Role of Sonic Hedgehog in Development of the CNS
PART I
Sonic hedgehog (Shh) functions as a morphogen to induce early dorso-ventral patterning of the central nervous system (CNS) and specifies cell fates of spinal motor neurons. Shh has also been known to promote cell cycle progression of embryonic stem cells as well as adult neural stem/progenitor cells. However, it has not been elucidated how Shh induces neural induction during early development of nervous system.
To elucidate the role of Shh in neural induction, we used a pluripotent embryonal carcinoma cell line, P19 and investigated the effects of Shh on inducing the expression of nestin, the marker for neural stem/progenitor cells. In early stages, we found that Shh promoted the CNS-specific enhancer of the nestin gene through regulation of E-box on nestin 2nd intron 5’-region by Nmyc that is a target gene of Shh signaling. Shh also increased population and proliferation of neural stem cells in vitro or in vivo. In late stages, Shh induced specification of motor neurons as well as dopaminergic neurons. Simultaneously, Shh inhibited the expression of meso-endoderm lineage markers, Gsc, Sox17, and GATA4. The results suggest that Shh promotes neurogenesis in multiple ways: by inducing the neuroectodermal cells, by repressing non-neural cell fates, and by promoting differentiation of neurally committed progenitor cells into motor and dopaminergic neurons.
PART II
Nestin is known as a specific marker for neural stem cells and regulated by the enhancer located on the nestin 2nd intron of nestin gene. Nestin 2nd intron is highly conserved among
species and crucial for nestin expression in central nervous system (CNS) progenitor cells. The transgenic mice that constructed to express green fluorescence protein (GFP) with nestin enhancer have been developed and have great advantage to represent live neural stem cells during research in vivo and vitro.
In this study, we characterized GFP expressing the neural stem cells or progenitors in vitro culture. We also investigated that Ngn1, which is a basic helix-loop-helix (bHLH) transcription factor that is expressed in neuronal precursors during development of the nervous system, can promote nestin expression by nestin 2nd intron enhancer. E-box 2 in 637bp of nestin 2nd intron were thought to be strong candidates for bHLH factors during the early neurogenesis. Ngn1 increased the E-box 2 mediated reporter gene activity. To evaluate the functional mechanism of E-box mediated nestin gene expression in vivo, we generated transgenic (Tg) mice with the 637bp fragment of nestin 2nd intron containing the wild or mutated E-box2. Wild type tg mice successfully exhibited CNS-specific LacZ expression. We also found that LacZ positive cells are proliferating neural progenitors or intermediate progenitors, which is expressed with Ki67 and Tbr2, and can differentiate into vGLUT2 positive glutamatergic neurons in developing cortex. These results indicate that pro-neural Ngn1 promotes nestin expression through nesitn 2nd intron E-box 2 and Ngns mediated LacZ positive cells can differentiate into glutamatergic neurons.
TABLE OF CONTENTS
ABSTACT --- i
TABLE OF CONTENTS --- iii
LIST OF FIGURES --- vi
LIST OF TABLE --- viii
I. INTRODUCTION --- 1
PART I --- 1
A. Hh(hedgehog) --- 1
B. The role of Hh signaling in neural --- 2
C. Dorso-ventral patterning --- 4
D. Cell type specification --- 5
E. Cell proliferation in tumors --- 6
F. P19 embryonic carcinoma cells (EC) --- 7
G. Purpose --- 8
PART II --- 9
A. Neural stem cells --- 9
B. Nestin --- 9
C. basic helix-loop-helix (bHLH) transcription factor --- 10
II. MATERIALS AND METHODS --- 12
A. Materials --- 12
1. Plasmids --- 13
2. P19 cell culture --- 13
3. Differentiation of P19 cells --- 14
3.1 Conventional differentiation --- 14
3.2 Current differentiation --- 14
4. Reverse Transcription Polymerase Chain Reaction (RT-PCR) --- 15
5. Western blotting --- 16
6. Luciferase assay --- 17
7. RNA interference --- 17
8. Immunocytochemistry --- 18
9. In utero injection and electroporation --- 18
10. Immunohistochemistry --- 19
11. Chromatin Immunoprecipitation (ChIP) assay --- 20
III. RESULTS --- 22
PART I --- 22
1. Shh induces neural lineage and blocks meso-endodermal line in P19 cells
--- 22
2. Shh regulates CNS specific Nestin expression through Nestin 2
ndintron in
P19 cells --- 24
3. Shh directly promtes CNS specific Nestin expression in P19 cells --- 27
4. Shh promotes CNS specific Nestin expression through N-myc --- 29
6. Shh directly induces neural differentiation of P19 cells --- 38
7. Shh promotes differentiation of P19 cells into motor and dopaminergic
neurons --- 40
8. Shh regulates the expression of Id and Hes1 during neural differentiation of
P19 cells --- 42
9. Id expression correlates with stemness of NSCs --- 44
10. Id proteins increase the proliferating neural precursors in the developing
brain --- 46
PART. II --- 51
11. Ngn1 regulates CNS specific nestin expression though the NE2 of Nestin
2
ndintron3’-region --- 51
12. Ngn1 promotes neural progenitor cells into glutamatergic neuron in vivo - 57
IV. DISCUSSION and CONCLUSIONS --- 66
REFERENCES --- 72
LIST OF FIGURES
Fig. 1. Shh induces neuroectoderm and blocks meso-endoderm markers in P19
cells --- 23
Fig. 2.
Shh promotes Nestin expression through Nestin 2
ndintron --- 25
Fig. 3. Shh promotes GFP expression regulated by Nestin 2
ndintron enhancer --- 26
Fig. 4. Shh induces Nestin expression through nestin 2
ndintron enhancer region
in P19 cells --- 28
Fig. 5. The sequence alignment of nestin 2
ndintron 5’-region in rat, mouse, and
human --- 31
Fig. 6. N-myc directly regulates CNS specific Nestin expression through Nestin
2
ndintron 5’-region --- 32
Fig. 7. Nmyc promotes Nestin expression in the response of Shh --- 33
Fig. 8.The ectopic expression of Shh in developing dorsal cortex --- 35
Fig. 9. The effect of Shh on prolifeaion of neural stem cells in dorsal cortex of
E15.5 --- 36
Fig. 10. The ectopic effect of Shh on neural stem cells in ventral cortex of E15.5 -- 37
Fig. 11. Shh induces neuronal differentiation of P19 cells --- 39
Fig. 12. Shh induces differentiation of motor and dopaminergic neuron --- 41
Fig. 13. Shh induces the expression of Id and Hes1 in P19 cells --- 43
Fig. 14. Expression of inhibitor of differentiation (Id) correlates with proliferation
Fig. 15. Id proteins increase population of neural stem cells in ventricular zone of
dorsal cortex --- 48
Fig. 16. Id proteins increase proliferation of neural stem cells in the embryonic
brain --- 50
Fig. 17. Ngn1 increased Nestin expression though nestin 2
ndintron in P19 cells ---- 53
Fig. 18. Nestin 2
ndintron conserved sequences in the 3’-region of nestin 2
ndintron
--- 54
Fig. 19. Ngn1 promoted E-box-mediated reporter gene activity in P19 cells --- 55
Fig. 20. Specific induction of Nestin 2
ndintron by pro-neural gene --- 56
Fig. 21. The 3’-region E-box of nestin 2
ndintron mediated expressing LacZ
transgenic mice --- 60
Fig. 22. NesGFP and NesLacZ double Transgenic mice --- 61
Fig. 23. NesGFP/LacZ double Transgenic mice --- 62
Fig. 24. In vivo reporter assay of nestin 2
ndintron 3’-region --- 63
Fig. 25. Characterization of b-gal expressing cells --- 64
Fig. 26. Transition of b-gal expressing cells in dorsal area into vGLUT2 positive
glutamatergic neurons --- 65
LIST OF TABLES
I. INTRODUCTION
PART I
A. Hh (hedgehog)
In vertebrates, three members of the Hedgehog (hh) gene family have been identified : Desert hedgehog (Dhh), Indian hedgehog (Ihh), and Sonic hedgehog (Shh). The most-studied member is Shh, which is initially expressed in the notochord and then in the floor plate (Echelard, Epstein et al. 1993; Chiang, Litingtung et al. 1996; Ingham and McMahon 2001). The signaling properties of the notochord and floor plate are associated with several developmental processes, among them the induction or self-induction of the floor plate, the specification of neuronal identity, and the induction of sclerotome development in ventral somite.
Hh is a potent morphogen involved in the development of limbs and muscles as well as the anterior/posterior organization of the neural tube in vertebrates (Goodrich and Scott 1998). Hh undergo and unusual auto-proteolytic processing event producing an active amino peptide which is covalently modified by cholesterol and/or palmitic acid (Porter, von Kessler et al. 1995; Porter, Young et al. 1996; Pepinsky, Zeng et al. 1998). The amino peptide binds to a heterodimeric receptor complex encoded by the patched (ptc) and smoothened (smo) genes (Marigo, Davey et al. 1996; Stone, Hynes et al. 1996). In the absence of Hh peptide, the 12-membrane pass protein Ptc suppresses the signaling activity of Smo, a seven-membrane pass protein that resembles a G-protein linked-receptor (Alcedo, Ayzenzon et al. 1996). Upon Hh signaling, Ptc-mediated suppression of Smo is relieved, allowing downstream signaling.
