dl. Moreover, this risk further increases up to 6 fold when the blood TG levels exceed 200 mg/dl and LDL/HDL (high density lipoprotein) ratio is higher than 5 (Kannel and Vasan, 2009;
Valdivielso et al., 2009). Although, the role of TG in atherogenesis is under investigation (Geurian et al., 1992), the precise mechanism has yet to be elucidated. In addition to the phagocytic role of macrophages, recruitment and accumulation of macrophages as well as smooth muscle cells are also important to the development of atherosclerosis (Johnson, 2007). In this context, the role of MMPs in breakdown of extracellular matrix can be considered a critical factor in cell migration and recruitment (Armstrong et al., 2007).
Previously, our laboratory examined the expression of MMPs during differentiation of monocytes into macro- phages and the signaling transduction pathways involved MMP expression (Lim J et al., 2012). In this study, we investigated the changes in the MMP expression profile of macrophages upon ingestion of TG in order to elucidate the role of TG in inflammatory diseases including athero- sclerosis. In addition, we investigated the impact of signal transduction pathways on MMP expression. These results should provide reference data for researchers regarding the role of TG on macrophages.
MATERIALS AND METHODS Reagents
Phorbol 12-myristate 13-acetate (PMA) was purchased from Sigma-Aldrich (St. Louis, MO, USA). TG was pro- vided by B. Braun Melsungen AG (Melsungen, Germany).
TRIzol for RNA isolation were obtained from Invitrogen (Carlsbad, CA, USA). Specific inhibitors for PKC (RO-31- 7549), MEK1 (PD98059), PI3-K (LY294002), p38 MAPK (SB203580) and cRAF kinase (GW5074) were purchased from Calbiochem (Darmstadt, Germany). Specific inhibitor of NF-κB (BAY11-7085) was purchased from Enzo Life Sciences (New York, NY, USA). Specific inhibitor of PLC (U73122) was purchased from Cayman Chemical (Ann
Arbor, MI, USA). Dimethyl sulfoxide (DMSO) was obtained from Sigma-Aldrich.
Cell culture and TG treatment
The human monocytoid THP-1 leukemia cell line was cultured in RPMI medium 1640 supplemented with 2 mM L-glutamine, 100 U/ml penicillin, 100 μg/ml streptomycin and 10% fetal bovine serum (FBS) at 37℃ under 5% CO2. THP-1 cells were seeded at a density of 1 × 106 cells/well in 6-well plates and treated with PMA (200 nM) for 48 hr to differentiate monocytes into macrophages. Differentiated macrophages were then treated with TG (1.0 mg/ml) and PMA (200 nM) for an additional 24 hr. For TG dose- dependent experiments, differentiated macrophages were simultaneously treated with different doses of TG (0, 0.1, 0.2, 0.5, 1.0, 2.0 mg/ml) and PMA (200 nM) for 24 hr. For time course experiments, differentiated macrophages were treated with TG (1.0 mg/ml) and PMA (200 nM) for the indicated time periods (0, 3, 6, 12, 18, 24 hr). To identify signaling transduction pathway(s) involved in MMP expression, THP-1 cells were treated with PMA (200 nM) for 48 hr and then the differentiated macrophages were treated with PMA (200 nM), TG (1.0 mg/ml) and chemical inhibitors for an additional 24 hr. A control group with no inhibitor treatment was included as a control. The doses of the chemical inhibitors used to identify the signaling trans- duction pathways are as follows: RO-31-7549 (50 nM), PD98059 (50 μM), BAY11-7085 (10 μM), LY294002 (10 μM), SB203580 (20 μM), GW5074 (0.5 μM) and U73122 (50 ng/ml). All inhibitors were dissolved as stock solutions in DMSO as stored at -20℃ prior to usage.
mRNA extraction and semi-quantitative reverse transcriptase-PCR (RT-PCR)
Total RNA was extracted from cultured cells by using TRIzol Reagent according to the manufacturer's instructions.
cDNA was synthesized by reverse transcription with 2 μg total RNA, 0.25 μg of random hexamer (Invitrogen) and 200 units of Murine Molony Leukemia Virus Reverse Transcriptase (MMLV-RT; Invitrogen) for 10 min at 25℃, 50 min at 37℃ and 15 min at 70℃. Diluted reverse- transcribed cDNA was used for subsequent PCR amplifi-
cation. PCR amplification was performed by using Prime Taq premix PCR kit (Genet Bio, Chungnam, Korea) and a thermocycler (Applied Biosystems, New York, NY, USA) for 25~40 cycles using MMP specific primers for MMP-1, MMP-2, MMP-3, MMP-7, MMP-8, MMP-9 and MMP-10.
