Vol. 14, No. 1, March, 2007
<접수일:2006년 12월 4일, 심사통과일:2006년 12월 6일>
※통신저자:박 성 환
서울시 서초구 반포동 505번지
가톨릭대학교 의과학 연구원 류마티스연구센터
Tel:02) 590- 2712, Fax:02) 599- 3589, E- mail:[email protected]
과학기술부/한국과학재단 우수연구센터 육성사업의 지원으로 수행되었음(R11- 2002- 098- 05003- 0).
Estrogen이 전신홍반루푸스 환자 B세포의 DNA Methylation에 미치는 영향
가톨릭대학교 의과학연구원 류마티스연구센터
박미경·박성환·곽승기·조미라·김호연
= Abstract =
The Effect of Estrogen on the DNA Methylation of B Cells in Patients with SLE
Mi-Kyung Park, B.S., Sung-Hwan Park, M.D.,
Seung-Ki Kwok, M.D., Mi-La Cho, Ph.D., Ho-Youn Kim, M.D.
The Rheumatism Research Center (RhRC), Catholic Research Institute of Medical Science, The Catholic University of Korea, Seoul, Korea
Objective: Epigenetics is an important, alternative mechanism of gene regulation that is independent of the nucleotide sequences of DNA. We investigated mRNA levels for DNA methyltransferase-1 (DNMT-1), and the effect of estrogen on the expression of DNMT-1 level in T cells and B cells from patients with systemic lupus erythematosus (SLE) and healthy subjects, and assessed the possible etiological role of DNA methylation in the pathogenesis of SLE.
Methods: mRNA levels for DNMT-1 in CD4+ T cells and CD19+ B cells from 37 patients with SLE and 12 healthy controls were examined using RT-PCR. We used specific primer for DNMT-1 and β actin, The effect of estrogen on the DNA methylation was measured by the mRNA level of DNMT-1 CD4+ T cells and CD19+ B cells treated with 100 nM of 17β-estradiol for 72 hour.
Results: The levels of DNMT-1 mRNA were significantly lower in CD4+ T cells and CD19+
B cells from SLE patients compared with healthy controls. We observed the suppression of the levels of DNMT-1 mRNA by stimulated with estrogen in patients with SLE patients, especially
서 론
전신홍반루푸스(systemic lupus erythematosus, 루푸 스)는 자가 항원에 대한 면역 반응으로 다양한 자가 항체를 나타내는 자가면역질환으로, 피부, 신경계, 폐, 관절, 혈관계, 신장 등의 조직에 염증을 일으킨 다. 루푸스의 원인은 불명확하지만, 비정상적인 면역 반응이 그 원인으로 생각되고 있으며 이러한 면역 반응의 이상에 관여하는 것으로 여러 종류의 유전자 와 감염, 자외선 등의 환경적 요인이 제시되고 있다 (1-3).
최근 DNA 염기 서열상 변화없이 유전자 조절에 변화의 기전을 밝히는 후생적 변화(epigenetics) 연구 가 활발해지고 있으며 (4), 메틸화의 변화로 인한 DNA 메틸화(DNA methylation)나 저메틸화(hypometh- ylation)는 암, 면역 결핍 질환, 자가면역질환의 발병 에 관계하는 것이 알려지고 있다. DNA의 조절 서열 (regulatory sequence)의 시토신 메틸화는 유전자의 전 사 불활성과 관련된 후생적인 메커니즘(epogeneitic mechanism)인 반면, DNA 저메틸화는 전사 활성의 원인이 된다 (5,6). 포유류의 DNA 메틸화에서 DNA methyltransferase-1 (DNMT-1)은 DNA methyltransfe- rase의 3가지 families (DNMT1, DNMT2, DNMT3a와 DNM-T3b) 중에서 genomic 메틸화 패턴의 결정과 유 전자 억제의 역할에 가장 중요한 효소이다 (7,8).
자가면역질환인 루푸스에서도 후생적인 DNA 변화 에 대한 연구가 진행 중이며 T세포 DNA 메틸화의 이상이 루푸스의 발병에 관여한다는 최근 보고가 있 다. 이중 T세포 DNA 저메틸화와 같은 조절 DNA 서 열(regulatory DNA sequence)상의 후생적인 이상이 루푸스 내 면역체계 변화의 원인이 될 수 있음이 주
장되고 있으며, CD4+ T세포의 DNA 저메틸화는 LFA-1, CD70, perforin 등의 발현을 통해 약물유발 루푸스와 SLE 발병에 관여한다는 연구 결과가 있다 (9-14).