The response to Hh signals includes certain conserved components, such as the up-regulation of expression of the ptc1 gene (Goodrich, Johnson et al. 1996) and members of the cubitus interuptus/gli gene family (Alexandre, Jacinto et al. 1996), which encode zinc finger-containing transcription factors. Increased Ptc expression is thought to play a role in further sequestering the Hh peptide, whereas Gli proteins mediate the expression of downstream effectors of Hh signaling. There are three gli genes in the mouse that appear to be functionally redundant (Hui, Slusarski et al. 1994; Mo, Freer et al. 1997). Recent studies suggest a mechanism whereby Hh signaling promotes expression of the downstream effecter gene gli1 and explain how protein kinase A (PKA) can inhibit Hh (Li, Ohlmeyer et al. 1995; Hammerschmidt, Bitgood et al. 1996; Aza-Blanc and Kornberg 1999). The polypeptides encoded by the gli2 and gli3 genes can be proteolytically processed, producing an N-terminal transcriptional repressor, whereas the full length protein acts as an activator. The processing event is promoted by PKA-mediated phosphorylation at multiple sites in the Gli1 and Gli3 polypeptides (Dai, Akimaru et al. 1999; Sasaki, Nishizaki et al. 1999). Hh signaling inhibits this processing and thereby promotes the expression of gli1. The Gli1 protein itself lacks the repressor domain and appears to act exclusively as a positive transcriptional mediator of Hh action (Dai, Akimaru et al. 1999).
B. The role of Hh signaling in neural development
Hh signaling plays a critical role in brain morphogenesis. In early developmental period, Shh is required for ventral specification of central nervous system. In the neural tube, a gradient of Shh is formed and determines ventral specification by influencing the fate of
neural precursors (Ruiz i Altaba, Palma et al. 2002; Ruiz i Altaba, Nguyen et al. 2003; Di Marcotullio, Ferretti et al. 2006). Similarly, Shh determines ventral forebrain and midbrain by specifying dopaminergic and serotoninergic neurons (Di Marcotullio, Ferretti et al. 2006). Holoprosencephaly, in which a loss of Shh signaling occurs, is a disease due to defective ventral patterning and failure of separation of the bilateral lobes of forebrain (Di Marcotullio, Ferretti et al. 2006). In a later embryonic and postnatal period, dorsal brain neocortex, tectum, hippocampus) and hindbrain (cerebellum) are significantly influenced by Shh signaling by regulating the proliferation of neural precursor/stem cells (Di Marcotullio, Ferretti et al. 2006). A number of loss-of-function genetic studies, underscores the role of Shh pathway in enhancing the proliferation of neural progenitor cells, showing that Gli1 function is redundant while Gli2 and Gli3 display specific defects but also overlapping functions (Di Marcotullio, Ferretti et al. 2006).
Shh signaling has an essential function in the maintenance and self-renewal of neural progenitors in stem-cell niches located in several regions of the embryonic, post-natal, and adult brain (Palma and Ruiz i Altaba 2004). In the developing embryonic cortex, Shh signaling increases the growth of cells with stem cell properties and maintains stem cell niches in which these cells exist and proliferate, together with EGF (Palma and Ruiz i Altaba 2004). Similar observations have been reported in the post-natal forebrain sub-ventricular zone (Palma, Lim et al. 2005). In the mammalian developing cerebellum, Shh pathway is also very active. Granule cell progenitors (GCPs), migrating from the rhombic lip over the outer layer of the cerebellar surface (external germinal layer (EGL) (Di Marcotullio, Ferretti et al. 2006), proliferate and are maintained in an undifferentiated state in the outer EGL, in
response to Purkinje cell-derived Shh. Shh signaling is interrupted at the transition from the outer to the inner EGL where GCPs start a differentiation program by exiting the cell cycle and further migrate inwards into the internal granule layer (IGL) where differentiated granule cells (Di Marcotullio, Ferretti et al. 2006).
C. Dorso-ventral patterning
The dorsal-ventral patterning of the telencephalon is a crucial for the normal brain function, because it determines the proportion of two different types of basic neurons, glutamatergic excitatory neurons and GABAergic inhibitory neurons (Yun, Potter et al. 2001). Shh is required for the ventral cell specification, whereas zinc finger transcription factor Gli3 seems to be important for the dorsal cell type specification (Theil, Alvarez-Bolado et al. 1999; Yu, Wang et al. 2009). Recent studies suggest that Gli3 and Shh control the normal proportion of dorsal and ventral cell types to generate appropriate tissue size and shape during brain development (Komada, Saitsu et al. 2008; Stecca and Ruiz i Altaba 2009; Takanaga, Tsuchida-Straeten et al. 2009).
In spinal cord, a concentration gradient of Shh is thought to pattern the ventral neural tube, and these ventral cell types are absent in Shh-/- mice . Based on in vitro and genetic studies, the zinc finger-containing transcription factors Gli1, Gli2, and Gli3 are mediators of the Shh intracellular response (Ruiz i Altaba 1998). The floor plate and adjacent cell types are absent in gli1-/-;gli2-/- mice, but part of the Shh-/- phenotype in the neural tube is alleviated in the Shh-/-;gli3-/- double mutant (Litingtung and Chiang 2000; Park, Bai et al. 2000; Bai and Joyner 2001). This is consistent with the predicted role of Gli3 as a repressor
of the Shh response. Gli3 repressor activity is blocked by Shh (Meyer and Roelink 2003). Misexpression of Gli3 in the chick neural tube caused a ventral expansion of class-I, dorsal progenitor proteins and a loss of class-II, ventral progenitor proteins consistent with expected activity as a repressor of the Shh response (Persson, Stamataki et al. 2002; Meyer and Roelink 2003; Oh, Huang et al. 2009). Activation of the BMP response is sufficient to maintain Gli3 expression in neural plate explants (Meyer and Roelink 2003).
D. Cell type specification
During vertebrate development, the specification of distinct cell types is thought to be controlled by inductive signals acting at different concentration thresholds. It has been demonstrated that the induction of spinal motor neurons (Roelink, Augsburger et al. 1994; Tanabe, Roelink et al. 1995; Litingtung and Chiang 2000; Lewis and Eisen 2001; Motoyama, Milenkovic et al. 2003), midbrain dopaminergic neurons (Hynes, Porter et al. 1995; Wang, Jin et al. 1995; Ye, Shimamura et al. 1998; Kim, Lee et al. 2003; Hammond, Blaess et al. 2009), and basal forebrain cholinergic neurons (Ericson, Muhr et al. 1995; Reilly, Karavanova et al. 2002) are dependent on exposure to Shh that is crucial for patterning and phenotype specification in vivo. In forebrain, expression of the early oligodendrocyte markers Olig2 and PDGFRa corresponds to regions of Shh expression (Nery, Wichterle et al. 2001). Oligodendrocyte progenitors (OLP) generate under the influence of Shh in the ventral forebrain (Lu, Yuk et al. 2000; Tekki-Kessaris, Woodruff et al. 2001).
E. Cell proliferation in tumors
The role of Shh signaling in controlling stem cell behavior raises the question of the relevance of its deregulation for brain tumorigenesis. Indeed, brain tumors (including medulloblastoma) have been described to contain cancer stem cells which retain the property of self-renewal, representing both the potential cell of origin of the tumor and a never-ending reservoir for the maintenance of the tumor mass responsible for disease recurrence. Therefore, the loss of the physiological control of Shh signaling might be determinant for the emergence of cancer stem cells. Accordingly, maintenance of an aberrantly active Shh signaling is responsible for the occurrence of medulloblastoma, a highly invasive malignancy arising from cerebellar progenitor neuronal cells, which accounts for 15–20% of all childhood brain tumors (Ferretti, De Smaele et al. 2005; Stecca and Ruiz i Altaba 2005). Most of the genetic and epigenetic alterations displayed by medulloblastoma are responsible for the disruption of the developmental program of cerebellar neural progenitor cells (Ruiz i Altaba, Palma et al. 2002; Di Marcotullio, Ferretti et al. 2006). In this regard, Shh signaling is a major oncogenic pathway (Ruiz i Altaba, Palma et al. 2002; Ferretti, De Smaele et al. 2005; Di Marcotullio, Ferretti et al. 2006). Interestingly, several medulloblastomas display known gain-of-function or loss-of-function mutations of genes encoding components of the Shh pathway in only a fraction of cases compared to the higher frequency of tumors displaying inappropriately high Shh signaling (Ferretti, De Smaele et al. 2005; Di Marcotullio, Ferretti et al. 2006). These findings suggest the presence of additional genetic or epigenetic hits in pathways which control Shh signaling. To this regard a subset of tumors which carrying deletion of chromosome 17p, also displayed high Shh signaling signature in
the absence of known mutations of components of the Shh pathway (Ferretti, De Smaele et al. 2005; Di Marcotullio, Ferretti et al. 2006).
F. P19 embryonic carcinoma cells (EC)
Mouse embryonic carcinoma (EC) P19 cell is a teratocacinoma cell line derived from transplanted epiblast stage cells of mouse embryo (McBurney, Jones-Villeneuve et al. 1982). P19 cells are pluripotent cells with the ability to differentiate into derivatives of three germ layers in response to different chemical inducers (McBurney, Jones-Villeneuve et al. 1982). These cells are considered to be equivalent to epiblast cells of the early post-implantation blastocyst according to their developmental stages (Riego, Perez et al. 1995; Minucci, Botquin et al. 1996) and to their cell-surface antigen expression and protein synthesis patterns (Evans and Kaufman 1981). It has been widely used as a representative cell system to study events that occur during gastrulation stages of embryonic development (van der Heyden and Defize 2003). Modern developmental biology has benefited significantly from studies on P19 cells to understand the mechanisms of neural and skeletal muscle differentiation (McBurney, Jones-Villeneuve et al. 1982; Bain, Ray et al. 1994; Skerjanc 1999). P19 cells can be induced to differentiate into neurons and glial cells when aggregated in the presence of retinoid acid (RA) (Jones-Villeneuve, McBurney et al. 1982). This method is commonly used for molecular analysis of neural induction and differentiation (Vojtek, Taylor et al. 2003; Akiyama, Yonezawa et al. 2004).
G. Purpose
In this study, I investigated the possible role of Shh during transition of P19 embryonic carcinoma cells into neural cells. To define the roles of Shh signaling in vitro or in vivo, I used a gain of function approach, using Shh-overexpressing P19 cells and in utero electroporation. I provided evidence that Shh promotes not only neural induction of P19 cells through CNS specific Nestin expression but also differentiation of Shh-induced neural stem cells into motor neuron and DA neuron.