The levels of gene expression were normalized against GAP-DH. PCR products were electrophoresed on 2.0%
(w/v) agarose gels containing 0.5 μg/ml ethidium bromide and the PCR product size compared to a 100 bp DNA ladder marker (Intron, Gyeonggi, Korea). Gel images were taken using Gel-Doc (Bio-Rad, Hercules, CA, USA). The intensity of each band amplified by RT-PCR was analyzed using Image Lab (version 2.0, Bio-Rad) and normalized to GAP-DH in corresponding samples.
Statistical analysis
P values were calculated using Student's t-test. Values are shown as mean and standard error of mean (SEM). Data were collected from at least three independent experiments.
RESULTS
TG treatment induces down-regulation of MMP-3, MMP-7, MMP-8, and MMP-9 expression in PMA- differentiated THP-1 macrophage cells
In a previous report, we found that expression of several MMPs increased when monocytes were differentiated into macrophages by PMA treatment (Lim J et al., 2012). We wondered whether exposure to TG by the PMA-induced differentiated cells resulted in further changes in MMP expression. Therefore, THP-1 cells were differentiated into macrophages by PMA treatment for 48 hr and then subsequently treated with TG for an additional 24 hr. MMP expression was determined by RT-PCR analysis of select MMPs. We found that compared to non-treated cells, TG treated THP-1 macrophages exhibited a decrease in expression of MMP-3, MMP-7, MMP-8, and MMP-9 (Fig.
1). In contrast, there was no change in expression of MMP- 1, MMP-2, and MMP-10 after TG treatment compared to non-treated controls. The decrease in MMP expression did not correlate with any certain class. These results demon- strate that TG treatment down-regulated expression of
select MMPs.
Dose-dependent effect of TG on MMP expression in PMA-differentiated THP-1 macrophages
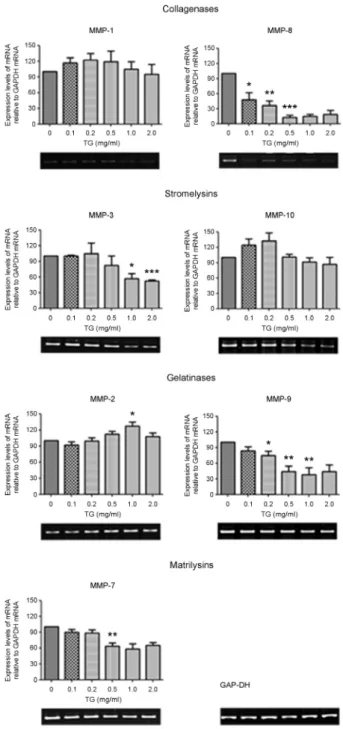
Next, we investigated the effect of TG dosage on MMP expression in THP-1 macrophages. PMA-differentiated THP-1 macrophages were treated with different doses of TG for 24 hr and then MMP expression examined by RT-PCR. We found that the mRNA levels of MMP-3, MMP-7, MMP-8, and MMP-9 decreased in a TG-dose- dependent manner (Fig. 2). In particular, MMP-8 expression was dramatically decreased even with 0.1 mg/ml of TG treatment. Expression of MMP-3, MMP-7, and MMP-9 decreased beginning at 0.5 mg/ml TG treatment. In contrast, expression of MMP-1, MMP-2, and MMP-10 either showed no change in expression or was transiently increased at certain concentrations of TG treatment. These results suggest that the expression of MMP-3, MMP-7, MMP-8, and MMP-9 is decreased in a TG-dose-dependent manner in THP-1 macrophages.
Time-dependent effect of TG on MMP expression in PMA-differentiated THP-1 macrophage
We next examined whether the kinetics of MMP
Fig. 1. TG treatment induces downregulation of MMP expression in PMA-differentiated THP-1 macrophages. THP-1 cells were differentiated into macrophages by treating the cells with PMA (200 nM) for 48 h. The differentiated cells were incubated with TG (1.0 mg/ml) or without TG for an additional 24 h. cDNA was prepared from total RNA extracts and subjected to PCR to amplify MMPs. The PCR products were resolved on a 2% agarose gel. GAP-DH was used as an internal control.
