348 책임저자:최은아, 676-805, 경남 함양군 함양읍 용평리 595-47
인산죽염촌(주)
Tel: 055-964-0566, Fax: 055-964-0399 E-mail: [email protected]
접수일:2011년 12월 1일, 1차 수정일:2011년 12월 6일, 2차 수정일:2011년 12월 12일, 게재승인일:2011년 12월 15일
Correspondence to:Eun-A Choi
Insan Bamboo Salt Inc., 595-47, Yongpeong-ri, Hamyang-eup, Ham- yang 676-805, Korea
Tel: +82-55-964-0566, Fax: +82-55-964-0399 E-mail: [email protected]
LPS에 의해 활성화된 BV2 Microglia로부터 유도되는 염증매개물질의 생성에 미치는 사리장의 효과
동의대학교 1한의과대학 생화학교실 및 한의학연구소, 2대학원 바이오물질제어학과 및
블루바이오소재개발센터, 3부산대학교 약학대학 약학과, 4인산죽염촌(주), 5인산한의원
박혜영1,3ㆍ최영현1,2ㆍ최은아4,5
Anti-inflammatory Effects of Sarijang in Lipopolysaccharide-Activated BV2 Microglial Cells
Hye Young Park1,3, Yung Hyun Choi1,2 and Eun-A Choi4,5
1Department of Biochemistry, College of Oriental Medicine and Research Institute of Oriental Medicine, 2Department of Biomaterial Control (BK 21 Program), Graduate School and Blue-Bio Industry Regional Innovation Center, Dongeui
University, Busan 614-052, 3Department of Pharmacy, Pusan National University, Busan 609-735,
4Insan Bamboo Salt Inc., 5Insan Oriental Medical Clinic, Hamyang 676-805, Korea
Because of chronic microglial activation endangers neuronal survival through the release of various toxic pro-inflammatory molecules, negative regulators of microglial activation have been identified as potential therapeutic candidates in many neurological diseases. In this study, we investigated the inhibitory effects of the sarijang, an aged bamboosalt sauce containing extracts of Rhynchosia nulubilis, sulfur fed duck, Ulmus davidiana roots, and Allium sativum on the production of lipopolysaccharide (LPS)-induced pro- inflammatory mediators in BV2 microglia, such as nitric oxide (NO), prostaglandin E2 (PGE2) and pro-inflammatory cytokines. Sarijang significantly inhibited the excessive production of NO and PGE2
in LPS-stimulated BV2 microglia. It also attenuated expression of inducible nitric oxide synthase (iNOS), cyclooxygenase (COX-2), and pro-inflammatory cytokines such as interleukin-1β(IL-1β) and tumor necrosis factor (TNF)-α. Moreover, sarijang exhibited anti-inflammatory properties by suppressing nuclear factor-kappaB (NF-κB) activation, and downregulating the p38 mitogen-activated protein kinase (MAPK) and extracellular-signal-regulated kinase (ERK) pathways. Our results indicate that sarijang has a potent suppressive effect on the pro-inflammatory responses of activated BV2 microglia. This finding suggests that sarijang may offer substantial therapeutic potential for the treatment of neurodegenerative diseases that are accompanied by microglial activation. (Cancer Prev Res 16, 348-357, 2011)
Key Words: Sarijang, Bamboosalt, Microglia, NO, PGE2, Cytokines
서 론
Microglia (소교세포)는 중추신경계(central nervous sys-
tem, CNS)에서 조직 손상에 대해 초기 대응에 관여하는 중요한 세포이다. 하지만 비정상적으로 과다 활성화된 microglia는 nitric oxide (NO) 또는 superoxide, pro-in- flammatory factors interleukin-1β(IL-1β), tumor necrosis fac-
tor-α (TNF-α), monocyte chemoattractant protein (MCP)-1 등과 같은 chemokines와 inducible nitric oxide synthase (iNOS) 및 cyclooxygenase (COX)-2를 포함하는 세포독성 및 염증 유도 인자들의 발현과 생성을 촉진하여 만성염 증성질환 발달에 기여하게 된다.1~4) 특히 microglia는 lip- opolysaccharide (LPS), β-amyloid, thrombin, interferon (IFN)- γ 등에 의해 활성화 되고, 비정상적으로 활성화된 mi- croglia에 의한 염증 매개물의 분비는 면역계의 항상성을 교란시켜서 다발성 경화증(multiple sclerosis, MS), 알츠하 이머병(Alzheimer's disease, AD), 파킨슨병(Parkinsonism di- sease, PD) 등과 같은 중추신경계 자가면역질환 관련 퇴 행성 질환을 유발 및 진전시킨다.5,6) 따라서 microglia의 과다 활성화에 따른 선천 면역계 조절 장애를 이해하고 비정상적으로 활성화된 microglia로부터 분비되는 염증 매개물을 조절하는 것은 만성 염증성 질환의 진행을 늦 추거나 손상에 대한 치료에 접근 방법이 될 수 있다.
사리장은 해독성이 강한 서목태(Rhynchosia nulubilis, 鼠 目太, 일명 쥐눈이콩)를 발효시켜 유황오리, 유근피 및 마늘을 달인 액에 죽염을 혼합하여 숙성시킨 죽염간장 으로 해독성과 항암 및 소염성이 우수할 것으로 추정된 다(대한민국 등록특허 10-0496531). 이러한 약리 효능이 있을 것으로 추정하는 이유는 사리장의 주 원료인 죽염 이 항염증 및 항암 효능이 뛰어난 것으로 알려져 있고,7,8) 서목태는 일반 콩에 존재하는 isoflavone인 genistein, daid- zin, genistein, daidzein 등뿐만 아니라 배당체와 결합되지 않은 aglycone 함량이 우수하여 항암, 항염증 면역증강 효 능 뿐만 아니라 폐경기 골다공증 예방에 유효할 것이 며,9∼12) 유근피는 느릅나무(Ulmus davidiana var. japonica) 뿌리 껍질을 말린 것으로서 세포독성 보호, 항염증, 골다 공증 예방, 천식 예방 및 면역증강 효능이 뛰어난 것으로 알려져 있기 때문이다.13∼16) 황 함유 화합물을 대량 함유 하고 있는 마늘이나,17∼20) 유황오리 추출액 역시 항염증, 면역 증강, 항암 및 암예방 효능 등이 우수한 것은 이미 잘 알려진 바 있다.21∼23) 현재까지 사리장의 구성성분에 대한 분석은 이루어지지 않았지만, 사리장의 주요 구성 원료들에 대한 선행 자료 결과들로 미루어 사리장에는 다양한 성분들이 혼재되어 있을 것으로 추정된다. 따라 서 사리장의 약리효능과, 그와 연관된 지표 물질의 탐색 을 위한 연구의 일환으로 사리장의 항염증 효능 여부를 우선적으로 조사하였다. 이를 위하여 전향적인 항염증 연구 모델로 널리 사용되고 있는 마우스의 뇌신경교세 포인 BV2 microglia에서 LPS에 의하여 유도되는 염증성 반응에 미치는 사리장의 효능을 조사한 결과, 유익한 결 과를 얻었기에 이를 보고하는 바이다.