PART II
A. Neural Stem cells
Neural stem cells (NSC) in the developing and adult mammalian nervous system proliferate, self-renew and migrate to become functional neurons, astrocytes and oligodendrocytes in right position by various signaling from environment. NSCs can also be regulated form more primitive embryonic stem cells. One of the most well defined characteristic of NSC is Nestin expression, intermediated filament protein (Lendahl, Zimmerman et al. 1990; Dahlstrand, Lardelli et al. 1995). During differentiation of NSC, Nestin protein is replaced to other intermediated protein that expressed in specific cell lineage such as glial ribrilary acidic protein (GFAP) and neurofilaments (NF) (Zimmerman, Parr et al. 1994; Dahlstrand, Lardelli et al. 1995; Lothian and Lendahl 1997).
B. Nestin
Nesin is one of class VI intermediate filament that is expressed in muscle precursors and neural precursors during development. Its gene expression is regulated by the enhancers located on the introns. nestin gene is composed of four exons and three introns. The first intron is important for expression of Nestin in mesoderm originated cells, and the second intron contains the enhancer sequence for expressing in neural precursor cells (NPC) (Zimmerman, Parr et al. 1994). The Nestin 2nd intron is evolutionary conserved; especially the 637bp sequence of 1.8kbp rat Nestin 2nd intron (from 1162bp to 1798bp) shows high homology among rat, mouse and human. This 637bp of Nestin 2nd intron is known that suitable to regulation Nestin expression in central nerves system (Lothian and Lendahl 1997).
To visualize the Nestin expression in neural stem cells, many groups constructed transgenic mice that contain Nestin promoter, the enhancer and reporter gene inside mouse genome (Yamaguchi, Saito et al. 2000; Kawaguchi, Miyata et al. 2001).
C. basic Helix-Loop-Helix (bHLH) transcription factor
During development of central nerves system, the expression of crucial proteins is regulated by complex signaling pathways. One of the regulate molecule called basic Helix-Loop-Helix (bHLH) group is a transcription factor that regulate gene expression. It binds to DNA domain called E-box (CANNTG) as hetero- or homodimer, and stimulates or inhibits the gene transcription (Gradwohl, Fode et al. 1996; Massari, Rivera et al. 1998). Especially, in central nerves system the bHLH transcription factors are known that have crucial roles in differentiation of glia and neuron from neural progenitor cells (Sun, Nadal-Vicens et al. 2001; Zhou and Anderson 2002).
One of the bHLH transcription factors, Neurogenin1 (Ngn1) is known that regulate neuronal cell fate. Expression of Ngn1 suppresses JAK-STAT pathway, which is important for glial differentiation and induces neurogenesis by functioning as a transcriptional activator. JAK is one of receptor-associated tyrosine kinase, which activated by leukemia inhibitory factor (LIF) or cilicary neurotrophic factor (CNTF). When LIF or CNTF binds to receptors, signal transducers and activators of transcription 1 and 3 (STAT1 and 3) is phosphorylated by JAK, and phosphorylated STAT makes complex with Smad and CBP/p300 to activate transcription of glial genes (i.e GFAP) to induce glial differentiation. However, Ngn1 blocks phosphorylation of STAT and STAT/CBP/p300/smad complex conformation to inhibit glial
gene expression, and stimulates neuronal differentiation genes (i.e NeuroD) expression with CBP/p300/smad complex (Sun, Nadal-Vicens et al. 2001).
In mammals, Ngns and Mash1 are expressed before neuronal differentiation in precursors and perform a determination function similar to that of the Drosophila pro-neural genes in neural development. Ngn1 and Ngn2 promote neurogenesis (Nieto, Schuurmans et al. 2001; Sun, Nadal-Vicens et al. 2001) and the dorsal character of telencephalic neurons through repression of the ventral pro-neural gene Mash1 (Fode, Ma et al. 2000). Expression of Ngn1 and Ngn2 are overlapped in the CNS although they control two distinctive waves of neurogenesis in developing dorsal root ganglia (Ma, Fode et al. 1999). Ngn1 and Ngn2 knock out mice exhibit defects in cranial ganglia and fail to activate downstream genes including Math2, NeuroD, and a Notch ligand Delta 1 (Ma, Chen et al. 1998). In Mash1 mutant embryos, Ngn1 and NeuroD fail to be expressed in some developing olfactory neurons (Cau, Gradwohl et al. 1997). While loss-of-function analysis in knock out mice have revealed the bHLH cascades, molecular details of these bHLH proteins have been determined by gain-of-gunction studies. In telencephalon, Ngn1 and Ngn2 expression begins at E8, peaks at E11-14, and disappears by E16 in mouse brain (Ma, Chen et al. 1998). Ngns and Mash1 bind to E-box promoter elements (CANNTG) like other bHLH transcription factors and have been shown to induce transcription driven by NeruoD1 and NeuroD2 promoter fragments (Shibata, Oda et al. 1999; Farah, Olson et al. 2000; Huang, Hsu et al. 2000).
In this work, I elucidated the novel function of pro-neural bHLHs, Ngn1, Ngn2 and Mash1, regulating the Nestin expression via E-box 2 of Nestin 2nd intron 3’-region.
II. MATERIRALS AND METHODS
A. MATERIRALS
βIII-tubulin (Tuj-1) specific antibody from Berkeley antibody company (Covance, CA, U.S.A); neurofilament-160kDa (NF-M) specific antibody from Zymed Laboratory (South San Francisco, CA, U.S.A); 2', 3'-cyclic nucleotide 3'-phosphodiesterase (CNPase) specific antibody, Choline acetyltransferase (ChAT), Neurogenin1, and Hb9 specific antibody from Chemicon (Temecula, CA, U.S.A); Mouse IgG , glial fibrillary acidic protein (GFAP) specific antibody, neuronal cell adhesion molecule (NCAM) specific antibody, All-trans-retinoic acid(RA) , Tyrosine Hydroxylase (TH) specific antibody, 2-mercaptoethanol, o-phenylenediamine dihydrochloride (OPD), Forskolin, Tris, Glycine, sodium dodecyl sulfate(SDS), polyacrylamide, bis-acrylamide, and TEMED from Sigma (St. Louis, MO, U.S.A); DMEM and N2-supplement from GibcoBRL (Grand Island NY, U.S.A); FBS from Hyclone Inc. (Logan, UT, U.S.A); 5E1 hybrydoma cell from DSHB (IA, U.S.A); Enhanced chemiluminescence (ECL) kit from Pierce (Rockford, IL, U.S.A); Westran PVDF membranes from Schleicher & Schuell (Dassel, Germany); RNAzolTM B regents from TEL-TEST Inc. (Frendwood, TX, U.S.A); Frist strand cDNA synthesis kit from Roche (Indianapolis, U.S.A); Super Taq DNA polymerase from Korea B&G (Suwon, Korea); Alexa Fluor 488 anti-mouse, rabbit, and chicken, Alexa Fluor 568 anti-mouse, rabbit, and chicken from Molecular Probes (Eugene, OR, U.S.A); Secondary antibody conjugated horse radish peroxidase (HRP) from Zymed (San Francisco, CA, U.S.A); antibody conjugated biotin, avtivin-biotin ABC Kit, and Vectashield mounting solution from Vector laboratories
(Burilingame, CA, U.S.A); pGL3-promoter vector, pRL-TK and luciferase assay system from Promega (Madison, WI, U.S.A).
All other chemicals were obtained from Sigma-Aldrich (St. Louis, MO, U.S.A).
B. METHODS 1. Plasmids
Korean male genomic DNA was used as a template for polymerase chain reaction (PCR) to obtain exons 1 and 2 of Shh. Exon 2 was elongated by the second PCR using a primer containing 30 bp of the 50 of exon 3. The two fragments (304 bp of exon 1 and 294 bp of exon 2 + 3) were inserted in pCDNA3.1 to obtain pShhN, which encoded the N-terminal peptide [1–197 amino acids] of Shh. LacZ and Shh DNA were inserted in front of internal ribosome entry site (IRES) and enhanced GFP sequence. pcDNA3.1/His-Ngn1 was previously described (Kim et al., 2004). Nestin 2nd intron 5’-region (1.0kb) were synthesized with BglII and BamHI compatible cohesive ends and were inserted into the BglII site of pGL3-promoter vector (Promega) to generate pGL3-NE1.0luc. the pNPEeGFP construct was previously described from Dr. Masahiro Yamaguchi (Yamaguchi, Saito et al. 2000). pCMV-N-myc construct was previously described from KH Suk (Jung, Lee et al. 2005).
2. P19 cell culture
P19 cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM) with 10% fetal bovine serum (FBS), 100U/㎖ penicillin, 100㎍/㎖ streptomycin in a standard humidified atmosphere at 37℃. After the cultures reached 80-90% confluence, the cells were
harvested with 0.25% trypsin and 1mM EDTA for 2-3min at 37℃, re-plated after 1:10 dilution on 100mm culture dish.
3. Differentiation of P19 cells
3.1 Conventional neuronal differentiation
P19 cells were allowed to aggregate in bacterial Petri dishes at a seeding density of 1 x 105 cells/㎖ in the presence or absence of 0.5 µM all-trans-retinoic acid in DMEM with 10% FBS. After 4 days of aggregation, cells were dissociated into single cells by 0.05 % trypsin-EDTA, and were replaced in tissue culture dish at a density of 1 x 105 cells/㎖ in DMEM:F12 supplemented with N2 supplement [5㎍/㎖ insulin, 50㎍/㎖ human transferring, 20mM progesterone, 60µM putresine, and 30nM sodium selenite]. The cells were then allowed to adhere and cultured for 4 days with replacement of the medium every 2 days.
3.2 Current neuronal differentiation
P19 cells were plated in tissue culture dish at a 2 x 105 cells/㎖ of density in DMEM with 10% FBS. To induce neuronal differentiation, the media were removed, and the cells were washed with HBSS and were replaced with DMEM:F12 with N2 supplement for 4 days.