Fig. 2. Dose-dependent effect of TG on MMP expression in PMA-differentiated THP-1 macrophages. Differentiated THP-1 macrophages were treated with indicated concentrations of TG (0, 0.1, 0.2, 0.5, 1.0, 2.0 mg/ml) for 24 hr. cDNA was prepared from extracted total RNA and subjected to PCR to amplify MMPs. The PCR products were analyzed on a 2% agarose gel and GAP-DH was used as an internal control. Densitometric analysis was per- formed on data from three independent experiments and the data expressed as the mean ± SEM with the expression levels of MMPs normalized to GAP-DH expression levels. The expression level MMPs relative to GAP-DH levels in differentiated THP-1 macro- phage without TG treatment was set as 100. P-values were deter- mined by the Student's t-test. *P<0.05, **P<0.01, ***P<0.001.
expression in THP-1 macrophages after TG treatment.
THP-1 cells were differentiated into macrophages by after 48 hr of PMA treatment and then MMP expression examined at different time intervals post-TG treatment by RT-PCR analysis. We found that the expression of MMP-3, MMP-7, MMP-8, MMP-9 and MMP-10 decreased in a time-dependent manner (Fig. 3). In contrast, MMP-1 expression remained unchanged during TG treatment.
MMP-2 expression transiently increased during TG treat- ment but decreased to pretreatment levels by 24 hr. These results show that the expression of MMP-3, MMP-7, MMP- 8, MMP-9 and MMP-10 decreased after TG treatment in a time-dependent manner.
Identification of signal transduction pathways involved in TG-induced changes in MMP expression in PMA- differentiated THP-1 macrophages.
Next, we determined the signaling pathways involved in TG-induced MMP down-regulation. PMA-differentiated
THP-1 macrophages were treated with both TG (1.0 mg/
ml) and signal transduction chemical inhibitors for 24 h and MMP expression examined by RT-PCR. The chemical inhibitors used were as follows: PKC (RO-31-7549), MEK1 (PD98059), PI3-K (LY294002), p38 MAPK (SB203580), cRAF kinase (GW5074), NF-κB (BAY11-7085) and PLC (U73122). Our results show that of the four MMPs that were down-regulated by TG treatment, only, MMP-3 down- regulation was inhibited by the cRAF kinase inhibitor (GW5074) and the PLC inhibitor (U73122) (Fig. 4). In fact, the cRAF kinase inhibitor and the PLC inhibitor treatment resulted in a trend toward increased expression of MMP-3 well beyond the levels TG-treated cells. This result suggests that TG-induced down-regulation of MMP-3 expression is may be acting through the cRAF kinase and the PLC pathways. We also observed a trend toward increased levels of MMP-10 and MMP-2 expression after treatment with the cRAF kinase inhibitor and MEK1 inhibitor, respectively.
However, these changes are likely not due to TG treatment Fig. 3. Time-dependent effect of TG on MMP expression in PMA-differentiated THP-1 macrophage. Differentiated THP-1 macrophages were treated with TG (1.0 mg/
ml) for the indicated times (0, 3, 6, 12, 18, 24 hr). Total RNA was extracted and cDNA was prepared. PCR analysis was performed using MMP-specific primers. The PCR products were resolved on a 2% agarose gel. GAP-DH was used as an internal control. Densitometric analysis was performed on data from three in- dependent experiments and the data expressed as the mean ± SEM with the expression levels of MMPs normalized to GAP-DH expression levels. The expression level MMPs relative to GAP-DH levels in differentiated THP-1 macrophage without TG treatment was set as 100. P-values were determined by the Student's t-test. *P<0.05, **P<0.01, ***P<0.001.
but rather changes in constitutive MMP expression affected by the chemical inhibitors.
DISCUSSION
It is well documented that macrophages alter their MMP expression profile in response to various external stimuli such as bacterial lipopolysaccharide or inflammatory cytokines.