루푸스는 강한 여성 성별 편중(sex bias)을 보여 90% 내외의 환자가 여자에서 발생하여 대표적인 성 호르몬인 에스트로겐이 루푸스의 발병 감수성에 중 요한 인자로 생각되고 있다 (15). 에스트로겐은 생리 학적인 용량에서도 사이토카인의 생성이나 자가 항 체를 생산하는 등의 면역 반응을 촉진시킬 수 있는 물질로 여겨지며, 면역 세포의 활성화와 사멸에 관 여하여 루푸스 발병에 있어서 성별 편중을 나타나게 하는데 중요한 인자로 생각되어진다 (16,17-19). 에스 트로겐이 루푸스 발병에 미치는 연구는 아직 그 기 전이 완전히 밝혀지지는 않았지만 최근 에스트로겐 이 정상인의 말초 혈액 내에서 인간내생레트로바이 러스(human endogenous retroviruses, HERV)와 같은 내 부(endogenous) 자가항원의 발현을 후생적인 메커니즘 을 통해 증가시킨다는 연구가 발표되고 있다 (20-22).
하지만 에스트로겐과 DNA 메틸화와의 상관성은 종양과 혈관 질환에서 일부 보고되고 있으며, 루푸 스 발병에 관여할 것으로 알려지고 있는 T세포 및 B세포의 후생적인 발현에 어떤 영향을 미치는지에 대해서는 조사가 부족하다.
본 연구에서는 루푸스의 병인에 중요한 T세포 및 B세포의 후생학적 변화를 조사하고, 특히 에스트로 겐이 DNA 메틸화 변화의 요인이 될 수 있는지를 조사하여 루푸스의 질병 활성도의 지표와 항DNA 항체의 역가와의 관련성을 분석함으로써 이러한 후 생적인 변화와 루푸스 병인과의 상관성을 조사하고 자 한다.
in CD19+B cells. DNA hypomethylation of B cells was tend to be correlated with the level of anti-ds DNA antibody without statistical significance (r=-0.43, p=0.3).
Conclusion: Our observations suggest that suppression of DNMT-1 by estrogen in B cells from patients with SLE might be related to the pathogenesis of SLE. Epigenetic studies may provide clues for developing new treatment strategies of SLE.
Key Words: Systemic lupus erythematosus, DNA methyltransferase-1, DNA methylation, Estrogen
대상 및 방법 1. 대상
1997년에 개정된 미국류마티스학회(American Col- lege of Rheumatology, ACR)의 루푸스 분류 기준을 만족시키는 37명의 환자(남자 2명, 평균 연령 35.5 세)와, 의료 종사자 중 최근의 건강검진에서 정상으 로 판명된 자원자 중 성별과 나이가 맞춰진 12명(남 자 1명, 평균 연령 33.2세)을 대상으로 하였다. 본 연구는 강남성모병원 임상연구관리 규정과 헬싱키 선언을 준수하여 시행하였다. 환자 군에서는 채혈 당시에 루푸스 질병 활성도(SLEDAI)를 측정하였고, 혈청 내 IgE 농도(정상 150 IU/mL 이하), 보체 활성 도, 항dsDNA 항체 역가 등을 측정하였고, 복용 약물 의 종류를 기록하였다.
2. 세포 분리
말초혈액 단핵세포는 헤파린을 처리한 주사기로 혈액을 Phoshpate-buffered saline (PBS, Gibco BRL.