재료 및 방법 1. 실험 재료 및 세포배양
본 실험에 사용된 시약과 항체들은 Sigma-Aldrich Che- mical Co. (St Louis, MO, USA), R&D Systems (Minneapolis, MN, USA), Bio-Rad Laboratories (Hercules, CA, USA), Invitrogen Life Technologies (Carldbad, CA, USA) 및 Santa Cruz (Santa Cruz,CA, USA)로부터 구입하여 사용하였다.
사리장은 인산죽염촌(주)에서 제공 받아서 0.4μl의 sir- inge filter로 여과 멸균하여 4oC 보관하며 사용하였다.
Murine microglia cell line인 BV2 세포는 인제대학교 의과 대학에서 분양받았으며, 10% fetal bovine serum (FBS)과 1% antibiotics (penicillin/streptomycin)를 첨가한 DMEM 배 지를 이용하여 5% CO2, 37oC incubator에서 주 3회 계대 배양하여 사용하였다.
2. MTT assay를 이용한 세포독성조사
실험 조건 설정을 위한 사리장의 세포독성(cytotoxicity) 여부를 확인하기 위해서 BV2 세포(3×105 cells/well)를 6-well culture plate에 24시간 동안 배양한 후 다양한 농도 의 사리장을 1시간 전 처리한 뒤 500 ng/ml의 LPS를 추가 적으로 24시간 처리하여 배양하였다. 24시간 후 배지를 제거하고 3-(4,5-dimethy;lthazol-2-yl-2,5-teterazolium bromi- de (MTT, Sigma-Aldrich)를 0.5 mg/ml 농도로 첨가하여 2시 간 동안 배양하면서 환원반응을 유도한 후 formazan 결정 을 dimethylsulfoxide (DMSO) 용액으로 용해한 후, micro- plater Reader (Molecular Devices, Sunnyvale, CA, USA)를 이 용하여 흡광도(540 nm)를 측정하였으며, LPS 및 사리장 이 처리되지 않은 조건에서 배양한 대조군을 기준으로 처리군의 세포생존율(%)을 계산하였다.
3. Nitric oxide (NO)의 측정
LPS에 의해 활성화된 BV2 세포로부터 생성되는 NO의 농도를 세포 배양액에 존재하는 NO2
−를 인지하는 Griess 시약을 이용하여 검출하였다. 이를 위하여 BV2 세포(5×
105 cells/well)를 6-well culture plate에 24 시간 동안 배양한 후 다양한 농도의 사리장을 1시간 전 처리한 뒤 LPS (500 ng/ml)를 다시 처리하였다. 24 시간 배양한 뒤 세포 배양 액을 수거하고 배양액에 동량의 Griess 시약(0.1% naph- thylethylenediamine & 1% sulfanilamide in 5% H3PO4)을 넣 어 실온에서 반응 시켰다. 반응액을 microplate reader를 이용하여 540 nm 흡광도에서 측정하였으며, 세포로부터 생성된 NO의 양은 NaNO2 표준액의 농도를 기준(stan-
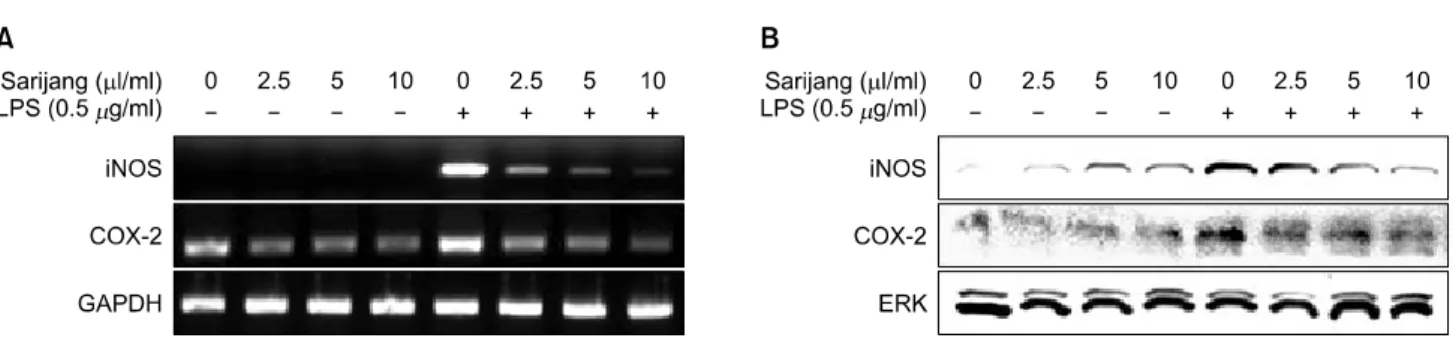
Table 1. Sequence of primers used for RT-PCR
Gene name Sequence
iNOS Sense
Antisense
5'-AAG-CAC-ATG-CAG-AAT-GAG-TAC-CG-3' 5'-GTG-GGA-CAG-CTT-CTG-GTC-GAT-3'
COX-2 Sense
Antisense
5'-GGA-GAG-ACT-ATC-AAG-ATA-GTG-ATC-3' 5'-ATG-GTC-AGT-AGA-CTT-TTA-CGA-CTA-3'
TNF-α Sense
Antisense
5'-CCC-CTC-AGC-AAA-CCA-CCA-AGT-3' 5'-CTT-GGG-CAG-ATT-GAC-CTC-AGC-3'
IL-1β Sense
Antisense
5'-AAT-CTC-ACA-GCA-GCA-CAT-CAA-3' 5'-AGC-CCA-TAC-TTT-AGG-AAG-ACA-3'
GAPDH Sense
Antisense
5'-ACC-ACA-GTC-CAT-GCC-ATC-AC-3' 5'-TCC-ACC-ACC-CTG-TTG-CTG-TA-3'
dard curve)으로 계산하였다.
4. Prostaglandin E2 측정
LPS 자극에 의하여 BV2 세포로부터 생성되는 prosta- glandin E2 (PGE2)의 양을 측정하기 위하여 동일한 조건에 서 배양된 세포 배양액을 수거하여 PGE2 enzyme immuno- assay (EIA) kit (Cayman Chemical Company, Ann Arbor, MI, USA)를 이용하여 PGE2의 생성량을 측정하였으며, 세포 배양액 내 존재하는 PGE2의 농도(pg/ml)는 표준용액의 농도를 기준으로 환산하여 계산하였다.