4. Reverse Transcriptase Polymerase Chain Reaction (RT-PCR)
Total RNA was isolated using a RNAzolTM B and cDNA was synthesized using First-strand cDNA synthesis kit and 1㎍ of total RNA following the manufacture’s instructions.
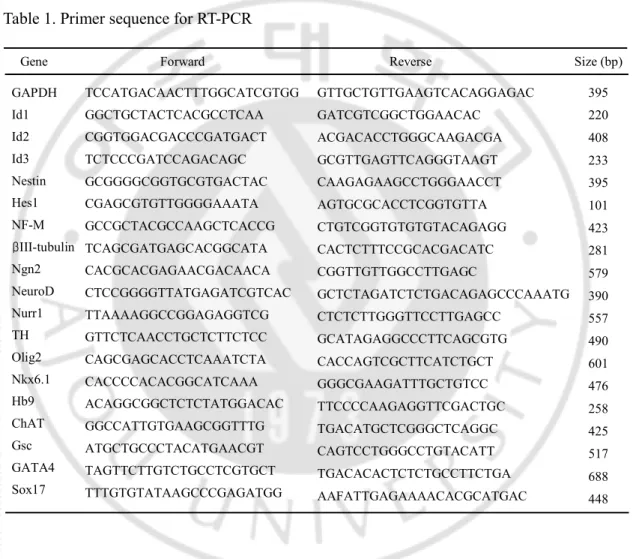
The PCR reactions were carried out the following condition; denaturation at 94℃ for 30s, annealing at indicated temperature (Table 1) for 30s, and extension at 72℃ for 1 min. Primer sequences used and product sizes were as follows (Table 1). Primer sequences (forward and reverse) and product sizes (base pairs) were as follows (Table 1). The PCR products were analyzed on a 1.5% agarose gel.
Table 1. Primer sequence for RT-PCR
Forward Reverse Size (bp)
TCCATGACAACTTTGGCATCGTGG
GAPDH GTTGCTGTTGAAGTCACAGGAGAC 395
Nestin GCGGGGCGGTGCGTGACTAC CAAGAGAAGCCTGGGAACCT
408
NF-M GCCGCTACGCCAAGCTCACCG CTGTCGGTGTGTGTACAGAGG 423
TH GTTCTCAACCTGCTCTTCTCC GCATAGAGGCCCTTCAGCGTG 490 Nurr1 TTAAAAGGCCGGAGAGGTCG CTCTCTTGGGTTCCTTGAGCC 557 Olig2 CAGCGAGCACCTCAAATCTA CACCAGTCGCTTCATCTGCT 601 Nkx6.1 CACCCCACACGGCATCAAA GGGCGAAGATTTGCTGTCC 476 Ngn2 CACGCACGAGAACGACAACA CGGTTGTTGGCCTTGAGC 579 NeuroD CTCCGGGGTTATGAGATCGTCAC GCTCTAGATCTCTGACAGAGCCCAAATG 390
Hb9 ACAGGCGGCTCTCTATGGACAC TTCCCCAAGAGGTTCGACTGC 258 ChAT GGCCATTGTGAAGCGGTTTG TGACATGCTCGGGCTCAGGC
425 Gene
Id2 CGGTGGACGACCCGATGACT ACGACACCTGGGCAAGACGA
220 Id1 GGCTGCTACTCACGCCTCAA GATCGTCGGCTGGAACAC
233 Id3 TCTCCCGATCCAGACAGC GCGTTGAGTTCAGGGTAAGT
Hes1 CGAGCGTGTTGGGGAAATA AGTGCGCACCTCGGTGTTA 101
Gsc ATGCTGCCCTACATGAACGT CAGTCCTGGGCCTGTACATT
517 GATA4 TAGTTCTTGTCTGCCTCGTGCT TGACACACTCTCTGCCTTCTGA
688 Sox17 TTTGTGTATAAGCCCGAGATGG
AAFATTGAGAAAACACGCATGAC 448 395
5. Western blotting
Whole cell lysates were prepared as follows; Cells were washed with cold phosphate buffered saline (PBS) twice and drained. Cells were resuspended in RIPA buffer [150mM Sodium chloride, 1% NP-40, 0.1% SDS, 50mM Tris-HCl (pH8.0), 0.5% sodium deoxycholate] and kept on ice for 5 min. After centrifugation at 12000 rpm for 20 min, the supernatant was collected and protein content was assayed by Bradford method. Each 60 ㎍ proteins were separated on 8% or 12% SDS polyacrylamide gel electrophoresis and transferred to PVDF membranes. Membranes were incubated in blocking solution [5% nonfat dry milk in 10mM Tris-HCl (pH7.4), 100 mM NaCl, and 0.1% Tween 20] for 1 hour at room temperature and with mouse anti- ShhN (R&D system, 1:100), mouse anti-N-myc (Calbiochem, 1:1000), mouse βIII -tubulin (Tuj1) (Covance, 1:1000), mouse anti-neurofilament-160 kD (NF-M) (Zymed, 1:1000), mouse anti-glial fibrillary acidic protein (GFAP) (Sigma, 1:1000), mouse anti-2', 3'-cyclic nucleotide 3'-phosphodiesterase (CNPase) (Milipore, 1:1000), and mouse anti-actin (Chemicon, 1:3000) in TTBS [10 mM Tris-HCl (pH7.4), 100 mM NaCl, and 0.1% Tween 20] solution for overnight at 4°C. The secondary antibody was horseradish-peroxidase (HRP)-conjugated to goat anti-mouse IgG (Zymed, 1:5000), rabbit IgG (Zymed, 1:10000) or goat IgG (1:5000) and the proteins were visualized using an enhanced chemiluminescence (ECL) kit following the manufacture’s recommendation.
6. Luciferase assay
P19 cells were transfected with plasmids containing pGL3-promoter (Promega, Madison, WI), pGL3-NE1.7kb, 1.0kb, 0.7kb, 0.4(383bp)kb, 0.3(319bp)kb, 0.4(390bp)kb , pCMV-RL-TK (Promega, Madison, WI) , pCMV-N-myc, pcDNA3.1-mNgn1 and pcDNA3.1-hShhN expression vector using LipofectAMINE (Invitrogen). After 48 hours, cell extracts were prepared according to the manufacturer’s protocol and analyzed by luciferase assay using the Dual-Luciferase assay system (Promega, Madison, WI).
7. RNA interference
The siRNA were Dharmacon ON-TARGETplus SMART for mouse N-myc and siGENOME Non-Targeting siRNA (Thermo Fisher Scientific). The sequences of siRNA are for N-myc-#1, 5′-UCUAACAACAAGGCGGUAA-3′; N-myc-#2, 5′-GAGGAUACCUUGA GCGACU-3′; N-myc-#3, UCGAAUUGGGCUACGGAGA -3′; and N-myc-#4, 5′-CCGGUGAACAAGCGAGAGA-3′. Cells were grown to 30% in 6 well plates. I was transfected siRNA into P19 cells at final concentration of 20nM using Metafectene reagent (Biontex, Germany) according to the manufacturer's instruction. Cells were harvested 24h after transfection, and N-myc expression was analyzed by reverse transcription-PCR (RT-PCR) as described above.
8. Immunocytochemistry
Differentiated P19 and P19/hShhN cells were fixed with 4% paraformaldehyde solution or 100% methanol for 10 min at 4℃. The fixed was washed with PBS solution, three times and incubated with 10% normal horse serum and goat serum, 0.1% BSA and 0.03% Triton X-100 in PBS (PBS-T) for 1hour. After sufficient washing, the cells were incubated with primary antibodies overnight at 4℃. The following primary antibodies were used at the concentrations given: GFP (1:500), β-III-tubulin (Tuj1) (1:200), NF-M (1:200), TH (1:200), ChAT (1:200) and Hb9 (1:200). Following several washing with 0.03% Triton X-100 containing PBS-T solution, cells were incubated in Alexa 488 anti-mouse IgG (1:500), Alexa 488 anti-rabbit IgG (1:500), Alexa 568 anti-mouse IgG (1:500) and Alexa 568 anti-rabbit IgG (1:500) secondary antibody for 1 hour in dark chamber. After sufficient washing with PBS-T, cells were mounted on cover slips by using Vectorshield (mounting medium for fluorescence, Vector) with Hoechst (Molecular probe) and photographed using a fluorescent microscope.
9. In utero injection and electroporation
Pregnant ICR mice wild type gestational age of E13.5 was anesthetized with ketamine (50㎎/㎏) and rumpun (2.5㎎/㎏). The uteri were taken out after a 3 cm midline laparotomy. DNA (4 ㎍/㎕) in PBS containing 0.05% fast green dye (Sigma) was drawn into a 90mm glass capillary tube (GD-1; Narishige, Tokyo, Japan) prepared a micropipette puller P-97 (Sutter Instrument, Novaato, CA). 1㎕ of DNA solution was injected into the lateral ventricle of the E13.5 embryo brain and electroporation was performed with a tweezertrode
and five pulses of 45V for 50 msec and the 950 msec interval using ECM830 pulse generator (BTX, USA), as described previously (Saito and Nakatsuji 2001).
10. Immunohistochemistry
Embryos were removed from uteri and fixed with 4% PFA at 4℃ for 2 hr and then in 30% sucrose in PBS at 4℃ overnight. The embryos were embedded in tissue freezing medium (Leica, Deerfield, IL, USA), and serially sectioned in a 10 ㎛ thickness using a cryostat (CM3050 S, Leica). The section was dry at RT for overnight and wash with PBS two times. After wash, the sample was incubated with antigen retrieval solution (0.01 M sodium citrate (pH 6.0), 0.05 % Tween-20) for 1hr at RT and heated for 5 min at 95 ℃. Then, the sample was replaced in permiablization buffer (0.1 % Triton X-100 in PBS) and incubated for 10 min three times. Specimens were incubated with primary antibodies against GFP (invitrogen, 1:500), Shh (R&D system, 1:500), Nestin (Rat401) (DSHB, 1:500), Ki67 (Abcam, 1:500), β-gal (Abcam, 1:1000), β-gal (Cappell, 1:4000), Mash1 (BD, 1:25), Olig2 (Milipore, 1:500), Tbr1 (Abcam, 1:500), Tbr2 (Abcam, 1:500), β-III-tubulin (Tuj1) (Covance, 1:1000) and Sox2 (Abcam, 1:500) in blocking solution for overnight at 4℃. Following several washing with PBST, tissues were incubated in Alexa 488 anti-mouse IgG, rabbit IgG, chicken IgG, Alexa 568 anti-mouse IgG, rabbit IgG and chicken IgG (Invitrogen, 1:500) from goat secondary antibody for 1hr in dark chamber. After sufficient washing with PBST, tissues were mounted on cover slips using Vectorshild (Mounting medium for fluorescence, Vector). The fluorescent images were obtained with a conforcal laser scanning microscope (LSM 510, Zeiss, Thornwood, NY, USA).