Carlsbad, CA)와 1:1로 섞어 Ficoll (Amercham Bio- sciences, burkinghamshire, England)과 1:4의 비율로 Ficoll층에 섞이지 않게 조심스럽게 50 mL tube에 천 천히 띄운 다음 혈액을 20oC의 2,000 rpm에서 30분 간 원심 분리하였다. Buffy coat층만을 따서 새 용기 에 옮긴 후 PBS로 세척하였다. 말초 혈액 단핵세포 의 CD19+ B세포와 CD4+ T세포의 분리는 MACS bead (Miltenyi Bitec, Bergisch Gladbach, Germany)를 이용해서 분리하였다. 먼저 CD19+ B세포를 분리하 기 위해 PBS 세척된 세포에 CD19 bead를 넣고 4oC 에서 15분간 반응시킨 다음 MACs buffer (1% BSA, 2 mM EDTA in PBS, pH 7.4)로 세척한 후 제조업체 의 설명서에 따라 AutoMACS (Miltenyi)를 이용하여 양성 분획으로 CD19+ B세포를 분리하였다. CD4+
T세포를 분리하기 위해 음성 분획의 CD19- 세포를 다시 한번 MACs buffer로 세척한 후에 anti-human CD4 biotin cocktail (Miltenyi)와 4oC에서 15분간 반응 시킨 후 anti-human CD4 biotin Microbead (Miltenyi) 와 4oC에서 15분간 다시 반응시켰다. 반응시킨 세포 를 MACS buffer로 세척하고 AutoMACS (Miltenyi) 를 이용하여 음성 분획으로 CD4+ T세포를 분리하
였다. 분리된 세포는 PBS 세척 후 insulin transferrin selenium A (ITS-A, Gibco BRL)이 포함된 serum free RPMI 1640 (Gibco BRL)세포 배양액에 재부유시켰다.
3. 세포 자극 및 배양
말초혈액 단핵세포의 T세포와 B세포 2.5×105/250 μL의 수의 세포를 ITSA가 포함된 serum free RPMI 1640 세포 배양액과 섞어 96 well plate (Iwaki Co., Tokyo, Japan)에 분주하고 17β-Estradiol (Sigma-Aldrich, St. Louis, MO, USA)을 100 nM로 처리하여 72시간 동안 37oC, 5% 배양기에서 배양하였다.
4. RNA 분리와 DNA methyltransferase-1 (DNMT-1) 의 RT-PCR
말초혈액 단핵세포의 T세포와 B세포 2.5×105/250 μL를 96 well plate에 부유한 후 17β-Estradiol로 처 리한 세포를 PBS로 세척하였다. 세척된 세포로부터 총 RNA를 RNAzolBTM (Tel.Test, Friendwood, TX)를 이용하여 추출하였다. 추출한 총 RNA를 주형으로 complementary DNA (cDNA)를 합성하기 위하여, 2 μg의 RNA에 0.5μg random primer (Takara, Shiga, Japan)와 70oC에서 5분 반응시킨 뒤 4oC에서 급냉시 켰다. 다음 10 mM dNTP mix (Invitrogen, Carlsbad, Califonia) 1μL, 역전사효소 M-MuLV (MBI Fermen- tas, Hanover, MD) 1 unit, 5× M-MuLV 희석용액 (MBI Fermentas) 4μL, RNase Inhibitor (MBI Fer- mentas) 0.5μL를 가하고 전체를 nuclease free water (Promega, Madison, WI) 20μL로 맞춘 뒤, 25oC에서 10분, 42oC에서 60분, 72oC에서 10분간 반응시켰다.
생성된 cDNA 산물을 이용하여 중합효소 연쇄반응 를 시행하였다. 즉, 25μL의 반응액 내에, cDNA 1 μL, 2.5 mM dNTP mix (Takara) 2μL, 10× Taq buf- fer (iNtRON Biotech, Seoul, Korea) 2.5μL, Taq 0.5 μL (iNtRON), 10 pmol DNMT-1 primer 2μL를 사용 하였고, 증폭을 위해 Dual-bay Thermal cycler system (MJ Research)를 사용하였다. 사용된 primer 서열은 β-actin (sense 5’-GGA CTT CGA GCA AGA GAT GG-3’, antisense 5’-TGT GTT GGC GTA CAG GTC TTT G-3’) DNMT-1 (sense 5’-TTC CG CTG AAC AAC CC-3’, antisense 5’-TGT CTC GCA CCG TGA TGGTCC G-3’) (21, 그림 1A)으로, β-actin은 조건은
변성 단계에서 94oC 30초, annealing 단계에서 60oC 30초, 신장 단계에서 72oC 30초간 반응을 25회 반복 하였으며 DNMT-1 증폭 조건은 변성 단계에서 94oC
30초, annealing 단계에서 62oC 45초, 신장 단계에서 72oC 1분간 반응을 30회 반복하였다. 음성 대조군으 로 추출한 cDNA 대신 증류수를 사용하여 시행한 PCR 반응에서 PCR 산물이 관찰되지 않도록 하여 PCR 오염이 없음을 확인하였다.