5. 사이토카인 측정
사리장의 염증성 사이토카인 생성억제 효과를 조사하 기 위해서 BV2 세포로부터 생성되는 TNF-α와 IL-1β의 양을 enzyme-linked immunosorbent assay (ELISA) kit (R&D Systems, Minneapolis, MN, USA)를 사용하여 세포배양액에 서 측정하였다. 이를 위하여 1× assay diluent 용액을 50μl 넣은 뒤 세포 배양액 또는 각 사이토카인의 표준단백질 을 50μl씩 넣어 2시간 동안 실온에서 반응시켰다. 이를 5회 washing buffer로 세척한 다음 detection antibody를 넣 어 실온에서 2시간 동안 반응시켰다. 다시 washing buffer 로 5회 세척하고 color reagent 용액을 100μl 넣어 실온에 서 30분간 암 반응시킨 후 반응 정지액을 100μl 첨가하 였다. Plate의 반응 정도를 microplate reader 에서 450 nm 흡광도로 측정하였다. BV2 세포로부터 분비된 각 사이 토카인의 농도는 표준액의 표준정량곡선을 기준으로 계 산하였다.
6. Reverase Transcriptase (RT)-PCR
사리장이 iNOS, COX-2 및 염증사이토카인들의 발현 에 미치는 효과를 전사 수준에서 조사하기 위해 RT-PCR 을 수행하였다. 다양한 조건에서 배양된 BV2 세포로부
터 TRIzol 시약(Invitrogen, CA, USA)을 이용하여 total RNA 를 분리하였다. 동량의 RNA를 oligo (dT)15 primer를 사용 해 cDNA로 역전사시켰다. PCR은 cDNA와 각 primer (Table 1)를 이용하였다. PCR을 위해 94oC에서 5분간 가열 한 후, 94oC에서 1분, 55oC에서 1.5분, 72oC에서 1분의 순 서로 30회 반복하고, 이후에 72oC에서 5분간 추가 단계 를 거쳤다. PCR 생성물들은 2% agaros gel을 사용하여 전 기영동시키고 ethidium bromide (EtBr)를 이용하여 염색하 였다.
7. 면역형광염색
준비된 세포를 0.01 M phosphate buffered saline (PBS, pH 7.4)으로 두 차례 세척한 후, 4% paraformaldehyde로 4oC에 서 15분 동안 고정시켰다. 다시 PBS로 세척한 다음 0.3%
Triton X-100이 함유된 PBS로 4oC에서 20분 동안 반응시 킨 후, 2% 소혈청알부민이 함유된 PBS로 실온에서 1시 간 반응시켜 비특이적 반응을 억제하였다. 다시 anti-p65 항체를 2% BSA가 함유된 PBS용액에 1:200으로 희석하 여 실온에서 2시간 반응시켰다. PBS로 세 차례 세척한 후 2차 항체인 fluorescein isothiocyanate (FITC)-conjugate IgG (Molecular Probe, Eugene, OR, USA)를 1:100으로 희 석하여 실온에서 1시간 동안 반응시켰다. 면역형광염색 된 세포를 표본을 공초점 주사현미경(FV300. Olympus, Japan)을 사용하여 관찰하였다. 레이저 광선의 파장으로 488 nm을 FITC용으로 사용하였으며, flow view softwave program (Olympus, Japan)을 사용하여 최종 3차원적 영상 으로 재조립하였다.
8. Western blot analysis
사리장이 iNOS, COX-2, MAPKs, IκB와 NF-κB의 단 백질 발현에 미치는 영향을 조사하기 위해 Western blot- ting을 수행하였다. 이를 위하여 준비된 BV2 세포를
Fig. 1. Effect of sarijang on the cell viability of BV2 microglia.
BV2 cells (3×105 cells/ml) were treated with the indicated concentrations of sarijang (0, 2.5, 5, or 10μl/ml) for 1 h before LPS (500 ng/ml) treatment for 24 h. Cell viability was assessed by MTT reduction assay and the results are expressed as percentage of surviving cells over control cells (no addition of sarijang). Results are expressed as means±S.D. from three independent experiments.
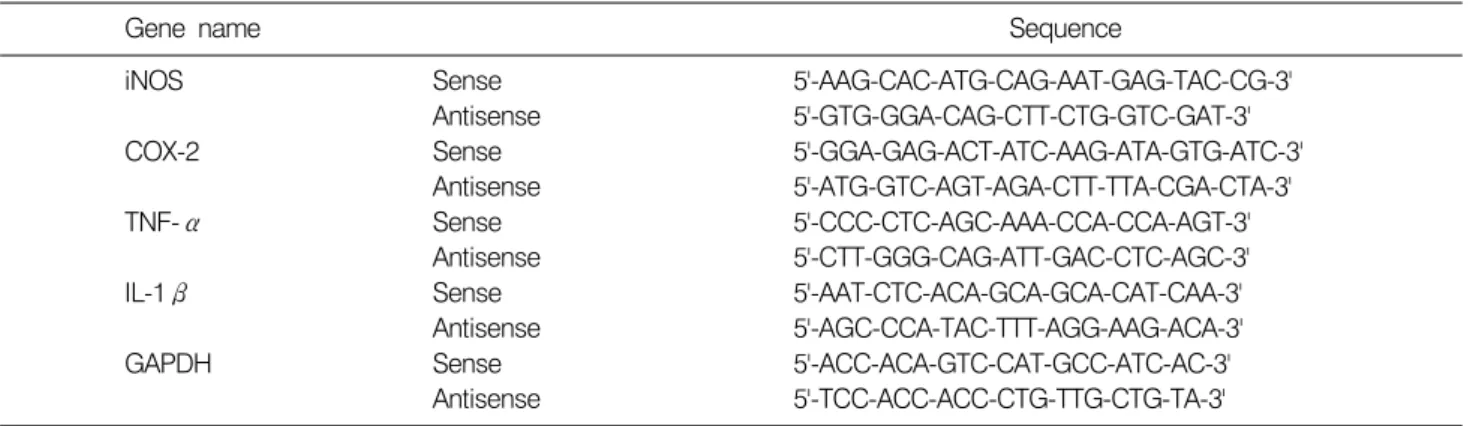
Fig. 2. Inhibitory effect of sarijang in LPS-induced NO and PGE2 production in BV2 microglia. Cells were treated with the indicated concentrations of sarijang (0, 2.5, 5, or 10μl/ml) 1 h before LPS (500 ng/ml) treatment for 24 h. Control values were obtained in the absence of LPS or sarijang. (A) The amounts of NO were determined using Griess reagent and a standard curve created using NaNO2 in culture medium. (B) The levels of PGE2 in the media were detected using a specific enzyme immunoassay according to the manufacturer’s instruction. Results are expressed as means±S.D. from three independent experiments. *p<0.05 indicates a significant difference from cells treated with LPS in the absence of sarijang.
ice-cold PBS로 두 번 세척한 다음 lysis buffer (1 M Tris-HCl, 5 M NaCl, 0.5% Triton X-100, protease inhibitor cocktail)를 이용하여 세포를 lysis하였다. 이를 14,000 rpm에서 15분 간 원심분리하여 상층액을 회수하고 Bradford 용액과 BSA 표준용액을 이용하여 단백질의 양을 정량한 후 50 μg의 단백질을 9∼12% polyacrylamide gel을 이용하여 전 기영동 하였다. Gel을 분리하여 nitrocellulose membrane에
transfer시킨 다음 5% skim milk로 실온에서 1시간 동안 blocking시키고, 각 표적 단백질들에 대한 항체를 첨가하 여 4oC에서 하룻밤 반응시킨 후 PBS-T 용액으로 3회 세 척한 다음 anti-IgG conjugated HRP 항체를 넣어 실온에서 1시간 반응시켰다. 이를 다시 PBS-T로 3회 세척하고 en- hanced chemiluminescence (ECL) Western blotting detection reagents (SuperSignal, Thermo Scientific, Rockford, IL, USA) 용액을 이용하여 X-ray film에 감광시켰다. 각 단백질의 발현에 대한 대조군으로 ERK 및 LaminB에 대한 항체를 이용하여 위와 동일한 방법으로 수행하였다.