11. Chromatin Immunoprecipitation (ChIP) assay
11.5 dpc embryos were sacrificed and the forebrains were dissociated with trituration in the presence of 0.25% trypsin-EDTA 5 min on ice. The 100 ㎎ of dissociated tissues were washed with PBS and diluted in 1 ㎖ of PBS and formaldehyde was added to a final concentration of 1 %(v/v), and the dishes were incubated on a shaker at room temperature for 10 min. The single cells were counted and added with 1% SDS lysis buffer as 106 cells per 200㎕ to swell on ice for 10 min. then, the cells were sonicated with 5 sec and three times pulse using 600 watt sonicator at 21 % amplification on ice. 10-fold ChIP dilution buffer was added into the tube with protease inhibitor. Chromatin samples (200 ㎕) were precleared with 75 ㎕ of protein A agarose/salmon sperm DNA (50 % slurry) for 30min at 4℃ with agitation. The beads were collected by brief centrifugation to a new tube, and 5 ㎍ of each antibody were added. Samples were incubated overnight at 4℃ on a rotator. 60 ㎕ of blocked protein A agarose/salmon sperm DNA (50 % slurry) were added, and incubation was continued for an additional 1 hr at 4 ℃ on the rotator. Beads were washed five times with low salt immune complex wash buffer (0.1 % SDS, 1% Triton X-100, 2 mM EDTA, 20mM Tris-HCl, pH 8.1 and 150mM NaCl), high salt immune complex wash buffer (0.1 % SDS, 1% Triton X-100, 2 mM EDTA, 20 mM Tris-HCl, pH 8.1 and 500mM NaCl), LiCl salt immune complex wash buffer (0.25 M LiCl, 1 % deoxycholic acid(sodium salt), 1 mM EDTA, 10 mM Tris-HCl, pH8.1) and twice with TE buffer (10 mM Tris-HCl, 1 mM EDTA, pH 8.0). the bound material was eluted by incubating the beads with 250 ㎕ of elution buffer (0.1 M sodium bicarbonate, 1.0 % (w/v) SDS) with vigorous shaking for 10 min. The elution was repeated two times. The two elutants were pooled, added by 20 ㎕ of 5 M NaCl
solution and heated to 65℃ for 4 hrs to reverse the Schiff base linkage. Then 10 ㎕ of 0.5 M EDTA, 20 ㎕ of 1 M Tris-HCl (pH 6.5) and 2 ㎕ (10mg/ml) of proteinase K were added into the tube and incubated at 45 ℃ for 1 hr. DNA was collected by ethanol precipitation and resuspended in 20 ㎕ of water. A portion of this (0.5 ㎕) was used for PCR amplification with SuperTaq polymerase.
III. RESULTS
PART I
1. Shh induces neural lineage and blocks meso-endodermal lineage in P19 cells.
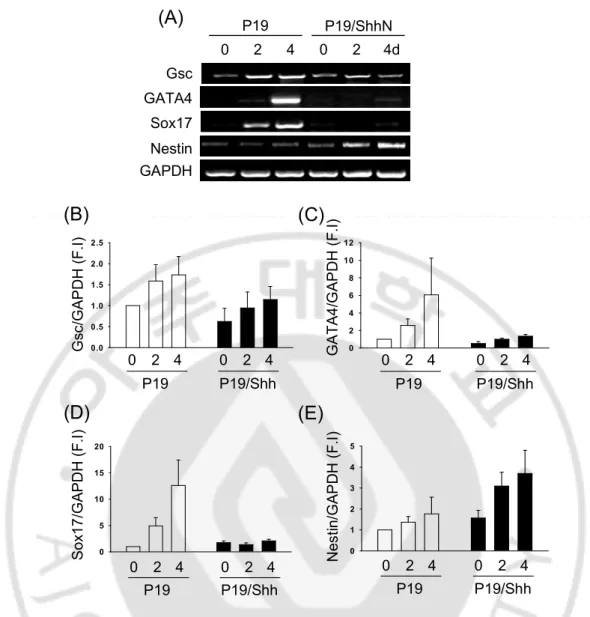
To know the function of Shh during early development, I used P19 embryonic carcinoma (EC) cells and generated Shh-overexpressing P19 cell line. I was cultured aggregation to know only Shh function without DMSO or RA in P19 and P19/Shh cells. I was analyzed the expression of three germ layer markers using RT-PCR. Shh increased dramatically expression of Nestin in P19 cells (Fig. 1A and E). However, the expression of Gsc, a meso-endodermal marker (Blum, Gaunt et al. 1992), and GATA4, a mesoderm marker (Heikinheimo, Scandrett et al. 1994; Gianakopoulos and Skerjanc 2005), were barely detectable in P19/Shh cells, even though these markers induced in P19 cells (Fig. 1A-C). The endodermal marker Sox17 (Hudson, Clements et al. 1997; Alexander and Stainier 1999; Kanai-Azuma, Kanai et al. 2002) was expressed a lower level in P19/Shh cells (Fig. 1A and D). These results suggest that Shh induced the expression of neural stem cell marker but blocked major mesodermal and endodermal markers in P19 cells.
GAPDH GATA4 Nestin Sox17 Gsc 2 4 2 4d P19 0 0 P19/ShhN 0 2 4 6 8 1 0 1 2 0 5 10 15 20 S o x 1 7 /G A P D H ( F .I ) 0 2 4 P19 0 2 4 P19/Shh 0 .0 0 .5 1 .0 1 .5 2 .0 2 .5 G s c /G A P D H ( F .I ) 0 2 4 P19 0 2 4 P19/Shh G A T A 4 /G A P D H ( F .I ) 0 2 4 P19 0 2 4 P19/Shh 0 1 2 3 4 5 N e s ti n /G A P D H ( F .I ) 0 2 4 P19 0 2 4 P19/Shh
(A)
(B)
(C)
(D)
(E)
Fig. 1. Shh induces neuroectodermal and blocks meso-endodermal markers in P19 cells. Cells were cultured to form aggregate in DMEM with 10%FBS for 4days without the treatment of RA (retinoic acid). (A) Reverse transcription polymerase chain reaction analysis of the expression of neuroectoderm (Nestin), meso-endoderm (Gsc, Sox17, and GATA4) markers of P19 cell aggregates and P19/Shh cell aggregates. The percentage of Nestin, Gsc, Sox17, and GATA4 expression in (B~E).
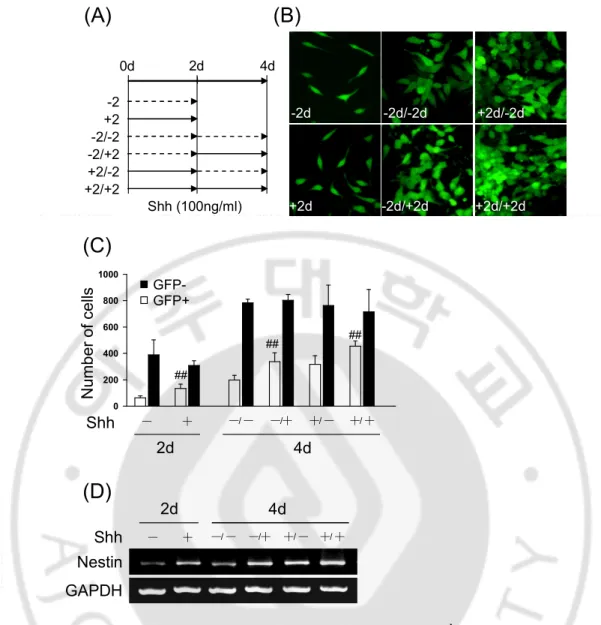
2. Shh regulates CNS specific Nestin expression through Nestin 2nd intron in P19 cells. Previous data showed that Shh induces Nesitn expression in P19 cells for aggregation culture. Because Nestin was expressed tissue specific by intron that first intron promotes differentiation into muscle or mesodermal cells and second intron promotes differentiation into neurons, I found whether the increased Nestin by Shh is CNS specific expression using pNPEeGFP construct that including Nestin promoter, EGFP and Nestin 2nd intron (Fig. 2A). I was cultured P19/pNPEeGFP cells with control or Shh-conditioned medium as well as Shh neutralizing antibody, 5E1, for 4day in differentiation condition. Shh increased the number of GFP+ cells compare with treatment of control-conditioned medium but Shh-induced GFP+ cells decreased by 5E1 (Fig. 2B and C). Shh also increased Nestin expression and decreased its expression by 5E1 (Fig. 2C). To confirm whether Shh is important to express Nestin gene, I was cultured time differently P19/pNPEeGFP cells with or without the treatment of Shh (100 ng/㎖) (Fig. 3A). I showed that GFP positive cells only were increased in the presence of Shh but were not in the absence of Shh (Fig. 3B and C). Nestin expression also increased in the presence of Shh (Fig. 3D). These results suggest that Shh is important factor for the regulation of CNS specific Nestin expression during neural differentiation.