In addition to these exogenous factors, alteration in MMP expression occurs when monocytes differentiate into macro- phages upon migration from the blood to tissues (Newby, 2008). Therefore, changes in the cellular environment can also trigger changes in MMP expression. It has been pro- posed that a subset of MMPs exhibit enhanced expression or increased protein activation when monocytes contact the extracellular matrix. The types of MMPs expressed during these processes would greatly impact the physiologic and/or the disease processes. In this study, we evaluated changes in the MMP expression profile by macrophages in response to TG treatment. We found that of the MMPs examined the expression of MMP-3, MMP-7, MMP-8, and MMP-9 were decreased in PMA-differentiated macrophages following TG treatment in a dose- and time-dependent manner. In contrast, MMP-1, MMP-2 and MMP-10 expression were not greatly affected by TG treatment. These results suggest that the change in MMP expression by TG shows a certain degree of specificity implying activation of distinct signaling pathways. Consistent with the hypothesis, we found that inhibition of select signal transduction pathways alleviated the TG-induced down-regulation of MMPs especially MMP-3.
Our results suggest the possibility that during athero- sclerosis, accumulation of TG in macrophages could lead to reduction of MMP expression. This decrease in MMP expression may have several effects one of which is prolonged retention of the macrophages in the tunica intima due to lack of proteases to allow movement of macrophages.
Thus accumulation of these macrophages may eventually lead to further formation of foam cells and promotion of atherosclerotic plaques. The strong association of MMPs and inflammatory diseases strongly argue for a causal role of MMPs in these diseases. Therefore, the molecular and cellular regulatory processes involved in MMP expression and activation are critical for understanding these patho- Fig. 4. Identification of signal transduction pathways in-
volved in TG-induced changes in MMP expression in PMA- differentiated THP-1 macrophages. Differentiated THP-1 macro- phages were treated with TG (1.0 mg/ml) and specific chemical inhibitors for 24 hr. cDNA was prepared from total RNA extracts and subjected to PCR to amplify MMPs. The PCR products were resolved on a 2% agarose gel. GAP-DH was used as an internal control. Densitometric analysis was performed on data from three independent experiments and the data expressed as the mean ± SEM with the expression levels of MMPs normalized to GAP-DH expression levels. The expression level MMPs relative to GAP-DH levels in differentiated THP-1 macrophage without TG treatment was set as 100. Control, untreated cells. TG only, TG treatment with no inhibitors. P-values were determined by the Student's t-test. *P<0.05, **P<0.01.
NF-κb
NF-κb NF-κB NF-κB NF-κB
NF-κB NF-κB NF-κB
NF-κB
logical states. In regards to atherosclerosis, the impact of TG on MMP expression in macrophages would greatly benefit researchers in understanding the basic mechanism of MMP regulation and utilization for clinical research.
REFERENCES
Adams LD, Geary RL, Li J, Rossini A, Schwartz SM. Expression profiling identifies smooth muscle cell diversity within human intima and plaque fibrous cap: loss of RGS5 distinguishes the cap. Arterioscler Thromb Vasc Biol. 2006. 26: 319-325.
Amalinei C, Caruntu ID, Giusca SE, Balan RA. Matrix metalloproteinases involvement in pathologic conditions.
Rom J Morphol Embryol. 2010. 51: 215-228.
Amorino GP, Hoover RL. Interactions of monocytic cells with human endothelial cells stimulate monocytic metalloproteinase production. Am J Pathol. 1998. 152: 199-207.
Armstrong C, Abilleira S, Sitzer M, Markus HS, Bevan S.
Polymorphisms in MMP family and TIMP genes and carotid artery intima-media thickness. Stroke. 2007. 38: 2895-2899.
Bar-Or A, Nuttall RK, Duddy M, Alter A, Kim HJ, Ifergan I, Pennington CJ, Bourgoin P, Edwards DR, Yong VW. Analyses of all matrix metalloproteinase members in leukocytes emphasize monocytes as major inflammatory mediators in multiple sclerosis. Brain. 2003. 126: 2738-2749.
Fanjul-Fernández M, Folgueras AR, Cabrera S, López-Otín C.
Matrix metalloproteinases: evolution, gene regulation and functional analysis in mouse models. Biochim Biophys Acta.
2010. 1803: 3-19.
Geurian K, Pinson JB, Weart CW. The triglyceride connection in atherosclerosis. Ann Pharmacother. 1992. 26: 1109-1117.