9. 통계 처리
모든 실험결과는 평균±표준편차로 표시하였고 Sigma- Plot을 이용하여 Student t-test를 이용하여 통계적 유의성 을 얻었다.
결 과 1. 세포독성 검정
사리장의 세포독성 여부를 조사하기 위해서 BV2 세포 에 다양한 농도(0, 2.5, 5 및 10μl/ml)의 사리장을 단독 및 LPS와 동시에 처리하여 24시간 배양한 후 MTT assay를 수행하였다. BV 세포에 대한 사리장의 세포독성은 사리 장을 단독으로 배양한 대조군의 세포생존율 100%를 기 준으로 2.5μl/ml, 5μl/ml 및 10μl/ml 농도의 사리장 처리 군에서 각각 101.6%, 94.3% 및 93%로 나타났다(Fig. 1).
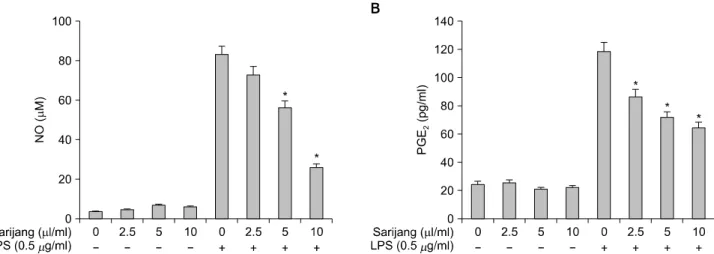
Fig. 3. Inhibitory effect of sarijang in LPS-induced NO and PGE2 expression in BV2 microglia. (A) BV2 microglia were pretreated for 1 h with sarijang prior to stimulation with LPS. After 24 h, total RNAs were isolated and reverse-transcribed using iNOS and COX-2 primers. Resulting cDNAs were then subjected to PCR and the reaction products were subjected to electrophoresis in a 1% agarose gel and visualized by EtBr staining. GAPDH was used as an internal control. (B) Cells grown under the same conditions as (A) were lysed and equal amounts (30 lg) of cell lysate were resolved by SDS–.polyacrylamide gel electrophoresis and transferred to nitrocellulose. Western blotting was then performed using anti-iNOS and COX-2 antibodies, and an ECL detection system. ERK was used as an internal control.
또한 동일 조건에서 LPS를 동시에 처리하였을 경우에도 이와 유사한 결과를 보여 사리장은 2.5∼10μl/ml 농도 범위 내에서 단독 및 LPS와 동시 처리 조건에서 유의적 인 세포독성을 나타내지 않는 것으로 나타났다. 따라서 이후 실험은 독성이 없는 상기 농도 범위에서 수행하였 다.
2. LPS로 활성화된 BV2 세포에서 사리장에 의한 NO 및 PGE2의 생성 억제 효과
사리장이 LPS에 의해 활성화된 BV2 세포로부터 염증 매개 물질인 NO의 생성 억제 효과를 확인하기 위해 Griess 반응법으로 세포배양액 내 NO의 농도를 측정하였 다. 이를 위하여 BV2 세포에 LPS (500 ng/ml) 처리로 NO 의 발생을 유도하고, 사리장을 처리하여 NO 발생 변화 를 확인하였다. 그 결과, LPS 자극이 주어지지 않은 BV2 세포에서는 3.9μM 정도로 낮은 농도의 NO가 검출된 반면 LPS를 처리하였을 때 87.59μM의 NO가 검출되어 LPS 처리에 의해 BV2 세포가 활성화되었음을 확인하였 다(Fig. 2A). 사리장을 0, 2.5, 5 및 10μl/ml로 선 처리한 결과 LPS에 의해 유도된 NO의 생성이 사리장 농도 의존 적으로 76.1μM, 58.8μM 및 27.2μM로 감소되었음을 확인 할 수 있었다. 아울러 사리장이 BV2 세포로부터 생 성되는 또 다른 염증매개 물질인 PGE2를 효과적으로 억 제할 수 있는지 EIA 반응법으로 측정하였다. LPS 자극이 주어지지 않은 BV2 세포에서는 낮은 농도의 PGE2가 검 출된 반면 LPS 처리에 의해 배양액 속에 축적되어있는 PGE2의 양이 현저히 증가되는 것을 확인하였다(Fig. 2B).
그러나 사리장을 각각 0, 2.5, 5 및 10μl/ml 농도로 처리 하였을 때 농도 의존적으로 PGE2의 생성이 매우 감소되
었음을 알 수 있었다.
3. LPS로 활성화된 BV2 세포에서 사리장 처리에 의한 iNOS 및 COX2 발현 억제 효과
이상의 결과에서 사리장의 처리에 의하여 LPS에 의한 NO 및 PGE2의 생성이 현저하게 억제되었기에, 사리장 처리가 NO의 합성효소인 iNOS와 PGE2 생성 주요 효소 인 COX-2의 발현에 미치는 영향을 RT-PCR과 Western blotting으로 조사하였다. 먼저 iNOS의 경우, 정상 조건에 서 배양된 BV2 세포와 사리장만을 단독으로 처리한 세 포에서 전사 및 번역 수준에서의 발현이 매우 낮게 관찰 된 반면, LPS (500 ng/ml)로 자극한 BV2 세포에서는 mRNA 및 단백질의 발현이 매우 높게 증가되었다(Fig. 3).
그러나 LPS에 의해 유도된 iNOS의 mRNA의 발현뿐만 아 니라 단백질의 발현도 사리장 처리 농도 의존적으로 현 저히 억제되었다. COX-2의 경우도 iNOS와 유사하게, LPS에 의하여 매우 강하게 발현이 증가되었으나, 사리장 의 처리에 의하여 전사 및 번역 수준에서 모두 현저히 억제되었음을 알 수 있었다. 즉 사리장에 의한 LPS 처리 로 유도된 NO 및 PGE2의 생성 억제 효과는 iNOS와 COX-2의 발현 억제와 연관이 있으며, 이들 유전자의 발 현 억제가 전사수준에서 이루어지고 있음을 알 수 있었 다.