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 N e s ti n /G A P D H (F .I ) ## ## Nestin GAPDH ┼ ─ ┼ ─ 5E1 IgG ┼ ─ ┼ ─ Control Shh 4 day Control ShhN Ig G 5 E 1 0 5 10 15 20 25 30 35 G F P + /T o ta l c e lls ( % ) ┼ ─ ┼ ─ 5E1 IgG ┼ ─ ┼ ─ Control Shh ## # 0 5 10 15 20 25 30 35 G F P + /T o ta l c e lls ( % ) ┼ ─ ┼ ─ 5E1 IgG ┼ ─ ┼ ─ Control Shh ## # 4d (A) (B) (C) (D)
Fig. 2. Shh promotes Nestin expression through Nestin 2nd intron. (A) Construct of pNPEeGFP including Nestin promoter, EGFP and Nestin 2nd intron. (B) GFP positive cells were increased in the presence of Shh-conditioned medium but not in the presence of control-conditioned medium in P19/pNPEeGFP cells. Increased GFP positive cells were decreased by 5E1 as Shh neutralizing antibody, and results are summarized in (C). The data are presented as mean relative GFP positive cells SD±. (D) RT-PCR analysis of Nestin expression in the presence of Shh or control conditioned medium with IgG or 5E1 in P19 cells. (##P<0.01, #P<0.05)
Nestin GAPDH Shh ─ ┼ ─/ ─ ─/┼ ┼/ ─ ┼/ ┼ 2d 4d -2 +2 -2/-2 -2/+2 +2/-2 +2/+2 2d 4d Shh (100ng/ml) -2d +2d -2d/-2d -2d/+2d +2d/-2d +2d/+2d 0d 0 200 400 600 800 1000 N u m b e r o f ce lls Shh ─ ┼ ─/ ─ ─/┼ ┼/ ─ ┼/ ┼ 2d 4d
(A)
(B)
(C)
(D)
## ## ## GFP-GFP+Fig. 3. Shh promotes GFP expression regulated by Nestin 2nd intron enhancer. Cells were cultured in N2 media with the treatment of Shh (100ng/㎖) for 4days. (A) The treatment time of Shh for 4days. (B) GFP positive cells increased in the presence of Shh but not in the absence of Shh. (C) RT-PCR analysis of Nestin expression.
3. Shh directly promotes CNS specific Nestin expression in P19 cells.
However, it is not clear how Shh regulates CNS specific Nestin expression. During neurogenesis, Nestin expression has been know to regulate CNS specific through Nestin 2nd intron enhancer by pro-neural genes; Neurogenin1/2 and Mash1 as well as Sox2 and POU domain proteins; Brn1/2 and Oct4. I hypothesized that Shh signaling may directly induce Nestin expression through regulation Nestin 2nd intron enhancer. I showed that Shh increased dose-dependently Nestin expression in P19/pNPEeGFP cells (Fig. 4B). To know whether Shh regulates CNS specific Nestin expression through Nestin 2nd intron, I used Nestin 2nd intron reporter gene assay in P19 cells with or without Shh and analyzed luciferase activity (Fig. 4A). Shh increased 2-fold of lucifease activity in Nestin 2nd intron full length reporter gene but not in 3’-region (Fig. 4C). Ngn1 as a positive control was increased luciferase activity in both Nestin 2nd intron full length and 3’-region reporter gene (Fig. 4C). These results suggest that Shh may induce CNS specific Nestin expression through Nestin 2nd intron 5’-region enhancer.
0 2 4 6 8 L u c if e ra s e a s s a y ( F .I ) Shh Ngn1
-
-
Shh Ngn1 NE1.7luc NE0.7luc ## 0 2 4 6 8 L u c if e ra s e a s s a y ( F .I ) Shh Ngn1-
-
Shh Ngn1 NE1.7luc NE0.7luc ## 0 2 4 6 G F P e x p re s s io n ( F .I ) 0 1 10 100 (ng/ml) Shh 0 2 4 6 G F P e x p re s s io n ( F .I ) 0 1 10 100 (ng/ml) Shh (A) (B) (C)Fig. 4. Shh induces Nestin expression through Nestin 2nd intron enhancer region in P19 cells. (A) A schematic presentation of rat Nestin 2nd intron full length and reporter genes linking Luc containing different lengths of the Nestin 2nd intron region, pGL3-NE1.7luc and pGL3-NE0.7(+2). (B) P19/pNPEeGFP cells were cultured in the present of Shh (0, 10, 100 ng/㎖). GFP expression was increased by Shh in a dose-depent manner. (C) P19 cells were transfected Nestin 2nd intron reporter gene, pGL3-NE1.0luc and pGL3-NE0.7(+2)luc and Shh (100ng/㎖) using Lipofectamine Reagent. The luciferase activity was increased by Shh in Nestin 2nd intron full length (1.7kb) but not 3’-region (0.7kb). (##P<0.001)
4. Shh promotes CNS specific Nestin expression through N-myc
N-myc is a target gene of Shh signaling and bHLH transcription factor to regulate target gene in various cells (Oliver, Grasfeder et al. 2003; Kenney, Widlund et al. 2004; Knoepfler and Kenney 2006). N-myc also can bind on E-box (CACGTG or CATGTG) of target gene promoter or enhancer to regulate cell proliferation (Ayer and Eisenman 1993; Vervoorts and Luscher 1999). Previous studies reported that N-myc binds on E-box of human Nestin 2nd intron 5’-region and promotes Nestin expression in human neuroblastoma (Thomas, Messam et al. 2004).
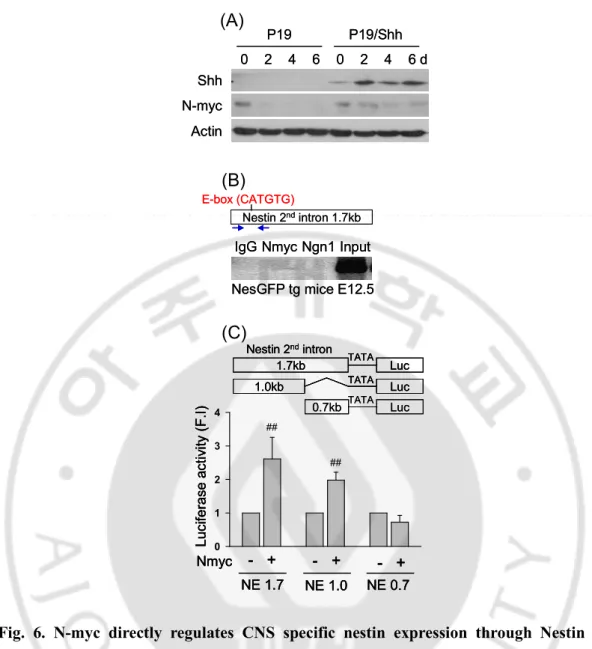
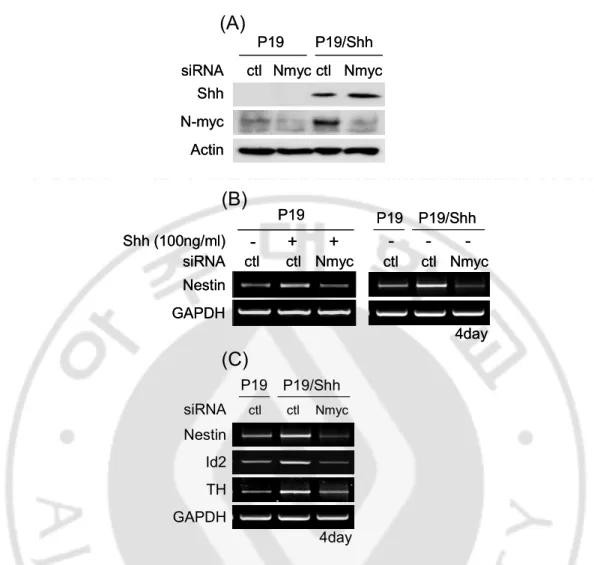
I found that N-myc binding E-box are in Nestin 2nd intron 5’-region of rat, mouse, and human (Fig. 5) and N-myc expressed in P19 cells (Fig. 6A). During differentiation, Shh maintained N-myc expression in P19 cells (Fig. 6A). To know whether N-myc can bind on E-box of Nestin 2nd intron 5’-region, I used ChIP assay to know N-myc binding affinity on E-box of Nestin 2nd intron in vivo. I showed that N-myc binds on Nestin 2nd intron 5’-region in E12.5 of ICR mice but not bHLH transcription factor as Ngn1 which can bind on E-box and promotes Nestin expression through E-box (CAGATG) of Nestin 2nd intron 3’-region (Fig. 6B). To confirm whether N-myc binds on Nestin 2nd intron to regulate Nestin expression, I found luciferase activity about Nestin 2nd intron reporter gene in P19 cells. N-myc was increased the luciferase activity of Nestin 2nd intron full length (1.7kb) and 5’-region 1.0.kb but not Nestin 2nd intron 3’-region (0.7kb) (Fig. 6C). These results suggest that N-myc may promote CNS specific Nestin expression through binding on E-box of Nestin 2nd intron 5’-region.
down-regulation of N-myc using siRNA in P19 cells. I validated siN-myc in P19 and P19/Shh cells that expressed N-myc (Fig. 7A). Nestin expression increased in both the presence of Shh (100 ng/㎖) and P19/Shh cells but decreased by siN-myc compared with siControl using RT-PCR (Fig. 7B). N-myc also regulates the expression of Id2 and TH that know to be regulated by Shh. Id2 and TH expression decreased by siN-myc in P19/Shh cells (Fig. 7C). These results suggest that N-myc directly promotes CNS specific Nestin expression in the presence of Shh.
Fig. 5. The sequence alignment of Nestin 2nd intron 5’-region in rat, mouse, and human E-box (CATGTG) that N-myc can bind is in Nestin 2nd intron 5’-region of rat, mouse, and human. Rat and mouse E-box were conserved but not human.