Hojilla CV, Mohammed FF, Khokha R. Matrix metalloproteinases and their tissue inhibitors direct cell fate during cancer development. Br J Cancer. 2003. 89: 1817-1821.
Johnson JL. Matrix metalloproteinases: influence on smooth muscle cells and atherosclerotic plaque stability. Expert Rev Cardiovasc Ther. 2007. 5: 265-282.
Kannel WB, Vasan RS. Triglycerides as vascular risk factors: new epidemiologic insights. Curr Opin Cardiol. 2009. 24: 345-350.
Ley K, Miller YI, Hedrick CC. Monocyte and macrophage dynamics during atherogenesis. Arterioscler Thromb Vasc Biol. 2011. 31: 1506-1516.
Lim J, Cho Y, Lee DH, Jung BC, Kang HS, Kim T-J, Rhee K-J, Kim TU, Kim YS. Upregulation of MMP is mediated by
MEK1 activation during differentiation of monocyte into macrophage. J Exp Biomed Sci. 2012. 18: 104-111.
Matias-Roman S, Galvez BG, Genis L, Yanez-Mo M, de la Rosa G, Sanchez-Mateos P, Sanchez-Madrid F, Arroyo AG. Membrane type 1-matrix metalloproteinase is involved in migration of human monocytes and is regulated through their interaction with fibronectin or endothelium. Blood. 2005. 105: 3956-3964.
Miyagawa J, Hanafusa T. Mechanism of atherosclerosis in diabetes:
altered cytokine network in the vascular wall. Nihon Rinsho.
1999. 57: 601-606.
Nagase H, Visse R, Murphy G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc Res. 2006. 69:
562-573.
Newby AC. Metalloproteinase expression in monocytes and macrophages and its relationship to atherosclerotic plaque instability. Arterioscler Thromb Vasc Biol. 2008. 28: 2108 -2114.
Parks WC, Wilson CL, Lopez-Boado YS. Matrix metallopro- teinases as modulators of nflammation and innate immunity.
Nat Rev Immunol. 2004. 4: 617-629.
Rivera S, Khrestchatisky M, Kaczmarek L, Rosenberg GA, Jaworski DM. Metzincin proteases and their inhibitors: foes or friends in nervous system physiology? J Neuroscience.
2010. 30: 15337-15357.
Ross R. Atherosclerosis: current understanding of mechanisms and future strategies in therapy. Transplant Proc. 1993. 25:
2041-2043.
Schafers M, Schober O, Hermann S. Matrix-metalloproteinases as imaging targets for inflammatory activity in atherosclerotic plaques. J Nucl Med. 2010. 51: 663-666.
Shimokawa H. Primary endothelial dysfunction: atherosclerosis. J Mol Cell Cardiol. 1999. 31: 23-37.
Souza HP, Frediani D, Cobra AL, Moretti AI, Jurado MC, Fernandes TR, Cardounel AJ, Zweier JL, Tostes RC.
Angiotensin II modulates CD40 expression in vascular smooth muscle cells. Clin Sci (Lond). 2009. 116: 423-431.
Stalenhoef AF, de Graaf J. Association of fasting and nonfasting serum triglycerides with cardiovascular disease and the role of remnant-like lipoproteins and small dense LDL. Curr Opin Lipidol. 2008. 19: 355-361.
Tabas I. Macrophage apoptosis in atherosclerosis: consequences on plaque progression and the role of endoplasmic reticulum stress. Antioxid Redox Signal. 2009. 11: 2333-2339.
Thorp E, Tabas I. Mechanisms and consequences of efferocytosis
in advanced atherosclerosis. J Leukoc Biol. 2009. 86: 1089 -1095.
Valdivielso P, Sanchez-Chaparro MA, Calvo-Bonacho E, Cabrera- Sierra M, Sainz-Gutierrez JC, Fernandez-Labandera C, Fernandez-Meseguer A, Quevedo-Aguado L, Moraga MR,
Galvez-Moraleda A, Gonzalez-Quintela A, Roman-Garcia J.
Association of moderate and severe hypertriglyceridemia with obesity, diabetes mellitus and vascular disease in the Spanish working population: results of the ICARIA study.
Atherosclerosis. 2009. 207: 573-578.