5. 통계
실험 결과는 평균±표준오차로 표현하였으며, SPSS 통계 프로그램(version 10.0)을 사용하였다. 정상인과 환자군의 평균 비교는 Mann-Whitney test를, DNMT-1 과 질병 활성도, 항체 역가 등과의 상관관계는 Spear- man 상관 계수를 이용하였으며, p값이 0.05 이하일 때 통계적으로 유의 하다고 분석하였다.
결 과
1. CD4+ T세포 및 CD19+ B세포 내 DNMT-1 mRNA 발현 수준
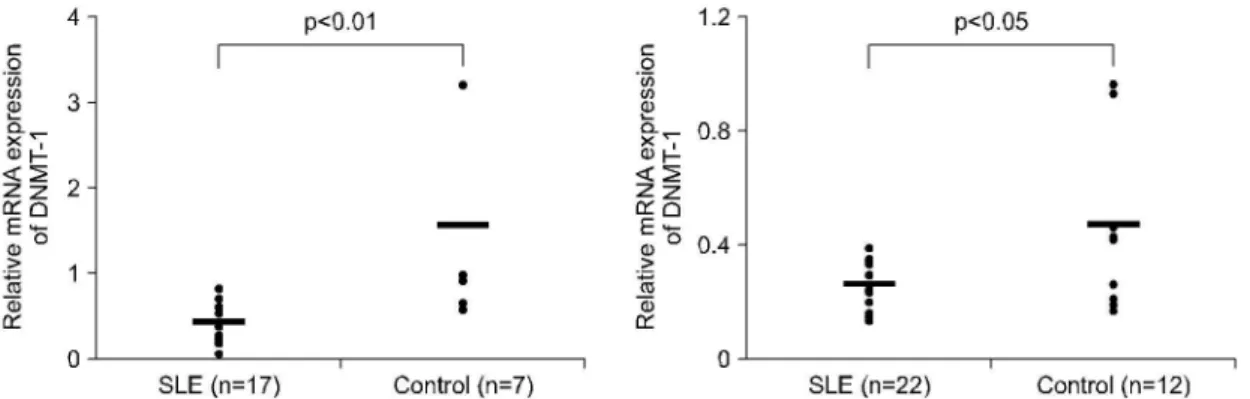
루푸스 환자와 정상인의 말초혈액 단핵세포로부터 분리한 직후의 CD4+ T세포와 CD19+ B세포의 DNMT-1 mRNA 발현 양상을 알아보고자 역전사 중 합효소 연쇄반응(RT-PCR)을 수행하였다. 그 결과 루 푸스 환자군의 CD4+ T세포에서 DNMT-1 mRNA 발 현이 정상 대조군에 비해 유의하게 감소되어 있었다 (그림 1B, 2A, mean±SD; 0.42±0.24 vs 1.56±1.17, p<0.01). 이러한 결과는 자극 없이 72시간 동안 오 직 세포 배양액과 함께 배양된(in vitro) CD4+ T세포
Fig. 2. The exprssion of DNMT-1 mRNA level on CD4 T cells of healthy controls and SLE patients. Freshly isolated CD4 T cells (2.5×105cells/well, left) and cultured CD4 T cells (2.5×105cells/well, right) for 72 hours in vitro from healthy controls and SLE patients were analyzed by RT-PCR as described in Fig. 1. β-actin was used as an internal control. The number of patients and control included is given below the x-axis.
Fig. 1. Structure and the exprssion of DNMT-1 mRNA level on CD4 T cell. (A) Structure of DNMT-1.
Numbers indicate the nucleotide numbers. (B) The exprssion of DNMT-1 mRNA level on CD4 T cell of healthy controls and SLE patients, Freshly isolated CD4 T cell (2.5×105 cells/well) and cultured CD4 T cell (2.5×105 cells/well) for 72 hours in vitro from healthy controls and SLE patients were analyzed by RT-PCR with specific primer of DNMT-1 and β-actin as described in Materials and Methods. β-actin was used as an internal control. The exprssion of DNMT-1 mRNA level on CD4 T cell visualized on 1.5%
agarose gel electrophoresis. Result are representa- tive of two or three experiment.
에서도 동일한 결과를 확인하였다(그림 2B, mean±
SD; 0.26±0.12 vs 0.47±0.31, p<0.05).