4. LPS로 활성화된 BV2 세포에서 사리장 처리에 의한 TNF-α 및 IL-1β의 생성 억제 효과
다음은 염증 촉진인자로 작용하는 cytokine의 생성에 미치는 사리장의 효과를 조사하기 위하여 다양한 cyto- kine 중, TNF-α와 IL-1β의 생성에 미치는 영향을 조사
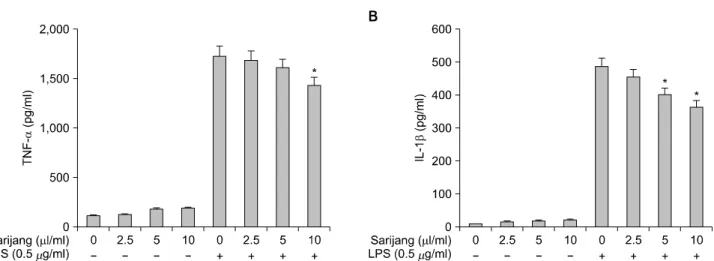
Fig. 4. Inhibitory effect of sarijang in LPS-induced pro-inflammatory cytokines in BV2 microglia. Cells were pretreated with the indicated concentrations of sarijang for 1 h before LPS treatment. After incubation for 24 h, the supernatants were taken, and the amounts of TNF-α (A) and IL-1β (B) were measured. Values are expressed as means±S.D. from three independent experiments. *p<0.05 indicates a significant difference from cells treated with LPS in the absence of sarijang.
Fig. 5. Inhibitory effect of sarijang in LPS-induced TNF-α and IL-1β expression in BV2 microglia. BV2 microglia were pre- treated for 1 h with sarijang prior to stimulation with LPS. After 6 h, total RNAs were isolated and reverse-transcribed using TNF-α and IL-1β primers. Resulting cDNAs were then sub- jected to PCR and the reaction products were subjected to electrophoresis in a 1% agarose gel and visualized by EtBr staining. GAPDH was used as an internal control.
하였다. 이를 위하여 BV2 세포에 LPS (500 ng/ml)를 단독 으로 처리한 후, TNF-α와 IL-1β 생성 변화를 ELISA 방 법으로 먼저 확인하였다. Fig. 4에 나타낸 바와 같이, 정 상 조건에서 배양된 BV2 세포에서 TNF-α와 IL-1β는 각각, 118.6 p/ml과 32.4 pg/ml 정도의 낮은 수준으로 검출 되었으나, LPS 단독 처리군에서 TNF-α와 IL-1β의 생성 이 각각 2049.7 pg/ml 및 463.1 pg/ml로 현저히 증가되었 다. 그러나 사리장은 LPS에 의해 증가된 TNF-α와 IL-1 β의 생성을 어느 정도 억제하였으며, 이러한 억제의 정 도는 사리장에 의한 TNF-α와 IL-1β 유전자의 발현 억 제 정도와 유사한 경향성을 보여주었다(Fig. 5).
5. LPS로 활성화된 BV2 세포에서 사리장 처리가 NF- κB 활성화에 미치는 영향
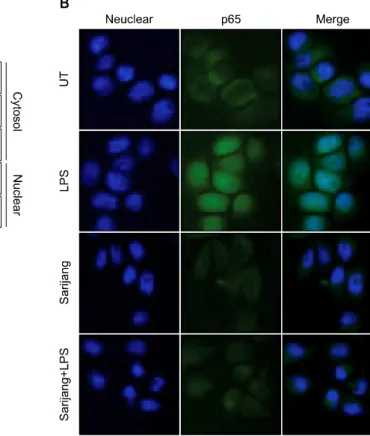
염증 반응 매개에 관여하는 중요 전사인자인 nuclear transcription factor-kappa-B (NF-κB)와, NF-κB의 negative regulator로 알려진 I-κBα의 발현에 미치는 영향을 조사 하였다. 이를 위하여 핵 단백질과 세포질 단백질을 분리 하여 두 유전자의 발현을 단백질 수준에서 비교하였으 며, 면역형광염색법을 이용하여 NF-κB 단백질의 세포 내 분포 양상을 비교하였다. 먼저 Fig. 6A의 결과에서 알 수 있듯이, LPS 처리군에서 NF-κB p65 subunit의 발현이 세포질 내에서 현저히 감소되었으며, 핵 내에서의 발현 이 증가되어 LPS 처리에 의하여 NF-κB가 핵 내로 이동 되었음을 알 수 있었으나, I-κBα의 경우 NF-κB와는 반대로 LPS 처리에 의하여 세포질에서의 발현이 감소되 었다. 그러나 사리장 동시 처리군에서는 LPS에 의한 NF- κB의 핵으로의 이동이 현저하게 억제되었다. 또한 NF- κB의 핵으로의 이동에 대한 추가적인 자료를 제시하기 위하여 면역형광염색법을 이용한 결과에서도, LPS로 자 극되지 않은 정상 조건에서 배양된 BV2 세포의 NF-κB 의 발현은 대부분 세포질에서 나타난 반면, LPS로 활성 화된 BV2 세포는 p65가 핵에서 발현이 증가하였다. 그러 나 사리장은 LPS 처리에 의한 NF-κB의 핵 내 이동이 현저하게 억제됨을 알 수 있었다(Fig. 6B). 이러한 결과는 사리장 처리가 LPS에 의한 NF-κB 단백질의 핵 내 이동 을 억제함으로써 LPS에 의해 유도되는 주요 염증신호전 달경로인 NF-κB pathway를 차단할 수 있음을 의미한다.
Fig. 6. Effect of sarijang on NF-κB activation in LPS-stimulated BV2 microglia. BV2 cells were pretreated for 1 h with the indicated concentrations of sarijang prior to stimulation with LPS for the indicated times. (A) The cytosolic and nuclear proteins were isolated and subjected to 9% SDS-PAGE followed by Western blotting using anti-NF-κB p65 and IκB-α antibodies, and an ECL detection system. ERK and lamin B were used as an internal control for the cytosolic and nuclear fractions. respectively. (B) In a parallel experiment, the cells were pretreated with 10μl/ml sarijang for 1 h, and then stimulated with LPS (500 ng/ml) for 60 min.
Localization of NF-κB p65 was visualized with a fluorescence microscope after immunofluorescence staining with anti-NF-κB p65 antibody (green). Cells were aslo stained with DAPI to visualize the nuclei (blue). Results are representative of those obtained from three independent experiments.