IgG Nmyc Ngn1 Input
Nestin 2ndintron 1.7kb E-box (CATGTG)
NesGFP tg mice E12.5 IgG Nmyc Ngn1 Input
Nestin 2ndintron 1.7kb E-box (CATGTG)
NesGFP tg mice E12.5
0 2 4 6 P19 P19/Shh Actin Shh N-myc 0 2 4 6 d 0 2 4 6 P19 P19/Shh Actin Shh N-myc 0 2 4 6 d 0 1 2 3 4 L u c if e ra s e a c ti v it y ( F .I ) Nmyc - + NE 1.7 NE 1.0 NE 0.7 - + - + ## ## Luc 1.7kb Nestin 2ndintron Luc 1.0kb Luc 0.7kb TATA TATA TATA 0 1 2 3 4 L u c if e ra s e a c ti v it y ( F .I ) Nmyc - + NE 1.7 NE 1.0 NE 0.7 - + - + ## ## 0 1 2 3 4 L u c if e ra s e a c ti v it y ( F .I ) Nmyc - + NE 1.7 NE 1.0 NE 0.7 - + - + ## ## Luc 1.7kb Nestin 2ndintron Luc 1.0kb Luc 0.7kb TATA TATA TATA Luc 1.7kb Nestin 2ndintron Luc 1.0kb Luc 0.7kb TATA TATA TATA
(A)
(B)
(C)
Fig. 6. N-myc directly regulates CNS specific nestin expression through Nestin 2nd intron 5’-region. (A) Western blot of Shh and N-myc expression during neural differentiation of P19 and P19/Shh cells. (B) ChIP assay of N-myc binding affinity on Nestin 2nd intron 5’-region in E12.5 of ICR mice. (C) Luciferase activity of N-myc on Nestin 2nd intron, full length (1.7kb), 5’-region (1.0kb), and 3’-region (0.7kb). luciferase activity increased in full length and 5’-region by N-myc but not 3’-region.
N-myc Actin siRNA ctl Nmyc P19 P19/Shh ctl Nmyc Shh N-myc Actin siRNA ctl Nmyc P19 P19/Shh ctl Nmyc Shh - + + ctl ctl Nmyc siRNA Shh (100ng/ml) Nestin GAPDH P19 P19/Shh ctl ctl Nmyc P19 4day - - -- + + ctl ctl Nmyc siRNA Shh (100ng/ml) Nestin GAPDH P19 P19/Shh ctl ctl Nmyc P19 4day - - -P19 P19/Shh siRNA ctl ctl Nmyc Nestin Id2 TH GAPDH 4day
(A)
(B)
(C)
Fig. 7. N-myc promotes Nestin expression in the response of Shh. Cells were transfected with siControl or siN-myc in Shh (100 ng/㎖) -treated P19 cells or P19 /Shh cells. (A) The validation of siN-myc in P19 and P19/Shh cells. (B) Nestin expression increased in both treated P19 cells and P19/Shh cells but decreased by siN-myc using RT-PCR. (C) Shh-induced Id2 and TH expression decreased by siN-myc.
5. Shh induces proliferation of neural stem cells in vivo
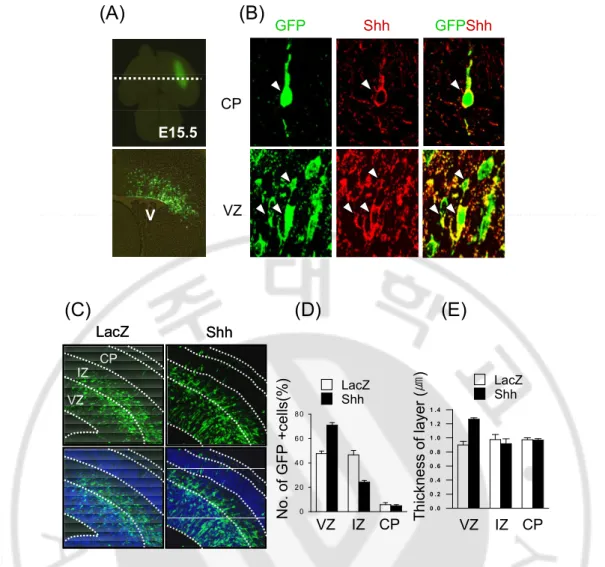
Shh is important factor about proliferation and survival factor of neural stem cells and concerned proliferation and differentiation of Nestin-expressing neural stem cells in hippocampus and cerebellum (Palma and Ruiz i Altaba 2004; Palma, Lim et al. 2005; Galvin, Ye et al. 2007). Previous data found that Shh induces proliferation of Nestin expressing cells in P19 cells. I investigated whether Shh is an essential factor about proliferation or survival of neural stem cells in vivo using in utero electroporation. At E13.5, I was transfected Shh plasmid including IRES-GFP into ventricular zone of dorsal cortex that is not Shh signaling (Fig. 8A). I showed that GFP positive cells co-expressed with Shh positive cells (Fig. 8B) and Shh was increased number of GFP positive cells in VZ but decreased in IZ compared with LacZ-transfected brain (Fig. 8C and D). The thickness increased in Shh-transfected VZ compared with lacZ-transfected VZ but not difference intermediated zone and cortical plate between LacZ and Shh-transfected brain (Fig. 8E). In Shh-transfected brain, neural stem cells markers such as Nestin and sox2 increased in ventricular zone compare with LacZ-transfected brain (Fig. 9A and B). Interestingly, Sox2 expression also increased in intermediated zone and cortical plate in Shh-transfected brain (Fig. 9B). Olig2 expression is known to regulate by Shh signaling in ventral region of cortex. The ectopic expression of Shh also was induced Olig2 expression in dorsal of cortex (Fig.10). These results indicate that Shh induces proliferation of neural stem cells and regulates Olig2 ectopic expression in dorsal cortex.
0 2 0 4 0 6 0 8 0 N o . o f G F P + c e lls (% ) LacZ Shh VZ IZ CP 0 . 0 0 . 2 0 . 4 0 . 6 0 . 8 1 . 0 1 . 2 1 . 4 T h ic k n e s s o f la y e r (㎛ ) LacZ Shh VZ IZ CP LacZ Shh VZ IZ CP LacZ Shh VZ IZ CP E15.5 V V GFPShh VZ CP Shh GFP
(A)
(B)
(C)
(D)
(E)
Fig. 8. The ectopic expression of Shh in developing dorsal cortex. Shh and LacZ were tranfected in ventricular zone of dorsal cortex at E13.5 using in utero electroporation. (A) EGFP was expressed in VZ of dorsal cortex. (B) GFP positive cells co-expressed with Shh in VZ and CP of dorsal cortex. (C) GFP-positive cells and thickness of layer in dorsal cortex were increased Shh compared with LacZ. (D, E) Quantitative analysis of GFP positive cells and thickness in Shh (filled) or LacZ (open)-tranfected brain. VZ, ventricular zone; SVZ, subventricular zone; IZ, intermediated zone; CP, cortical plate.
0 10 20 30 40 VZ IZ CP D e n s it y o f n e s ti n LacZ Shh 0 50 100 150 200 250 300 N 0 . o f S o x 2 + c e lls VZ IZ CP LacZ Shh GFPNestinHoechst GFPSox2Hoechst L a c Z S h h L a c Z S h h
(A)
(B)
Fig. 9. The effect of Shh on proliferation of neural stem cells in dorsal cortex of E15.5. (A, B) Immunohistochemistry of Nestin and Sox2 in dorsal cortex of Shh or LacZ-transfected brain. Quantitative analysis of Nestin density or Sox2+ cells in the dorsal cortex of Shh (filled) or LacZ (open) -transfected brain.
Contralateral A Contralateral A Ipsilateral B Ipsilateral B G F P R a t4 0 1 S o x 2 O lig 2 (A) (B) Contralateral C Contralateral C Ipsilateral D Ipsilateral D (C) (D)
Fig.10. The ectopic effect of Shh on neural stem cells in ventral cortex of E15.5. (A, B, C, D) Immunohistochemistry of Nestin, Sox2 and Olig2 in dorsal (A and B) and ventral cortex (C and D) of Shh-transfected brain.
6. Shh directly induces neuronal differentiation of P19 cells
To know effect of Shh in neuronal differentiation, I used new differentiation method from P19 cells into neuronal cells without the treatment of RA and aggregation culture for forming aggregate. This method is useful to study the function of Shh without the effect of RA that can induce neuronal differentiation of P19 cells. P19 and P19/Shh cells were cultured in DMEM:F12 with N2 supplement for 4 days. During differentiation, P19/Shh cells were appeared time dependently neuron like morphology (Fig. 10A). Shh increased expression of neuronal specific marker NF-M and Tuj1 in P19 cells (Fig. 11B-D) as well as neural stem cells marker, Nestin. Nestin expression started to appear at day 2 and increased at 4 days by Shh (Fig. 11B). But, glial markers such as GFAP and CNPase increased not during differentiation by Shh (Fig. 11D and data not shown). These results suggest that Shh can promote neuronal differentiation of P19 cells in serum-free media containing N2 supplement without RA.
Tuj1Hoechst P19 P19/ShhN std NF-M GFAP P19 P19 /Shh N GAPDH Nestin 0 2 4 0 2 4d P19 P19/Shh NF-M βIII-tubulin βIII-tubulin
(A)
(B)
(C)
(D)
Fig. 11. Shh induces neuronal differentiation of P19 cells. Cells were cultured to differentiate into neural cells in DMEM:F12 with N2 supplement for 4 days. (A) Neuron-like processes were increased in P19/Shh cells compared with P19 cells. (B) RT-PCR of Nestin, β-III-tubulin (Tuj1), and NF-M in P19 and P19/Shh cells. (C) Immunocytochemistry revealed expression of Tuj1 in P19/Shh cells but not in P19 cells. (D) Western blot analysis of NF-M, Tuj1, and GFAP in P19 and P19/Shh cells.