또한 자극 없이 72시간 동안 오직 세포 배양액과 함께 배양된 CD19+ B세포에서도 DNMT-1 mRNA
발현이 정상 대조군에 비해 유의하게 감소되어 있었 다(그림 3, mean±SD; 0.43±0.30 vs 1.02±0.54, p<
0.05).
2. CD4+ T세포 및 CD19+ B세포에서 estrogen 처리에 의한 DNMT-1 mRNA 발현 변화
루푸스 환자와 정상인의 말초 혈액 단핵세포로부 터 분리한 CD4+ T세포 및 CD19+ B세포를 에스트 로겐과 72시간 동안 처리 후 세포에서 RNA를 추출 하여 RT-PCR을 수행하였다. 루푸스 환자군에서 에스 트로겐을 처리한 CD19+ B세포의 DNMT-1 mRNA 발현 수준이 정상 대조군과 비교하였을 때 통계적으 로 유의하게 낮게 나타났다(그림 4A, mean±SD; 0.66
±0.44 vs 1.24±0.71, p<0.05), 에스트로겐을 처리한 CD4+ T세포의 DNMT-1 mRNA 발현 수준이 정상 대조군과 비교하였을 때 더 낮게 나타났지만 두 그 룹 간의 통계적인 유의한 차이는 보이지 않았다(그 림 4B, mean±SD; 0.71±0.37 vs 1.58±1.37, p>0.05).
3. 루푸스 환자의 임상 특징과 DNMT-1 mRNA 발 현과의 상관관계
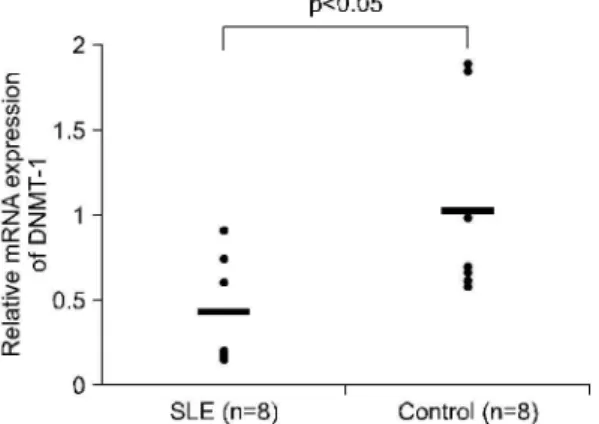
루푸스 환자의 CD4+ T세포 CD19+ B세포의 DNMT-1 mRNA 발현 정도와 루푸스의 질병 활성도, 항dsDNA 항체 역가, 혈청 IgE 농도, 복용 중인 스테 Fig. 3. The exprssion of DNMT-1 mRNA level on CD19+
B cell of healthy controls and SLE patients.
Cultured CD19+ B cell (2.5×105 cells/well) for 72 hours in vitro from healthy controls and SLE patients were analyzed by RT-PCR with specific primer of DNMT-1 and β-actin as described in Materials and Methods. β-actin was used as an internal control. The number of patients and control included is given below the x-axis.
Fig. 4. The exprssion of DNMT-1 mRNA level by 17 β- estradiol on CD19 B cell and CD4 T cell of healthy controls and SLE patients. CD19+ B cells (2.5×105cells/well, left) and CD4+ T cells (2.5×105cells/well, right) from healthy controls and SLE patients treated with 100 nM of 17β-estradiol for 72 hours. Cultured CD19+ B cell and CD4+ T cell were analyzed by RT-PCR with specific primer of DNMT-1 and β-actin as described in Materials and Methods. β-actin was used as an internal control. The number of patients and control included is given below the x-axis.
로이드 용량과의 상관관계를 조사하였다. 그 결과 CD19+ B세포의 DNMT-1 mRNA 발현 정도와 항 dsDNA 항체 역가와는 음의 상관관계 경향을 보였으 나 통계적 유의성에는 도달하지 못했다(그림 5, r=
-0.43, p=0.3). CD19+ B세포의 DNMT-1 mRNA 발 현 정도와 다른 임상 양상과는 유의한 상관관계는 관찰되지 않았으며, 또한 CD4+ T세포의 DNMT-1 mRNA 발현 정도와 임상 양상과는 유의한 상관관계 는 관찰되지 않았다(결과 제시하지 않음).