6. LPS로 활성화된 BV2 세포에서 사리장 처리가 MAPKs 활성에 미치는 영향
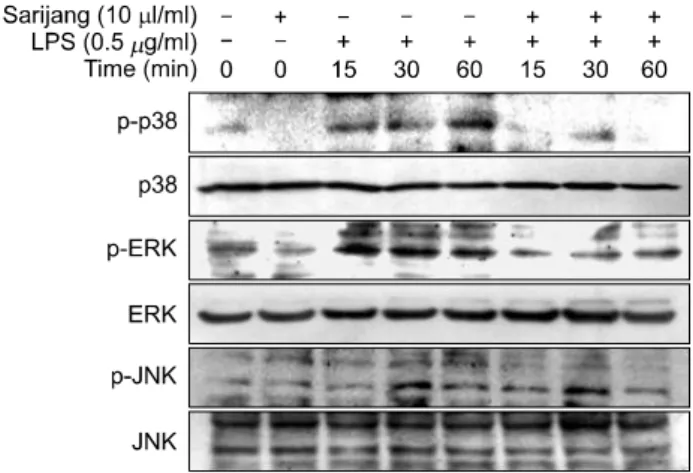
Mitogen activated protein kinases (MAPKs)는 세포의 증식 과 분화를 포함하는 다양한 면역반응을 조절하며, 특히 외부 감염에 의해 활성화되어 염증매개물질들의 생성을 유도하는데 중요한 역할을 하는 것으로 알려져 있다. 따 라서 LPS로 자극한 BV2 세포에 사리장 처리에 의한 염증 반응 억제 과정에 관여하는 MAPKs의 인자들을 발현 변 화를 추가적으로 조사하였다. Fig. 7의 결과에서 알 수 있듯이, LPS가 처리된 BV2 세포에서 p38MAPK, ex- tracellular signal-regulated kinase (ERK) 및 c-Jun N-terminal kinase (JNK)의 인산화(phosphorylation)가 전반적으로 강하 게 유도되었으며, 사리장 동시 처리군에서는 p38 MAPK 와 ERK의 인산화가 현저히 억제되는 것으로 나타났다.
이 결과 사리장의 항염증 효능이 p38MAPK와 ERK의 신
호 전달 억제를 통하여 이루어질 수 있음을 의미하는 것 이다.
고 찰
Microglia는 CNS에 존재하는 식균세포(phagocytic cell)로 서의 CNS의 초기 면역반응을 조절하는 고전적인 기능 외에 TNF-α, IL-1β 등과 같은 염증성 사이토카인이나 eicosanoids, 활성산소종(reactive oxygen species, ROS), NO, superoxide (O2−) 등을 과도하게 분비함으로 퇴행성 질환 의 발달에 기여한다.24~26) 따라서 비정상적으로 활성화 된 microglia로부터 생성되는 다량의 염증매개물질의 활 성 및 생성의 조절이 퇴행성 질환의 치료 표적인자로 인 식되고 있다.
세균내독소인 LPS는 염증을 유발하는 주요 inflam- matogen으로 microglia를 활성화하여 NO, PGE2, TNF-α,
Fig. 7. Effect of sarijang on MAPKs activation induced by LPS in microglia. BV2 cells were treated with the indicated con- centrations of sarijang 1 h before LPS treatment for indicated times. The cells were lysed, and cellular proteins were then separated by 9% SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. Membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system.
IL-1β, monocyte chemoattractant protein-1 (MCP-1), macro- phage inflammatory peptide-1 (MIP-1), IL-16, IL-18, macro- phage colony-stimulating factor (M-CSF) 등과 같은 다양한 염증성 매개물질들을 과도하게 분비하며, 이들은 세포 독성물질로 작용하여 염증반응을 가속화시킴으로 만성 질환 발달에 기여한다.27∼30) 따라서 활성화된 BV2 세포 로부터 생성되는 염증 매개인자의 생성 및 관련 유전자 의 발현 제어는 염증성 질환의 치료뿐만 아니라 다양한 질병의 예방과 치료에 매우 유용성이 높은 것으로 인식 되고 있다. 본 연구에서는 사리장의 항염증 효능을 확인 하고 관련 기전의 해석을 위하여 LPS에 의하여 유도되는 다양한 염증 매개인자들의 생성과 그들의 생성을 조절 하는 주요 유전자들의 발현 변화를 세포독성이 없는 범 위에서 조사하였다.
먼저 NO는 활성질소종의 일종으로 독성이 있는 저분 자 라디칼로서 신체 면역계의 방어기능에 관여하고 있 다.31) NO는 체내 대식세포에서 L-arginine으로부터 NO 합성효소인 c-NOS (constitutive NOS)와 iNOS (inducible NOS)를 통해 생성되는데, NO는 LPS, TNF-α, IFN-γ 또 는 IL-1β와 같은 염증성 자극을 받은 microglia가 생성해 내는 iNOS에 의해 생성된다. 생체 내 과도한 NO 분비는 세포 독성을 일으키고 염증반응을 촉진하며 중추신경계 에서 염증반응을 매개하여 퇴행성 뇌질환의 발병에 관 여하게 된다.31,32) 본 연구의 결과에 의하면 사리장은 LPS 처리에 의하여 활성화된 BV2 세포로부터 생성되는 NO
를 유의적으로 감소시켰으며(Fig. 2A), 이는 NO 합성효 소인 iNOS 유전자의 전사 단계를 억제하는 것으로 나타 났다(Fig. 3). 한편 특정 prostaglandin 계열 물질들은 점진 적인 염증 반응을 심화시키는 것으로 알려져 있는데,33,34) COX, lipoxygenase (LOX) 또는 cytochrome P-450 mono- ocygenase에 의해서 arachidonic acid로부터 PGE2, PGD2, PGF2α, PGI2, thromboxane A2 등의 5개의 중요한 prosta- glandin이 합성된다. 특히 PGE2는 가장 잘 알려진 염증 반응의 매개체로서 NO와 마찬가지로 다양한 염증질환 의 발달에 기여한다.28,35) 본 연구의 결과에 의한 NO의 결과에서처럼 사리장은 LPS에 의한 PGE2 생성 및 COX-2 의 발현 증가를 현저하게 억제시켰으며, 이 또한 전사 수준에서 조절하고 있음을 알 수 있었다(Fig. 2B 및 Fig.
3). 즉 사리장은 iNOS 및 COX-2의 발현을 전사 수준에서 억제하고 NO 및 PGE2의 생성을 억제함으로서 염증 반 응 억제 효과가 있음을 알 수 있었다.
Cytokine은 면역, 감염, 조혈기능, 조직회복, 세포의 성 장, 생체의 방어 등에 필수적인 역할을 하며, 다양한 질 병의 병태ㆍ생리적 반응을 매개하는 물질이다. 활성화 된 microglia는IL-1β, IL-6, IL-10, TGF-β 및 TNF-α 등과 같은 cytokime을 분비하는데, 이들은 정상 상태에서는 매 우 낮은 수준으로 발현되지만, 과도한 손상 상태에서는 높게 발현되어 염증 촉진 인자로 작용할 수 있다.36~38) 특히 TNF-α와 IL-1β는 다양한 퇴행성 질환과 관련된 대표적인 전구염증 사이토카인으로서 prostaglandins, leu- kotrienes, NO, plateletactivating factor 등의 매개물질들을 합성시킴으로서 염증질환을 유발하는 요인이 된다.39,40) 특히 이들의 발현이 뇌졸중이나 파킨슨병과 알츠하이머 병 환자들의 혈청과 뇌 조직에서 증가되어 있고, 뇌신경 세포의 사멸 등의 병인기전에 중요한 역할을 하는 것으 로 알려져 있다.40∼42) 본 연구에서 사리장이 LPS에 의해 활성화된 BV2 세포로부터 분비되는 TNF-α와 IL-1β의 생성과 유전자 발현에 미치는 영향을 조사한 결과, 사리 장은 두 물질의 생성과 그들 유전자의 발현을 전사수준 에서 유의적으로 억제시켰다(Fig. 4 및 5). 이는 사리장이 염증성 사이토카인의 유전자 발현단계 조절을 통하여 이들 사이토카인의 생성을 억제할 수 있음을 의미하며, 염증관련 뇌질환 개선 및 증상완화에 효과적일 수 있음 을 시사한다.