7. Shh promotes differentiation of P19 cells into motor and dopaminergic neurons. Shh has been known to promote differentiation from neural stem cells into various type neurons. Shh promotes motor neuron or oligodendrocyte differentiation in spinal cord and dopoaminergic neuron with FGF8 in midbrain. To know whether Shh can differentiate from P19 cells into these neurons, I was differentiated P19 cells in serum free medium supplement with N2 (induction medium) for indicated period of time as described previously with embryonic carcinoma cells (Darmon, Bottenstein et al. 1981; Pachernik, Bryja et al. 2005). Shh increased time dependently expression of MN specific marker, Nkx6.1, Olig2, Hb9 and ChAT as well as dopaminergic neuron markers, Nurr1 and TH in P19 cells (Fig. 12A and B). These results suggest that Shh promotes differentiation of motor and dopaminergic neuron in P19 cells.
1 2 3 4 P19 P19/ShhN 1 2 3 4 d Nkx6.1 Olig2 Ngn2 NeuroD GAPDH ChAT Hb9 GAPDH TH Nurr1 1 2 3 4 P19 P19/ShhN 1 2 3 4 d Nkx6.1 Olig2 Ngn2 NeuroD GAPDH ChAT Hb9 GAPDH TH Nurr1 a b P19 P19/ShhN T u j1 H b 9 T u j1 C h A T c d T u j1 T H
(A)
(B)
e fFig. 12. Shh induces differentiation of P19 cells into motor and dopaminergic neuron. Cells were cultured to differentiate into neural cells in DMEM:F12 with N2 supplement for 4 days. (A) Immunocytochemistry of motor neuron markers and dopaminergic neuron markers. (B) RT-PCR of motor and dopaminergic-related markers expression in P19 and P19/Shh cells.
8. Shh regulates the expression of Id and Hes1 during neural differentiation of P19 cells With evidence suggesting that N-myc plays an important role in the mitogenic response of P19 cells to Shh, I sought to identify possible downstream components that participate in P19 cells. Id proteins previously showed to be induced by the myc family members (Lasorella, Noseda et al. 2000; Light, Vernon et al. 2005; Swarbrick, Akerfeldt et al. 2005). N-myc proteins have been known to induce Id proteins that expresses in neural stem cells and can both promotes proliferation and inhibits differentiation of neural stem cells (Tzeng, Bresnahan et al. 2001; Bai, Sheng et al. 2007; Galvin, Ye et al. 2007). To know whether N-myc directly regulates Id proteins expression, I first analyzed expression of the Id proteins in P19 cells. Expression of Id1 and Id2 was higher in undifferentiated P19/Shh cells compared with P19 cells but not higher Id3 in undifferentiated P19 and P19/Shh cells (Fig. 13). During differentiation, the expression of Id proteins was decreased time dependently in P19 cells. However, in P19/Shh cells, Id proteins expression were slowly decreased compared with P19 cells. At that time, Nestin expression also increased by Shh. I also analyzed Id expression following enforced expression of siN-myc in P19 cells. Id2 expression was increased with Nestin expression by Shh in P19 cells. Shh-induced Id2 expression was decreased by siN-myc in P19 cells (Fig. 7C). Negative bHLH transcription factor Hes1 can inhibit neural stem cells from neurogenesis and neural differentiation through pro-neural gene as a neurogenin and mash1 (Nakamura, Sakakibara et al. 2000; Bai, Sheng et al. 2007). At 2days, Hes1 increased in P19/Shh cells compared with P19 cells (Fig. 13). These results suggest that Shh may regulate the expression of Ids and hes1 to promote proliferation of neural stem cells in early stage of neural development.
P19
P19/Shh
0
2
Id1
Id2
Id3
Nestin
Hes1
GAPDH
0
2 d
Fig. 13. Shh induces the expression of Id and Hes1 expresion in P19 cells. Cells were cultured in DMEM:F12 with N2 supplement. RT-PCR of Id1, Id2, Id3, Hes1 and Nestin expression in P19 and P19/Shh cells.
9. Id expression correlates with stemness of NSCs
To determine the role of Id proteins in NSCs, I first investigated the expression levels of Id1-3 during neuronal differentiation. NSCs were isolated from the E12.5 embryonic forebrain and grown as neurospheres in the presence of bFGP and EGF (Fig. 14A). The cells were expressed Nestin, a neural precursor cell marker, in neurospheres (Fig. 14A and B). In the absence of growth factors, these cells differentiated into neural cells that expressed Tuj1 and GFAP (Fig. 14A and B). RT-PCR analysis showed that Id, Id2, and Id3 were highly expressed in neurospheres but decreased in differentiated cells (Fig. 14B and C). These data indicate that expression of Id protein is tightly correlated to the proliferative stage of NSCs.
0 1 2 3 4 5 6 7 8
Id1 Id2 Id3 Nestin NFH GFAP
R e la ti v e R N A a m o u n t D if fe r. /u n d if fe r. ( F .I ) * * ** * * ** Undiffer. Differ. 0 1 2 3 4 5 6 7 8
Id1 Id2 Id3 Nestin NFH GFAP
R e la ti v e R N A a m o u n t D if fe r. /u n d if fe r. ( F .I ) * * ** * * ** Undiffer. Differ. Id1 Id2 Id3 nestin NF-H GFAP GAPDH Undiff. Diff. Tuj1 GFAP Id1 Id2 Id3 nestin NF-H GFAP GAPDH Undiff. Diff. Tuj1 GFAP Tuj1 GFAP
(A)
(B)
(C)
Fig. 14. Expression of inhibitor of differentiation (Id) correlates with proliferation status of neural stem cells (NSCs). (A) A phase contrast microscopic image of a neurosphere grown in the presence of bFGF and epidermal growth factor (EGF). A corresponding fluorescent image of the neurosphere stained with an anti-Nestin antibody. Cells were differentiated into β-tubulin III-positive neurons (green) or GFAP-positive astrocytes (red). (B) RT-PCR showed the expression of endogenous Id and Nestin genes decreased in differentiated cells at the expense of NF-H and GFAP. GAPDH was used as a control for equal loading. (C) The results were normalized to the value of GAPDH and shown as means ± S.E. with respect to the value in the presence of the growth factors (*P < 0.05; **P < 0.01).
10. Id proteins increase the proliferating neural precursors in the developing brain To confirm the roles of Id genes in neural precursor cells in vivo, I was overexpressed Id proteins in the developing forebrain using in utero electroporation on embryonic day 13.5. Id expression vectors, Id-IRES-GFP, contained the internal ribosome entry site (IRES) sequence (Pelletier and Sonenberg 1988) in front of the GFP sequence and allowed us to identify the transfected cells. At E15.5, I removed the embryonic brain expressing GFP and immunostained for the expression of Nestin (Zimmerman, Parr et al. 1994; Lothian, Prakash et al. 1999; Kawaguchi, Miyata et al. 2001) and Sox2 (Zappone, Galli et al. 2000; Bylund, Andersson et al. 2003), a NSC marker or Ki67 marker (Gerdes, Li et al. 1991), a proliferation. In Id-transfected brain, thickness of ventricular zone were increased compared with LacZ-trasnfected brain (Fig. 15A and B). But, in cortical plate, the layer decreased in Id-trasnfected brain compared with LacZ-transfected brain (Fig. 15A and B). The density of Nestin expression in the hemispheres electroporated with Id-IRES-GFP plasmids was increased in VZ and IZ (Fig. 15 C and D). The number of Ki67-positive cells in the hemispheres electroporated with Id-IRES-GFP plasmids were increased in VZ and SVZ; 13% ± 2.9% for Id1, 17% ± 2.1% for Id2, 13% ± 2.8% for Id3 (Fig. 16 A and B); in the intermediate zone (IZ) 5% ± 1.7% for Id1, 5% ± 2.1% for Id2, 5% ± 2.9% for Id3 (Fig. 16 A and B). In contrast, Ki67-positive cells were not found in IZ of the control animal brain (Fig. 16 A and B). In the ventricular zone (VZ), most cells expressed Sox2 (Fig. 16C and D), and the expression of Id proteins did not further increase the Sox2 immunoreactivity (Fig. 16 C and D). Overexpression of Id proteins increased the number of Sox2- positive cells in the subventricular zone (SVZ). The number of Sox2-positive cells among transfected cells was
14% ± 11.3% for Id1, 26% ± 10.7 % for Id2, and 10% ± 0.5% for Id3 (Fig.16C and D). Such Sox2-positive cells were not found in the SVZ of control embryos that were electroporated with LacZ-IRES-GFP (Fig.16C and D). Taken together, these results demonstrate that Id proteins induce proliferation of NSCs in the developing brain.
0 50 100 150 200 250 T h ic k n e s s o f la y e r (㎛ ) VZ IZ CP * * * * * * 0.0 0.5 1.0 1.5 2.0 2.5 3.0 VZ IZ D e n s it y o f n e s ti n + ( F .I ) LacZ Id1 Id2 Id3 LacZ Id1 Id2 Id3 VZ SVZ IZ CP VZ GFPNestin
LacZ Id1 Id2 Id3
VZ SVZ IZ CP LacZ Id1 Id2 Id3 LacZ Id1 Id2 Id3
(A)
(B)
(C)
(D)
Fig. 15. Id proteins increase population of neural stem cells in ventricular zone of dorsal cortex. Mouse telencephalic vesicles (E13.5) were electroporated with LacZ-IRES-GFP or Id-IRES-LacZ-IRES-GFP plasmids. (A and B) The thickness of each layer in Ids and LacZ transfected brain of E15.5. (C) Immunohistochemistry of Nestin expression in Ids and LacZ transfected brain. (D) The density of Nestin expression increased by Ids protein.
0 5 10 15 20 K i6 7 + /G F P + c e lls (% ) VZ/SVZ IZ * * * * * * 0 5 10 15 20 K i6 7 + /G F P + c e lls (% ) VZ/SVZ IZ * * * * * * LacZ Id1 Id2 Id3 LacZ Id1 Id2 Id3 IZ C P V Z
LacZ Id1 Id2 Id3
S V Z IZ IZ C P V Z V Z S V Z S V Z 0 20 40 60 80 100 120 S o x 2 + /G F P c e lls (% ) VZ SVZ * * * LacZ Id1 Id2 Id3 LacZ Id1 Id2 Id3