고 찰
DNA 염기 서열에 아무런 변화 없이 유전자 조절 에 변화를 나타내는 것을 후생적 변화라고 하며, 가 장 잘 알려져 있는 후생적 변화에 의한 유전자의 조 절은 DNA 메틸화와 히스톤의 조절에 의한 기작이 다. DNA 메틸화는 CpG 염기로 구성된 시토신의 5’
위치에서 주로 일어나며 CpG가 특히 많이 군집되어 있는 프로모터의 CpG island의 DNA 메틸화나 저메 틸화는 암, 면역결핍질환, 자가면역질환의 발병에 관 계하는 것이 알려지고 있다. 이러한 후생적 변화가 특정 질병의 발병에 미치는 영향에 대한 연구는 최 근 들어 활발히 이루어지고 있으며 종양억제 유전자 들의 과메틸화가 전사 조절에 영향을 미쳐 암의 악 성 변화의 원인이 될 수 있으며 특정 유전자의 저메 틸화 또는 과메틸화가 종양의 진행에 관여한다는 연
구 결과가 있다. 후생적 변화는 개인의 일생을 통해 서서히 축적될 수 있으며 이러한 변화들이 유전적으 로 동일한 일란성 쌍생아에서 질병의 발병 유무의 차이를 나타내는 환경적 요인을 설명하는 하나의 가 설로 제시되고 있다 (23,24).
대표적인 자가면역질환의 하나인 루푸스의 발병에 대한 후생적 연구는 아직까지는 미미하다. 루푸스의 가장 큰 특징 중 하나인 다양한 자가 항체의 존재는 루푸스 환자의 단핵구나 림프구의 세포사멸 속도가 증가하고 세포 사멸 동안 배출된 자가항원을 비정상 적으로 인식하는 면역 반응의 결과로 설명하고 있으 며 이러한 과정에 후생적 변화가 관여할 수 있음이 제시되고 있다. DNA 메틸화의 변화는 유전자의 전 사 활성을 조절하며 저메틸화는 전사 활성을 유도하 는 것으로 알려져 있고, 활성기에 있는 루푸스 환자 의 T세포에는 메틸화가 저하된 DNA가 다수 존재함 이 알려졌으며, 최근 연구에는 DNA의 메틸화억제를 설명해 줄 수 있는 DNMT의 활성도가 감소됨이 보 고되었다. 또한 대표적인 메틸화 억제제인 5-azacyti- dine을 쥐에 처리한 경우 DNA 저메틸화가 나타나면 서 루푸스가 유도되었으며 약제 유발 루푸스와 관련 된 procainamide는 경쟁적인 DNMT 억제제로 약물 노출로 인해 저메틸화된 T세포가 정상상태에서는 역치 이하인 자극에 대해 과반응하거나 자가면역 반 응을 초래하여 약제 유발 루푸스가 발병하는 가설이 제시되고 있으며 이러한 변화와 B세포, 대식세포와 의 상호작용에 대한 연구가 진행되고 있다 (5,6,12, 13,16,23,25). T세포의 메틸화의 변화가 어떤 기전으 로 루푸스를 유발할 수 있는지를 포함하여 후생적 변화가 어떤 기전으로 인해 발생에 기여하는지는 아 직까지 잘 알려져 있지 않다. 본 연구에서는 기존에 알려진 루푸스 환자의 T세포뿐 아니라 B세포에서도 메틸화가 저하되어 있음을 알게 되었으며, 이러한 변화는 72시간 시험관 배양 후에도 T세포, B세포 모 두 메틸화가 지속적으로 감소됨을 조사하였다. 이러 한 세포들끼리의 상호 작용을 통한 면역 반응의 조 절 이상이 루푸스 발병에 기여할 것으로 생각되며, 루푸스가 강한 여성 성별 편중을 보이는 것에 관여 한다고 알려진 성 호르몬인 에스트로겐이 T세포, B 세포의 메틸화에 어떤 영향을 미치는지 조사하였다.
17β-Estradiol을 처리한 T세포 및 B세포의 메틸화 Fig. 5. Correlation between DNMT-1 levels of CD19+
B cell and serum anti-ds DNA antibody levels in SLE patients (n=8).