한편, 병원체의 감염에 의해 영향 받는 대식세포 신호 전달 분자로는 serine/threonin kinase로써 세포 밖 신호를 핵 내로 전달하게 하는 MAPKs가 중요한 역할을 담당한 다. MAPK의 subfamily에는 ERK, JNK 및 p38MAPK가 있 으며, MAPKs의 신호 전달과정은 염증반응을 유도하는
염증매개물질들의 합성을 유도한다. 병원체의 감염에 의해 시작되는 LPS에 의하여 활성화되는 염증반응은 MAPK 분자들을 인산화시켜 NF-κB, activator protein-1 (AP-1)과 같은 전사조절인자가 활성화 되어 NO 및 super- oxide anion 등과 같은 free radical들이 생성된다.43) 또한 NF-κB는 세포 분화, 염증반응, 세포 부착 등 다양한 세 포 활동을 조절하는 유전자들의 발현에 중대한 역할을 하는 전사인자이다.44) NF-κB는 proinflammatory cytokine 의 자극에 의해 세포질에 존재하던 NF-κB가 I-κB와의 분리되어 핵 내로 이동함으로서 표적 유전자의 promoter 부위에 있는 kB 결합 자리에 결합하여 염증에 관여하는 TNF-α, IL-1β, IL-6 등의 염증성 cytokine이나 면역세포 이동에 중요한 접합 분자들(cell adhesion molecule), chemo- kines 등과 같은 염증 매개물질의 전사를 촉진한다.45) 본 연구에서 사리장의 항염증 효과가 이들 유전자 산물의 활성 조절을 통하여 이루어지는지를 조사한 결과, LPS 처리에 의해 유도되는 p38MAPK와 ERK의 활성화를 억 제시켰으며(Fig. 7), 핵 내로의 NF-κB p65 subunit의 이동 을 억제시켰다(Fig. 6). 따라서 사리장에 의한 MAPKs와 NF-κB의 활성 억제는 염증사이토카인의 발현을 억제 하는 작용에 직접적인 영향을 미치는 것으로 보인다.
결론적으로 사리장은 LPS에 의해 활성화된 BV2 세포 로부터 유도되는 NF-κB 및 MAPKs 신호전달경로를 차 단함으로서 염증매개물질인 NO, PGE2 및 염증성 사이 토카인(TNF-α 및 IL-1β)의 생성을 억제하여 항염증 효 과를 나타내고 있음을 알 수 있었다. 따라서 사리장은 염증성 질환을 매개로 하는 각종 퇴행성뇌질환의 예방 및 치료제 개발에 활용할 수 있을 것으로 기대된다.
결 론
본 연구에서는 죽염과 서목태를 주원료로 제작된 죽 염간장의 일종인 사리장의 항염증 효능 및 관련 기전을 조사하였다. 이를 위하여 BV2 microglia 세포에서 LPS로 유도된 중요한 염증성 매개인자인 NO 및 PGE2의 생성 에 미치는 사리장의 영향을 조사한 결과, LPS 처리에 의 하여 매우 증가된 NO 및 PGE2의 생성이 사리장의 처리 에 의하여 강력하게 차단되었으며, 이는 NO 및 PGE2의 생성의 주요 인자인 iNOS와 COX-2의 발현 차단에 의한 것임을 알 수 있었다. 또한 사리장은 LPS에 의하여 증가 된 염증성 cytokine인 IL-1β와 TNF-α의 생성을 유의적 으로 억제하였으며, 이는 전사수준에서 이들 유전자의 발현 차단에 의한 것임을 알 수 있었다. 아울러 사리장은 항염증 효능은 염증성 반응 조절에서 가장 중요한 전사
조절인자인 NF-κB의 활성 억제와 p38MAPK 및 ERK 인 산화의 억제와 연관성이 있었다. 향후 지속적인 연구를 통하여 사리장의 항염증 성분 활성물질에 대한 동정 및 추가적인 관련 기전 연구가 요구되지만, 본 연구의 결과 는 사리장이 다양한 염증성 질환 조절제로 개발될 수 있 음을 보여 주는 것이다.
감사의 글
본 연구는 지식경제부 지역특화선도기업지원사업 및 농림수산부 식품 기능성 평가 지원 사업(Food Function- ality Evaluation program)의 지원에 의해 이루어진 것임.
참 고 문 헌
1) Sawada M, Kondo N, Suzumura A, Marunouchi T. Produc- tion of tumor necrosis factor-α by microglia and astrocytes in culture. Brain Res 491, 394-397, 1989.
2) Kriz J. Inflammation in ischemic brain injury: timing is important. Crit Rev Neurobiol 18, 145-157, 2006.
3) Hailer NP. Immunosuppression after traumatic or ischemic CNS damage: it is neuroprotective and illuminates the role of microglial cells. Prog Neurobiol 84, 211-233, 2008.
4) Monji A, Kato T, Kanba S. Cytokines and schizophrenia:
Microglia hypothesis of schizophrenia. Psychiatry Clin Neurosci 63, 257-265, 2009.
5) McGeer PL, Itagaki S, Boyes BE, McGeer EG. Reactive microglia are positive for HLA-DR in the substantia nigra of Parkinson’s and Alzheimer’s disease brains. Neurology 38, 1285-1291, 1988.
6) Dheen ST, Kaur C, Ling EA. Microglial activation and its implications in the brain diseases. Curr Med Chem 14, 1189- 1197, 2007.
7) 김일훈. 우주와 신약. 서울, 동문출판사, pp142-145, 1980.
8) 신혜영, 이은희, 김창영, 신태용, 김상덕, 송윤선, 이기
남, 김형민, 홍승헌. 자죽염의 항염활성 연구. 대한자죽
염연구회지 1, 56-64, 2003.
9) Nielsen IL, Williamson G. Review of the factors affecting bioavailability of soy isoflavones in humans. Nutr Cancer 57, 1-10, 2007.
10) Chirumbolo S. The role of quercetin, flavonols and flavones in modulating inflammatory cell function. Inflamm Allergy Drug Targets 9, 263-285, 2010.
11) Ishimi Y. Dietary equol and bone metabolism in postmeno- pausal Japanese women and osteoporotic mice. J Nutr 140, 1373S-1376S, 2010.
12) La Ferla B, Airoldi C, Zona C, Orsato A, Cardona F, Merlo S, Sironi E, D'Orazio G, Nicotra F. Natural glycoconjugates with antitumor activity. Nat Prod Rep 28, 630-648, 2011.