가 정상인과 비교하여 루푸스 환자에서 더욱 많이 억제됨을 확인하였고 T세포에서는 통계적인 유의성 에 도달하지 못했지만 B세포에서는 통계적으로 유 의하게 억제됨을 알게 되었다. 최근의 항DNA 항체 형질전환 쥐를 이용한 연구는 에스트로겐이 고친화 성의 DNA 반응 B세포의 관용을 깨는 기전을 통해 루푸스를 유발할 수 있음이 보고되고 있으며, 자가 반응 B세포의 생존과 활성을 촉진하는 것으로 보고 되고 있다 (25). 따라서 루푸스 환자에서 B세포의 메틸화 억제는 이러한 자가반응을 촉진시키는 데 기 여하는 것을 알 수 있으며 항dsDNA 항체 역가와의 관계에서도 B세포의 메틸화가 억제될수록 항DNA 항체의 역가가 증가하는 경향을 보임을 확인하였으 나 적은 실험대상 환자로 인해 통계적 유의성을 관 찰하지는 못했다.
에스트로겐은 여러 사이토카인이나 항체의 생산과 같은 면역반응을 촉진시키는 것으로 알려져 있으며 이러한 기전에는 본 연구에서 관찰한 B세포의 메틸 화 억제도 기여할 것으로 추정된다. 또한 루푸스 환 자의 낮은 DNMT-1의 활성도는 자가항원뿐만 아니 라 외래 항원에 대한 과민반응과도 관련되어 있어 저메틸화가 항원에 대한 과민반응에 관련된 중요한 인자라 할 수 있다 (16,19-21). 하지만 이번 연구에서 는 루푸스 환자의 혈청 내 IgE 농도, 질병 활성도 정도, 보체 수치 등과 CD4+ T세포 및 CD19+ B세 포의 DNMT-1 mRNA 발현과의 상관관계를 본 결과 유의한 상관관계를 발견할 수는 없었다. 이는 대상 환자 수가 적어 정확한 결론을 내리기 어려우며, 이 에 앞으로 좀더 많은 환자를 대상으로 장기적인 연 구가 필요할 것으로 생각된다.
최근에는 이러한 후생적 변화가 질병의 발병과 진 행에 미치는 영향을 연구하고 이를 통해 후생적 변 화에 관한 약물을 개발하려는 시도가 진행되고 있 다. 본 연구를 통해 루푸스의 발병 및 활성도에 메 틸화의 변화가 관여할 수 있음을 확인할 수 있었으 며 이러한 후생적 변화가 발병 및 진행에 어떤 기전 으로 작용하는지의 연구를 통해 새로운 후생적 변화 에 대한 약물개발에 근거자료가 될 수 있을 것으로 기대된다 (24).
결론적으로 저자들은 루푸스 환자의 말초혈액 내 T세포와 B세포의 메틸화에 관여하는 메틸화효소의
활성이 저하되어 있음을 관찰하였고, 에스트로겐이 이러한 메틸화 억제에 한 요소로 작용함을 관찰하였 다. 이러한 후생적 변화가 루푸스의 발병 및 진행에 관여하는 기전의 연구를 통해 루푸스의 새로운 치료 제 개발에 응용될 수 있을 것으로 기대된다.
결 론
루푸스 환자의 T세포 및 B세포에서 DNA 저메틸 화를 관찰하였으며 에스트로겐이 루푸스 환자 B세 포의 DNA 저메틸화에 기여하는 것을 확인하였다.
DNA 저메틸화의 기작에 대한 연구는 루푸스의 발 병 및 치료 기전을 연구하는 데 중요할 것으로 생각 된다.
REFERENCES
1) Mills JA. Systemic lupus erythematosus. N Eng J Med 1994;330:1871-9.
2) Boumpas DT, Austin HA 3rd, Fessler BJ, Balow JE, Klippel JH, Lockshin MD. Systemic lupus erythe- matosus: emerging concepts. Part 1: renal, neuropsy- chiatric, cardiovascular, pulmonary, and hematologic disease. Ann Intern Med 1995;122:940-50.
3) Boumpas DT, Fessler BJ, Austin HA 3rd, Balow JE, Klippel JH, Lockshin MD. Systemic lupus erythe- matosus: emerging concepts. Part 2: dermatologic and joint disease, the antiphospholipid antibody syndrome, pregnancy and hormonal therapy, morbidity and mor- tality, and pathogenesis. Ann Intern Med 1995;123:
42-53.
4) Wolffe AP, Matzke MA. Epigenetics: regulation through repression. Science 1999;286:481-6.