13) Lee MY, Seo CS, Ha H, Jung D, Lee H, Lee NH, Lee JA, Kim JH, Lee YK, Son JK, Shin HK. Protective effects of Ulmus davidiana var. japonica against OVA-induced murine asthma model via upregulation of heme oxygenase-1. J Ethnopharmacol 130, 61-69, 2010.
14) Lee Y, Park H, Ryu HS, Chun M, Kang S, Kim HS. Effects of elm bark (Ulmus davidiana var. japonica) extracts on the modulation of immunocompetence in mice. J Med Food 10, 118-125, 2007.
15) Kim EJ, Jang MK, Yoon EH, Jung CY, Nam DW, Lee SD, Kim KS. Efficacy of pharmacopuncture using root bark of Ulmus davidiana Planch in patients with knee osteoarthritis:
a double-blind randomized controlled trial. J Acupunct Meri- dian Stud 3, 16-23, 2010.
16) Choi Y, Lee MK, Lim SY, Sung SH, Kim YC. Inhibition of inducible NO synthase, cyclooxygenase-2 and interleukin-1β by torilin is mediated by mitogen-activated protein kinases in microglial BV2 cells. Br J Pharmacol 156, 933-940, 2009.
17) Resch KL, Ernst E. Garlic (Allium sativum)-a potent medicinal plant. Fortschr Med 113, 311-315, 1995.
18) Gorinstein S, Jastrzebski Z, Namiesnik J, Leontowicz H, Leontowicz M, Trakhtenberg S. The atherosclerotic heart disease and protecting properties of garlic: contemporary data.
Mol Nutr Food Res 51, 1365-1381, 2007.
19) Sabater-Molina M, Larqué E, Torrella F, Zamora S. Dietary fructooligosaccharides and potential benefits on health. J Physiol Biochem 65, 315-328, 2009.
20) Gullett NP, Ruhul Amin AR, Bayraktar S, Pezzuto JM, Shin DM, Khuri FR, Aggarwal BB, Surh YJ, Kucuk O. Cancer prevention with natural compounds. Semin Oncol 37, 258-281, 2010.
21) 김일훈. 우주와 신약. 서울, 동문출판사, pp103, 1980.
22) 최귀헌, 김창한. 유황오리 추출물의 각종 암세포에 대 한 생육억제 효과. 한국축산식품학회지 22, 348-351, 2002.
23) 윤원호, 황진용, 김창한. 유황오리로부터 항종양 활성 성분의 분리 및 정제. 한국축산식품학회지 24, 293-297, 2004.
24) Murabe Y, Sano Y. Morphological studies on neuralgia Ⅵ Postnatal development of microglia cells. Cell Tissue Res 225, 469-485, 1982.
25) Voca K, Wendt E. Divergent effects of astroglia and microglia secretions on neuronal. Exp Neurol 118, 62-72, 1992.
26) Yokozawa T, Satoh A, Nakagawa T, Yamabe N. Attenuating effects of wen-pi-tang treatment in rats with diabetic neph- ropathy. Am J Chin Med 34, 307-321, 2006.
27) Levi G, Minghetti L, Aloisi F. Regulation of prostanoid synthesis in microglial cells and effects of prostaglandin E2 on microglial functions. Biochimie 80, 899-904, 1998.
28) Knott C, Shern G, Wilkin GP. Inflammatory regulators in Parkinson’s disease: iNOS, lipocortin-1, and cyclooxygenase-1 and -2. Mol Cell Neurosci 16, 724-739, 2000.
29) Rivest S. Molecular insights on the cerebral innate immune system. Brain Behav Immun 17, 13-19, 2003.
30) Benicky J, Sánchez-Lemus E, Pavel J, Saavedra JM. Anti- inflammatory effects of angiotensin receptor blockers in the brain and the periphery. Cell Mol Neurobiol 29, 781-792, 2009.
31) Mason RP, Cockcroft JR. Targeting nitric oxide with drug therapy. J Clin Hypertens (Greenwich) 8, 40-52, 2006.
32) McGeer PL, McGeer EG. Glial reactions in Parkinson’s disease. Mov Disord 23, 474-483, 2008.
33) Gilroy DW, Lawrence T, Perretti M, Rossi AG. Inflammatory resolution: new opportunities for drug discovery. Nat Rev Drug Discov 3, 401-416, 2004.
34) Lawrence T, Willoughby DA, Gilroy DW. Antiinflammatory lipid mediators and insights into the resolution of inflam- mation. Nat Rev Immunol 2, 787-795, 2002.
35) Tassoni D, Kaur G, Weisinger RS, Sinclair AJ. The role of eicosanoids in the brain. Asia Pac J Clin Nutr 1, 220-228, 2008.
36) Boka G, Anglade P, Wallach D, Javoy-Agid F, Agid Y, Hirs- ch EC. Immunocytochemical analysis of tumor necrosis factor and its receptors in Parkinson’s disease. Neurosci Lett 172, 151- 154, 1994.
37) Davies CA, Loddick SA, Toulmond S, Stroemer RP, Hunt J, Rothwell NJ. The progression and topographic distribution of interleukin-1β expression after permanent middle cerebral artery occlusion in the rat. J Cereb Blood Flow Metab 19, 87-98, 1999.
38) Rothwell NJ, Luheshi GN. Interleukin 1 in the brain: biology, pathology and therapeutic target. Trends Neurosci 23, 618-625, 2000.
39) Hunot S, Dugas B, Faucheux B, Hartmann A, Tardieu M, Debre P, Agid Y, Dugas B, Hirasch EC. FceRII/CD23 is expressed in Parkinson’s disease and induces, in vitro, pro- duction of nitric oxide and tumor necrosis factor-α in glial cells. J Neurosci 19, 3440-3447, 1999.
40) Hanada T, Yoshimura A. Regulation of cytokine signaling and inflammation. Cytokine Growth Factor 3, 413-421, 2002.
41) Aggarwal BB, Shishodia S, Takada T, Jackson-Bernitsas D, Ahn KS, Sethi G, Ichikawa I. TNF blockade: an inflammatory issue. Ernst Schering Res Found Workshop 56, 161-186, 2006.
42) Wilms H, Zecca L, Rosenstiel P, Sievers J, Deuschl G, Lucius R. Inflammation in Parkinson's diseases and other neurode- generative diseases: cause and therapeutic implications. Curr Pharm Des 13, 1925-1928, 2007.
43) Jew SS, Bae ON, Chung JH. Anti-inflammatory effects of asiaticoside on inducible nitric oxide synthase and cycloo- xygenase-2 in RAW-264.7 cell line. J Toxicol Pub Health 19, 33-37, 2003.
44) Park SJ, Shin JS, Cho W, Cho YW, Ahn EM, Baek NI, Lee TK. Inhibition of LPS induced iNOS, COX-2 and cytokines expression by kaempferol-3-o-β-D-sophoroside through the NF-kB inactivation in RAW 264.7 cells. Korean J Pharmacogn 39, 95-103, 2008.
45) Liu SF, Malik AB. NF-κB activation as a pathological mechanism of septic shock and inflammation. Am J Physiol Lung Cell Mol Physiol 290, L622-L645, 2006.