5) Richardson B, Yung R. Role of DNA methylation in the regulation of cell function. J Lab Clin Med 1999;
134:333-40.
6) Nakao M. Epigenetics: interaction of DNA methylation and chromatin. Gene 2001;278:25-31.
7) Bestor TH. Activation of mammalian DNA methyl- transferase by cleavage of a Zn binding regulatory domain. EMBO J 1992;11:2611-7.
8) Okano M, Xie S, Li E. Cloning and characterization of a family of novel mammalian DNA (cytosine-5) methyltransferases. Nat Genet 1998;19:219-20.
9) Richardson B, Powers D, Hooper F, Yung RL,
O'Rourke K. Lymphocyte function-associated antigen 1 overexpression and T cell autoreactivity. Arthritis Rheum 1994;37:1363-72.
10) Yung R, Powers D, Johnson K, Amento E, Carr D, Laing T, et al. Richardson B. Mechanisms of drug- induced lupus. II. T cells overexpressing lymphocyte function-associated antigen 1 become autoreactive and cause a lupuslike disease in syngeneic mice. J Clin Invest 1996;97:2866-71.
11) Kaplan MJ, Deng C, Yang J, Richardson BC. DNA methylation in the regulation of T cell LFA-1 ex- pression. Immunol Invest 2000;29:411-25.
12) Lu Q, Wu A, Richardson BC. Demethylation of the same promoter sequence increases CD70 expression in lupus T cells and T cells treated with lupus-in- ducing drugs. J Immunol 2005;174:6212-9.
13) Oelke K, Lu Q, Richardson D, Wu A, Deng C, Hanash S, et al. Overexpression of CD70 and overstimulation of IgG synthesis by lupus T cells and T cells treated with DNA methylation inhibitors. Arthritis Rheum 2004;50:1850-60.
14) Kaplan MJ, Lu Q, Wu A, Attwood J, Richardson B.
Demethylation of promoter regulatory elements con- tributes to perforin overexpression in CD4+ lupus T cells. J Immunol 2004;172:3652-61.
15) Masi AT, Kaslow RA. Sex effects in systemic lupus erythematosus: a clue to pathogenesis. Arthritis Rheum 1978;21:480-4.
16) Sekigawa I, Naito T, Hira K, Mitsuishi K, Ogasawara H, Hashimoto H et al. Possible mechanisms of gender bias in systemic lupus erythematosus: a new hypo- thesis involving a comparison of systemic lupus ery- thematosus with atopy. Lupus 2004;13:217-22.
17) Cutolo M, Sulli A, Seriolo B, Accardo S, Masi AT.
Estrogens, the immune response and autoimmunity.
Clin Exp Rheumatol 1995;13:217-26.
18) Cutolo M, Sulli A, Villaggio B, Seriolo B, Accardo S. Relations between steroid hormones and cytokines in rheumatoid arthritis and systemic lupus erythema- tosus. Ann Rheum Dis 1998;57:573-7.
19) Verthelyi D. Sex hormones as immunomodulators in health and disease. Int Immunopharmacol 2001;1:
983-93.
20) Okada M, Ogasawara H, Kaneko H, Hishikawa T, Se- kiawa I, Hashimoto H et al. Role of DNA methylation in the transcription of human endogenous retroviruses in the pathogenesis of systemic lupus erythematosus.
J Rheumatol 2002;29:1678-82.
21) Ogasawara H, Okada M, Kaneko H, Hishikawa T, Sekigawa I, Hashimoto H. Possible role of DNA hypomethylation in the induction of SLE: relationship to the transcription of human endogenous retroviruses.
Clin Exp Rheumatol 2003;21:733-8.
22) Groudine M, Eisenman R, Weintraub H. Chromatin structure of endogenous retroviral genes and activa- tion by an inhibitor of DNA methylation. Nature 1981;292:311-7.
23) Sekigawa I, Kawasaki M, Ogasawara H, Kaneda K, Kaneko H, Takasaki Y, et al. DNA methylation: its contribution to systemic lupus erythematosus. Clin Exp Med 2006;6:99-106.
24) Ballestar E. Esteller M, Richardson BC. The epigenetic face of systemic lupus erythematosus J Immunol 2006;176:7143-47.
25) Grimaldi CM. Sex and systemic lupus erythematosus:
the role of the sex hormones estrogen and prolactin on the regulation of autoreactive B cells. Curr Opin Rheumatol 2006;18:456-61.